How to Grow a Network on the Go
Posted by schenz, on 8 June 2018
Transportation networks play a central role in enabling efficient mass flow over extended domains, where diffusion alone would be too slow. Therefore, transportation networks often play a central role for an organism’s physiology and a high degree of energetic efficiency has been proposed as a guiding principle for the layouts of these networks.
However, biological organisms usually do not construct their networks only after fully developing the body plan, but instead extend them together with the growth of the body, for example in trees, fungi, or myxomycetes. Then it is interesting to ask how they achieve a high degree of efficiency, given that at the time of network construction the final shape of the organism is yet unknown.
Physarum polycephalum: a Crawling Network

The true slime mould Physarum polycephalum (see fig. 1) is a good model for this question, for two reasons. First, it is an easy-to-handle, macroscopic organism that prominently features an adaptive tube network as it extends over surfaces. More importantly, though, we have an understanding of the rules that shape this network under static conditions: a simple set of equations formulated by Tero et al. (J. Theor. Biol. 2007) known as the ‘current reinforcement rule’.
To learn how network formation occurs during extension, it makes sense to first study the organism’s behaviour under this condition and then find a model that explains this behaviour. A model enables us to make testable predictions that can decide whether we have, in fact, understood the basic characteristics of the organism and to include certain assumptions about the mechanism behind them.
Study Approach: Learn, Model, Check
1. Learn
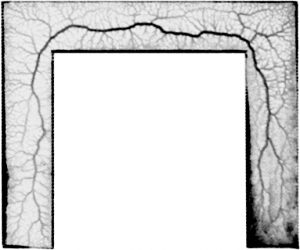
In a study published last year (Schenz et al., J. Phys. D 2017), we first studied how the network of Physarum under extension is shaped. We let the organism extend through a narrow lane that includes some turns (see fig. 2). The organism constructed its main veins at a small distance behind the growth front and did so at a time long before it had fully explored the complete layout of the arena. The main characteristic of the resultant vein trajectory is that it cuts corners at turns but then returns back to the centre line of the arena, even between two turns, where the globally shortest path would dictate to remain on the inside edge.
Nevertheless, analysis showed that the slime mould’s main vein was only 6% longer than the shortest possible route through the arena. To appreciate this, one has to consider that a naïve strategy of constructing the vein in the centre of the corridor would result in a trajectory 18% longer than the minimum. How can we explain such a high efficiency in the absence of foresight?
2. Model
As a model we considered first the classical current reinforcement model, but it failed to reproduce the characteristic vein pattern. Therefore, current reinforcement dynamics alone are insufficient to explain the organism’s capability. As a consequence we constructed a novel model consisting of three core elements that have each been successfully used in the literature to describe specific aspects of Physarum physiology: wave front dynamics, Calcium-driven oscillation waves, and the current reinforcement tube model. We linked these three elements with appropriate interactions under the assumption that the observed expansion behaviour and network structure are a consequence of their interplay.
The resulting model explained the phenotypical features well both qualitatively and quantitatively, and also contained some physiological assumptions that are testable. We could thus determine that the coupling between growth front extension and tube network evolution has to be of just the right strength to allow the organism some spatial integration necessary to find local optimality of the route trajectory as well as efficient transportation of body mass through the tube network from the rear to the front.
3. Check
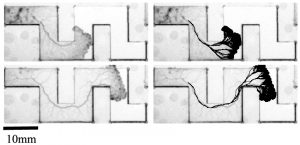
The corollary of this finding is that growth front extension history alone should be sufficient to make a prediction on the main vein trajectory (see fig. 3). This hypothesis yielded much better predictions than alternative explanations we tested. This then yields a mechanistic explanation of how the organism can achieve this high degree of optimality in the face of uncertainty and at the same time an interpretation for the biological context for the current reinforcement rule: to enable efficient locomotion of Physarum.
In our ongoing research we develop further the question of what measure of optimality is likely guiding an expanding network. Total network length, as considered above, is not the only dimension along which a network can be evaluated, especially if it is embedded in a foraging organism. We will, therefore, search for such a measure that best predicts the behaviour of the organism given greater degrees of freedom to then evaluate well established concepts such as Optimal Foraging Theory in the context of continuous, network-based systems. This will allow us to consider how efficient networks have to be structured in an unknown environment.
 (1 votes)
(1 votes)