BSDB Gurdon Summer Studentship Report (11)
Posted by BSDB, on 27 January 2017
Our final report from the 2016 class comes from Christopher Taylor, who undertook his studentship with Karim Sorefan in Sheffield.
A dual hormone response in Arabidopsis thaliana
The population problem
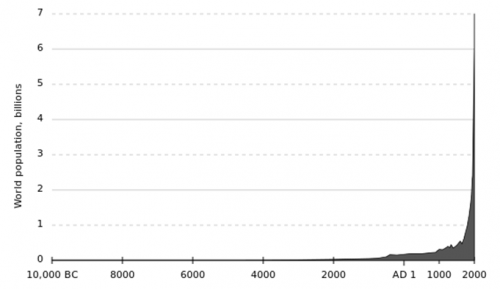
Human population growth is not a new issue. For much of mankind’s history, the species has existed in small, dispersed populations. It is only in the last 100 years where the population has truly boomed with over a fourfold increase in the number of mouths to feed (Fig 1). Demand for food is predicted to increase by 50% by 2050 and already 1 in 7 people are starving, a figure only predicted to increase.
The problems of this unsustainable growth are exacerbated by two issues:
- Climate change – in certain areas agricultural productivity is severely reduced by rising temperatures and an increase in extreme events.
- Geography – areas with greatest population growth coincide with areas most likely to be impacted by climate change. These include less economically developed countries such as those in the Sub Saharan belt.
Evidently there is an urgent need for a greener revolution.
A solution?
Plants are nature’s biochemical engineers, producing everything they need to grow, survive and reproduce from practically thin air (and a little help from the soil). Plants provide the food we eat, the food our food eats and the food that supports the very ecosystems we depend on. ‘Improving nature’s inefficiencies’ by exploiting plants’ existing biochemical pathways is one way of increasing food production in the finite space we have.
The production, transportation and signalling of plant hormones (phytohormones) allows plants to orchestrate growth and development and importantly for a changing climate, facilitate a response to environmental cues. The classic example of auxin’s role in phototropism may spring to mind. Here auxin exhibits a negatively phototropic response, moving away from the prevailing light direction. This leads to an accumulation of auxin molecules in the side furthest from the light causing acidification of the cell walls and allowing for cell elongation and plant growth. Ultimately this produces a plant (or stem) whose curvature is maximised for intercepting incoming light.
Exploiting these pathways already has agricultural applications. Think of taking a plant cutting, dabbing the base in auxin and growing a whole new plant.
My work in the Sorefan lab in the University of Sheffield’s department of molecular biology and biotechnology (MBB), was to investigate the role of a novel hormonal response produced by auxin and cytokinin. Previous microarray data by the lab revealed an interesting occurrence when Arabidopsis thaliana seedlings were grown under conditions of elevated auxin (IAA) and cytokinin (BAP) concurrently. When grown under a dual hormone combination, the seedlings showed a significant change in the number and types of genes that were either significantly up or downregulated in the microarray. This response was larger than either responses of auxin or cytokinin alone and has been termed the ‘dual hormone response’ (Fig 2).
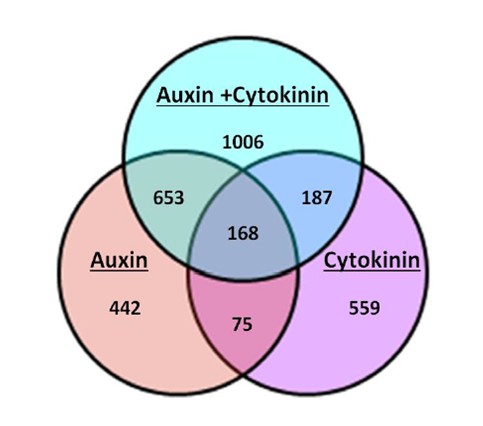
Functions of the dual hormone response
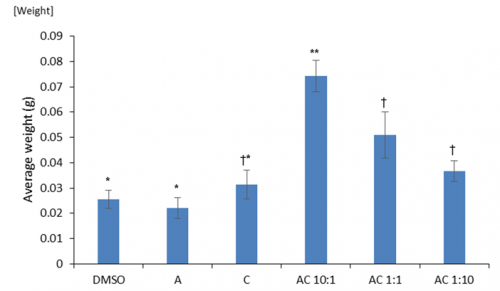
Prior to environmental stresses we tested if there were any general positive growth responses of the dual hormone response (DHR). We investigated the effects of differing ratios of auxin to cytokinin on cotyledon weight. Cotyledons are embryonic leaves that are among the first structures to emerge following germination and their weight is an accurate proxy of seedling health. We found significantly greater cotyledon mass under DHR conditions particularly with ratios showing high concentrations of auxin (Fig 3). Interestingly, this was significantly higher than the effects of just auxin alone, implying that the exogenous application of even a small amount of cytokinin may produce positive benefits.
Comparing our lab’s dual hormone microarray data with that of other labs highlighted approximately 11.2% of our identified genes may show some response to heat stress conditions. I produced cDNA from seedlings grown under different combinations of hormone treatments and designed primers for the heat stress responsive genes identified by these microarrays. Using multiple quantitative real-time polymerase chain reactions (qRT-PCR), I confirmed that a number of these heat-responsive genes were also significantly affected by the dual hormone response.
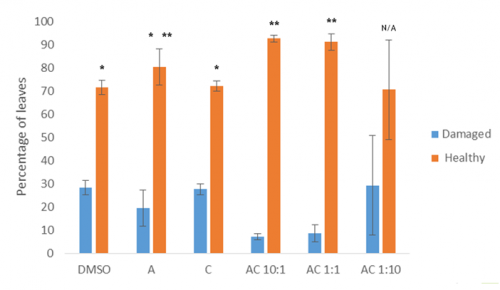
I then conducted an experiment to investigate the phenotypic effects of heat shock on seedlings grown under DHR conditions (Fig 4).

Preliminary data from this experiment suggests that the dual hormone response may convey some resistance to heat stress, as combinations of the two hormones results in significantly fewer damaged / unhealthy leaves than the control or either single hormone alone (Fig 5).
Future work aims to quantify chlorophyll more effectively as our own analyses were confounded by the production of purple anthocyanin pigments, which are often produced as a response to stress.
These data may therefore allow us to apply dual hormone responses to agriculture. The dual hormone treated plants were larger and appear to be somewhat de-sensitized to heat stress, which may allow us to manipulate crop plants to convey some heat resistance to them. Knowing the optimal hormone ratios is important for applying this information. Manipulating plants in this way is a promising route for producing ‘climate ready’ crop species and ensuring that we can continue to feed a world of 7 or 10 billion in a vastly deteriorating climate.
Christopher Taylor
 (1 votes)
(1 votes)