The First International Hemichordate Meeting: the Birth of a Community
Posted by Tetsuto, on 24 January 2017
Tetsuto Miyashita
Certain scientific meetings have their place in the book of history as a visible shift — or a turning point, if you will — in a particular field. The Cuvier-Geoffroy debate at the French Academy of Sciences in 1830 set the tone for the subsequent two centuries of biology in search for determinants of organismal forms. Closer to our times, you might think of the Santa Fe workshop that kick-started the Human Genome Project or, even more recently, the creation of a new society (the Pan-American Society of Evolutionary Developmental Biology) following a NESCent workshop. Common threads connecting many such examples are small sizes that allow intellectual catalysis (typically no more than 50 people participating) and timing them just ahead of the curve. The First International Hemichordate Meeting held at Hopkins Marine Station (Stanford University) from December 9 to 11 2016 may be remembered as just such a shift in the history of study in chordate origins.

A Prelude to the First Hemichordate Meeting
Hemichordates are a small phylum of deuterostomes. The most conspicuous members — enteropneusts — are commonly called acorn worms, and their preposterous appearances are unforgettable. Hemichordates contain another living lineage of deep-sea, tube-dwelling, filter-feeding forms called pterobranchs. As the name suggests, hemichordates have long been considered our close invertebrate relatives, nested just outside chordates. Unlike their sister lineage that is echinoderms, hemichordates sport pharyngeal slits, and since William Bateson, embryologists have been looking for a homologue of notochord or even a bona fide notochord itself in acorn worms. So, classically, this is the group of animals that supposedly root many characters of chordates. To the post-Bateson chordate researchers, however, hemichordates seemed too distant and too different to inform vertebrate origins directly. Cephalochordates and tunicates each have clearly vertebrate-like morphological traits, even though a closer look at their development does not support simple progression to vertebrates from either body plan.
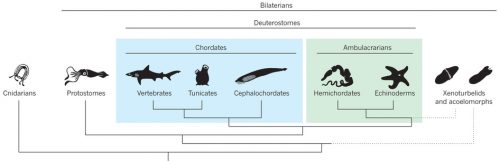
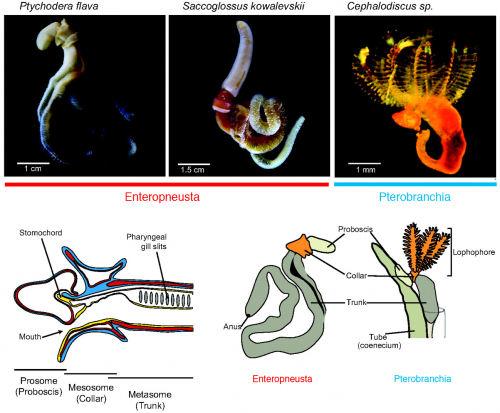
But the tide is turning, and hemichordates are making a remarkable comeback onto the scene of the great chordate debate. I will touch on how and why in this report, and the change is palpable. In 2013, I attended a symposium “The Making of a Vertebrate” organized by RIKEN CDB in Japan, where even the conveners expressed their astonishment at how they ended up with so many (five!) hemichordate talks and how insights from hemichordates led the discussion throughout the symposium. I remembered this vividly. It did feel like that hemichordates are catching on.


Three years since the RIKEN symposium, Chris Lowe (Stanford University) and Yi-Hsien Su (Academica Sinica) organized the First International Hemichordate Meeting. Hemichordate workers are sending a signal that the community is coming together, and that there is a plenty of exciting stuff to come. In my view, there are three different streaks among modern hemichordate workers: the Saccoglossus school that mainly uses the direct-developing acorn worm S. kowalevskii, geographically centered in North America; the Ptychodera school that mainly uses the indirect-developing P. flava, centered in Japan, Taiwan, and Hawaii; the pterobranch school that traces its origin to the study of ‘lophophorates’ (and perhaps most naturalist in its spirit — so this school also includes those who look at the strange family of deep-sea acorn worms, torquaratoriids). My report is organized along with this classification.
Hemichordates and the ‘Holy Grail’ of the Chordate Debate
Chris Lowe opened the discussion by providing an overview of the research that fueled this hemichordate revival. This includes the seminal papers describing expression profiles in the ectoderm of S. kowalevskii here and here. He outlined the prospect and challenges to identify shared features of hemichordate and vertebrate development. This introduction reflects a cautious approach to what some may consider as support for homology, especially around the holy grail of notochord. Nori Satoh (OIST) certainly resonates with it. About the hotly debated stomochord — an endodermally derived supporting structure of the proboscis, proposed as a notochord homologue by Bateson — he pointed out FoxE expressions shared not with notochord but rather with the endostyle. Coupled with the fact that hemichordate D-V axis is opposite to that of vertebrates, Satoh asked at what developmental stage hemichordates should be compared with chordates. The prospect is bright to recognize developmental traits of deuterostomes, but the window of comparison may be narrow for comparison of chordate traits. He highlighted a locomotory shift from ciliary bands to tail muscles as a significant event toward chordates and outlined his current efforts to characterize these evolutionary transitions through comparative genomics. He explores these thoughts further in the new book from the Academic Press, Chordate Origins and Evolution.

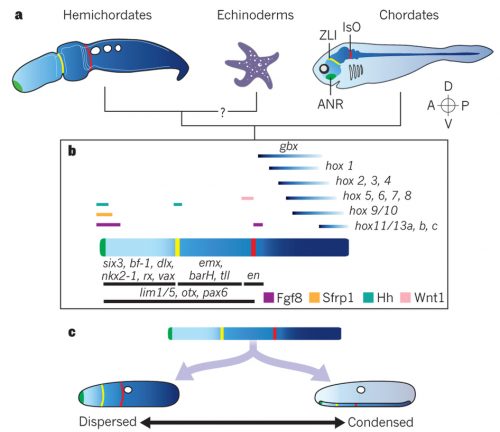
Genomics Facilitate Refined Comparative Approaches
When it comes to genomics, Dan Rokhsar (University of California, Berkley) has acted as a hub of the Hemichordate Genome Project that published genomes on the direct-developing S. kowalevskii and the indirect-developing P. flava. Some of his comments that attracted interest from the audience are about conservation of synteny and about ‘pharyngeal’ gene clusters. The published genomes to which many other participants of the meeting contributed now form a foundation for some of the exciting ‘omics’ research going on. Jessica Gray (Harvard University) is certainly among the movers and shakers on this frontline. Her work on miRNA regulatory networks in S. kowalevskii, particularly those of neural and muscle development, captured attention from the audience (which also won a presentation award earlier in the inaugural meeting of the Pan-Am Evo-Devo Society). She and Marc Kirschner (Harvard University) are pioneering single-cell transcriptomics in S. kowalevskii. Kirschner’s talk and Gray’s poster caused quite an excitement — many participants can think of unique and interesting applications that could advance their research, especially by correlating the transcriptomic landscape with the observed expression profiles of marker genes.
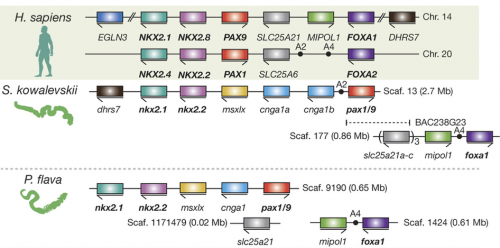
These technical advances are indispensable to the ongoing efforts to compare S. kowalevskii with vertebrates. Stephen Green (California Institute of Technology) brought mesoderm to the discussion. Green is unique among the participants for using lamprey as a comparative model. He suggests that differential patterning of anterior and posterior mesoderm in S. kowalevskii perhaps shares an evolutionary root in the deuterostome stem with the cranial versus somitic differentiation in vertebrates. Rather than looking at region of a germ layer, Elena Silva (Georgetown University) focused on deriving neural progenitors. Here, too, parallels are observed with the pathways specifying neural progenitors in vertebrates. In the poster session, the Lowe lab displayed the current state of the art in research on S. kowalevskii that employs reporter expression, functional tools (CRISPR, RNAi), behavioral assays. These are Paul Minor, Nat Clarke, and José Lopez. But it was John Gerhart (University of California, Berkeley) who provided weight to this discussion. In the closing lecture of the meeting, he explored the question whether or not S. kowalevskii has a vertebrate organizer. This isn’t a run-of-the-mill comparison. Gerhart and his colleagues have been cloning almost every thinkable gene from S. kowalevskii that would be important in evolutionary context. Based on this massive dataset — a major portion of which remains to be published — he critically tested predictions arising from a hypothesis that the organizer evolved in the hemichordate-vertebrate common ancestor. This analysis intersects with such standing questions in hemichordate research as: Do hemichordates have a notochord homologue? How is the A-P patterning set in hemichrodates? How did the D-V axis evolve in deuterostomes?
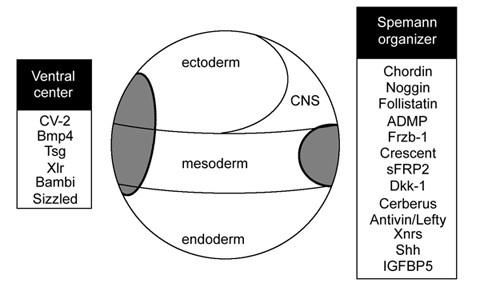
Regarding the polarity, the A-P patterning had multiple talks. Sébastien Darras (L’Observatoire Océanologique de Banyuls-sur-Mer) took advantage of functional tools available in S. kowalevskii to characterize the role of Wnt in establishing the A-P axis. Jens Fritzenwanker (Georgetown University) addressed axial elongation with a focus on the posterior growth zone. Yann Le Petillon (Academia Sinica) used the indirect-developing enteropneust, P. flava, to illustrate an interaction of Wnt and BMP signaling pathways to pattern and restrict anterior neural ectoderm. Ryan Range (Mississippi State University) gave an engaging talk by mapping complex GRNs for the anterior sensory organ of sea urchin larvae and dissecting multiple and complementing functions of Wnt pathways. This talk features his latest and very neat paper in Development.
Perspectives from Indirect-developing Enteropneusts
The indirect developing P. flava constitutes another stream of research on hemichordate development with the Asia-Pacific distribution. Kuni Tagawa (University of Hiroshima) provided this historical context by looking back on Tom Humphreys’s career. Tagawa was called on to Hawaii by Nori Satoh to work with Humphrey and collect P. flava. Early results from this collaboration include the paper describing Brachyury expression in the context of testing for a notochord homologue. Hiroshi Wada (University of Tsukuba) is another student of Satoh’s. He discussed a pygochord in Balanoglossus simodensis (a Japanese enteropneust). Whether this structure can be considered potential homologue of notochord will remain controversial, to say the least.

{kind=link}
Over the last several years, Yi-Hsien Su and Jr-Kai “Sky” Yu (Academica Sinica) have built a team now leading the P. flava research (as well as echinoderms and cephalochordates). Su has a stable supply of embryos from a Taiwanese locality, and some of the research that is taking off from the lab was featured in the meeting. Her own work concerns the regulation of BMP signaling to establish D-V axis among ambulacularians (the lineage containing hemichordates and echinoderms). In addition to Le Petillon’s work on the A-P patterning, Tzu-Pei Fan from the Su lab reported on FGF signaling in the mesoderm development of P. flava.
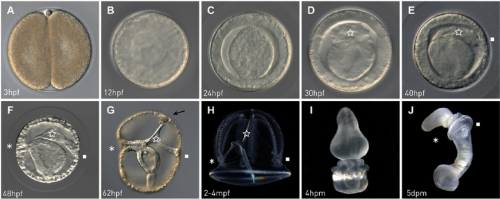
Billie Swalla (Friday Harbor Laboratories) is another long-time P. flava expert and a chief architect of the current chordate phylogenetic framework. Her team contributed some unique perspectives to the meeting by looking at regeneration by regions — proboscis, collar, and trunk. Her student Shawn Luttrell explored this work further in the context of stem cells. Formerly at the Swalla lab and now at the Halanych lab, Michael Tassia (Auburn University) called our attention to the immune system with phylogenetics and bioinformatics of Toll-like receptor pathways in bilaterians and showed that hemichrodates are the key taxon to consider the evolution of immunological memory toward vertebrates.
Paul Gonzalez (Stanford University) delivered a talk literally hot off the press — published in Current Biology just days ahead of the meeting. He pioneered another indirect-developing enteropneust, Schizocardium sp. in the Lowe lab. Surprisingly, the larvae lack expression profiles characteristic of the Hox-patterned trunk — in Richard Strathmann’s words, they are ‘swimming heads’. The trunk expression profiles appear across the metamorphosis. His work hints great potentials in comparison between direct- and indirect-developing enteropneusts in the context of deuterostome evolution.

{kind=link}
Pterobranchs Are No Red Herring
Speaking of unconventional, you must have realized by now that I saved the most curious topic in hemichordate research to the last. Although a major class of hemichordates called enteropneusts receives much of the attention — and although this hemichordate conference so far could have been an enteropneust conference — a minor class called pterobranchs has always generated interesting questions. The living pterobranchs are represented by only two, mostly deep-sea, tube-dwelling genera: Cephalodiscus and Rhabdopleura. They used to be a thriving group, says Chris Cameron (Université de Montréal). He and his collaborators in paleontology are reaching consensus that graptolites, common fossils of the Paleozoic rocks, are nested within the Pterobranchia. So the two genera are quite literally living fossils. Excitingly, now he has a Mediterranean locality that provides a stable supply of the living pterobranchs, although the initial work is still ongoing. Cameron’s interest is to identify root conditions for hemichordates and ambulacularians. In that vein, he also reviewed stem enteropneusts from the Cambrian and Carboniferous localities (Cambrian stem enteropneusts were tube-dwelling! But, as Ken Halanych points out, so are the living Antarctic torquaratoriids) and described echinoderm-like mineralized ossicles in the living enteropneusts.

{kind=link}
It probably becomes apparent by this point that no good work in biology is possible without systematics, whether talking about enteropneusts or pterobranchs. Gonzalez’s species of Schizocardium does not even have a name yet. The evolutionary narratives based on developmental traits — a major justification for why looking into hemichordates — entirely depends on the current phylogenetic scheme. Even the major ambulacularian-chordate schemes accepted by most workers today can be shaken up easily. With xenoturbellids falling out from the deuterostome stem, for example. Ken Halanych (Auburn University) has been instrumental to reorganize and clean up this area over the last two decades, and his eloquent talk on hemichordate systematics reflected his naturalist spirit well. The audience was captivated by the giant torquaratoriid from Antarctica, for example. His pep talk for pterobranchs certainly upped the enthusiasm for this minor class by a notch for Cameron’s talk. Joie Cannon (University of California, Santa Barbara) extended Halanych’s theme by going through her PhD work that took her around the world several circles to collect specimens for molecular phylogeny (see her papers here, here, and here). In hemichordates, so many taxa remained undescribed or even yet to be discovered. There is not a single tome or even a global guide that the workers can refer to for species identification. Cannon’s work laid the track, but no train has rolled on it yet. Like many workers who work with rather uncharismatic invertebrates, there is an acute need for systematics. The field is waiting.

{kind=link}
In conclusion of the meeting, Lowe led a roundtable discussion. Some points that stood out are: the Marine Biological Laboratory, Woods Hole, has been a center of research on S. kowalevskii. Lowe is consulting with the Lab for increasing accessibilities and improved facities that could accommodate more researchers. Su followed up on this by extending an open invitation to collaborate on P. flava in Taiwan as well. Cameron and Halanych proposed creating some kind of portal — a listserver would be a start. Green described the current challenges to maintaining a lab culture of S. kowalevskii in his home lab at Caltech.
Prospectus of Hemichordate Research
As a graduate student, I was thrilled to witness a community forming. The modern hemichordate research has clearly hit the critical mass that now some community efforts are required to carry it to the next stage. The Hemichordate Genome Project got the train moving, and more and more people are jumping on it to take a ride. The ever-increasing developmental information, coupled with evolutionary insights based on sound phylogenetic framework, should transform our understanding of this small phylum over the next decade or two. The meeting showcased enough of the momentum.

Simultaneously, some challenges still remain. In my view, the poor understanding of hemichordate natural history (taxonomy, anatomy, behavior, etc.) is holding back further progress. This gap in our knowledge is even more glaring today than ever, not least because favourite habitats of enteropneusts used for lab research appear to be shrinking rapidly. It echoes the outcry from biological communities in general: “We need more taxonomists!” On other counts, pterobranch development remains as a bottleneck. The debate on the deuterostome stem has not settled. None of the currently used hemichordate models has undergone a full lifecycle in lab.
Historically, the hemichordate research has relied on no more than several active labs working concurrently. So the meeting had the feel of a party in the neighbourhood: polite and friendly. No heated debate ensued even when philosophical differences were apparent. This civil attitude is welcome, but for the community to be vibrant, a controversy, too, may help. Wild speculations should also have a place in the field that is still in its infancy. One curious observation from the meeting is that all but one of those who compared hemichordates and vertebrates explicitly referred to Xenopus. Surely, Xenopus is a useful model, and has been the one of choice at the Gerhart and Kirschner labs. But would different predictions and interpretations be possible if compared with other vertebrate models? How should one identify deep homology and cooption event in that comparative framework? To create this kind of discourse, perhaps there were few opportunities for all of the participants to get together except for reception on the eve and banquet in the finale. All in all, though, the prospect is wide open, and the next meeting is being planned a couple of years from now. The second meeting will mark another memorable point in the history of hemichordate research. Until then, my new worm-loving friends!
 (5 votes)
(5 votes)Get involved
Create an account or log in to post your story on the Node.
Sign up for emails
Subscribe to our mailing lists.