In Development this week (Vol. 141, Issue 2)
Posted by Seema Grewal, on 31 December 2013
Here are the highlights from the current issue of Development:
Peri important role for Notch
Pericytes are specialised cells that wrap around the endothelial cells of the vasculature to regulate vascular integrity, permeability and blood flow. Despite this crucial role, the molecular mechanisms that control pericyte development are not well understood. In this issue, two papers identify a requirement for Notch in pericyte development in the brain and kidney vasculature.
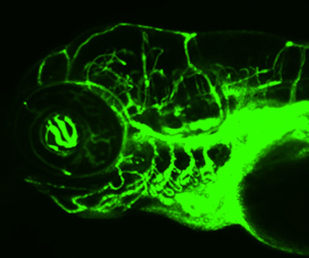 On p. 307, Bruce Appel and colleagues investigate the role of Notch in regulating pericyte number in the developing zebrafish brain vasculature. The authors interrogate a panel of Notch genes and identify notch3 as expressed in the developing vasculature, specifically in cells positive for pdgfrb, a known pericyte marker. Loss-of-function of Notch3 leads to disruption of the blood-brain barrier and cerebral haemorrhaging, which is likely to be due to the reduction in pericyte number. Importantly, the authors show that Notch3 is required for pericyte development and specifically for promoting proliferation and expansion of the cells. Using pdgfrb expression as a readout, the authors observe that overexpression of the Notch3 intracellular domain is associated with increased numbers of pericytes, whereas interference with Notch3 activity causes a reduction. Based on varying levels of pdgfrb expression observed throughout the study, the authors hypothesize that Notch3 may positively regulate pdgfrb in order to regulate pericyte proliferation.
On p. 307, Bruce Appel and colleagues investigate the role of Notch in regulating pericyte number in the developing zebrafish brain vasculature. The authors interrogate a panel of Notch genes and identify notch3 as expressed in the developing vasculature, specifically in cells positive for pdgfrb, a known pericyte marker. Loss-of-function of Notch3 leads to disruption of the blood-brain barrier and cerebral haemorrhaging, which is likely to be due to the reduction in pericyte number. Importantly, the authors show that Notch3 is required for pericyte development and specifically for promoting proliferation and expansion of the cells. Using pdgfrb expression as a readout, the authors observe that overexpression of the Notch3 intracellular domain is associated with increased numbers of pericytes, whereas interference with Notch3 activity causes a reduction. Based on varying levels of pdgfrb expression observed throughout the study, the authors hypothesize that Notch3 may positively regulate pdgfrb in order to regulate pericyte proliferation.
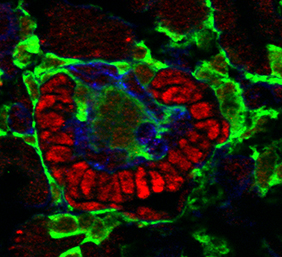 The role of Notch signalling in pericyte development is also investigated by Raphael Kopan and colleagues (p. 346), who report a critical requirement for Notch during the development of the pericytes of the mammalian kidney, known as mesangial cells. These cells, along with the smooth muscle and interstitial cells of the kidney, derive from Foxd1+ stromal precursors; however, Notch signalling appears to be only required for the emergence of the mesangial cells. Inactivation of Notch specifically in the stromal precursors results in the formation of glomeruli that lack mesangial cells, leading to glomerular aneurism and kidney failure at birth. The authors go on to show that, in this case of pericyte development in the kidney, Notch1 and Notch2 appear to act redundantly.
The role of Notch signalling in pericyte development is also investigated by Raphael Kopan and colleagues (p. 346), who report a critical requirement for Notch during the development of the pericytes of the mammalian kidney, known as mesangial cells. These cells, along with the smooth muscle and interstitial cells of the kidney, derive from Foxd1+ stromal precursors; however, Notch signalling appears to be only required for the emergence of the mesangial cells. Inactivation of Notch specifically in the stromal precursors results in the formation of glomeruli that lack mesangial cells, leading to glomerular aneurism and kidney failure at birth. The authors go on to show that, in this case of pericyte development in the kidney, Notch1 and Notch2 appear to act redundantly.
Roadmap for neuronal specification
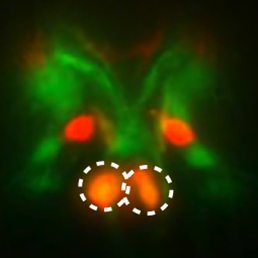 Neuronal subtype specification is regulated by the coordinated action of transcription factors. Any one factor may be expressed in multiple subtypes, but specification is achieved based on the precise combination of factors and is therefore context dependent. In this issue (p. 422), Oliver Hobert and colleagues explore neuronal differentiation in C. elegans and focus on the role of the TTX-3 LIM homeodomain transcription factor in regulating neural subtype specification. The authors find that TTX-3 is broadly required in multiple neuron classes of relatively unrelated identity, but that the interacting partners and downstream targets of TTX-3 are subtype specific. TTX-3 is required for cholinergic AIY interneuron specification, while an interaction with the POU domain protein UNC-86 leads to the specification of serotinergic NSM neurons. Furthermore, UNC-86 itself can specify cholinergic IL2 sensory and URA motoneurons via cooperation with the ARID-type transcription factor CFI-1. This detailed analysis of transcriptional cascades reveals a programming roadmap for neuronal subtype specification.
Neuronal subtype specification is regulated by the coordinated action of transcription factors. Any one factor may be expressed in multiple subtypes, but specification is achieved based on the precise combination of factors and is therefore context dependent. In this issue (p. 422), Oliver Hobert and colleagues explore neuronal differentiation in C. elegans and focus on the role of the TTX-3 LIM homeodomain transcription factor in regulating neural subtype specification. The authors find that TTX-3 is broadly required in multiple neuron classes of relatively unrelated identity, but that the interacting partners and downstream targets of TTX-3 are subtype specific. TTX-3 is required for cholinergic AIY interneuron specification, while an interaction with the POU domain protein UNC-86 leads to the specification of serotinergic NSM neurons. Furthermore, UNC-86 itself can specify cholinergic IL2 sensory and URA motoneurons via cooperation with the ARID-type transcription factor CFI-1. This detailed analysis of transcriptional cascades reveals a programming roadmap for neuronal subtype specification.
How the zebra(fish) got its stripes
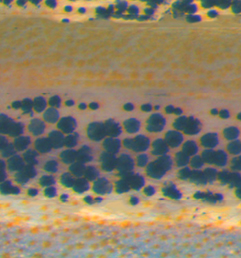 The striped pattern of the zebrafish skin offers an excellent model system in which to study biological pattern formation. Previous studies have shown that the interactions between melanophores and xanthophores are crucial for pattern formation, but little is known regarding the molecular mechanisms that regulate this phenomenon. Now, on p. 318, Shigeru Kondo, Masakatsu Watanabe and colleagues uncover a role for long-range Delta/Notch signalling between the melanophore and xanthophore pigmented cell types that is crucial for proper stripe formation. The authors show that Delta/Notch signalling is required for melanophore survival, since disruption of the pathway by DAPT treatment results in loss of melanophores, while constitutive Notch activation in transgenic fish rescues this effect. The authors use targeted laser ablation to show that the source of this survival signal is the xanthophore. Interestingly, the authors observe long protrusions that originate from the melanophores and extend to the xanthophores, which might serve as a means to mediate the Delta/Notch signalling over long distances.
The striped pattern of the zebrafish skin offers an excellent model system in which to study biological pattern formation. Previous studies have shown that the interactions between melanophores and xanthophores are crucial for pattern formation, but little is known regarding the molecular mechanisms that regulate this phenomenon. Now, on p. 318, Shigeru Kondo, Masakatsu Watanabe and colleagues uncover a role for long-range Delta/Notch signalling between the melanophore and xanthophore pigmented cell types that is crucial for proper stripe formation. The authors show that Delta/Notch signalling is required for melanophore survival, since disruption of the pathway by DAPT treatment results in loss of melanophores, while constitutive Notch activation in transgenic fish rescues this effect. The authors use targeted laser ablation to show that the source of this survival signal is the xanthophore. Interestingly, the authors observe long protrusions that originate from the melanophores and extend to the xanthophores, which might serve as a means to mediate the Delta/Notch signalling over long distances.
Neural progenitors divide and conquer
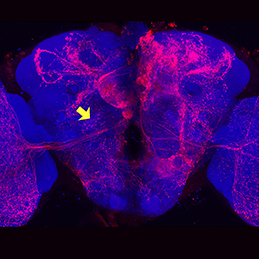 Neuronal diversity in Drosophila is generated by the temporal specification of type II neuroblasts (NBs) and their progeny, the intermediate neural progenitors (INPs). Multiple transcription factors are expressed in a birth order-dependent manner within each INP lineage, but whether this temporal patterning gives rise to discrete neuronal sets from each individual INP cell is unclear. Now, on p. 253, Tzumin Lee and colleagues describe extensive fate-mapping of individual neurons derived from specific type II NB lineages. The authors use targeted clonal labelling to specifically label neurons in individual INP clones, and by restricting the clonal induction to specific time windows they are able to generate and characterise clones of neurons that are born from two successively produced INPs. The resulting analyses demonstrate that the temporal specification of INPs does indeed translate to distinct types of neurons, suggesting that neuronal fate diversification might operate as a function of age.
Neuronal diversity in Drosophila is generated by the temporal specification of type II neuroblasts (NBs) and their progeny, the intermediate neural progenitors (INPs). Multiple transcription factors are expressed in a birth order-dependent manner within each INP lineage, but whether this temporal patterning gives rise to discrete neuronal sets from each individual INP cell is unclear. Now, on p. 253, Tzumin Lee and colleagues describe extensive fate-mapping of individual neurons derived from specific type II NB lineages. The authors use targeted clonal labelling to specifically label neurons in individual INP clones, and by restricting the clonal induction to specific time windows they are able to generate and characterise clones of neurons that are born from two successively produced INPs. The resulting analyses demonstrate that the temporal specification of INPs does indeed translate to distinct types of neurons, suggesting that neuronal fate diversification might operate as a function of age.
Case closed: ion channels mediate dorsal closure
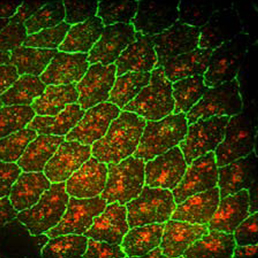 Dorsal closure is a morphogenic process that involves the interplay of mechanical forces as two opposing epithelial sheets come together and fuse. These forces impact cell shape and the rate of morphogenesis, but the molecular pathways that translate mechanical force into phenotype are not well understood. Now, on p. 325, Daniel Kiehart and colleagues demonstrate a role for calcium signalling via mechanically gated ion channels (MGCs) in Drosophila dorsal closure. Using UV-induced calcium release, the authors show that increased calcium levels stimulate contractility during dorsal closure, whereas treatment with a calcium-chelating agent disrupts closure. Via a series of pharmacological perturbations, the authors demonstrate that MGCs regulate actomyosin contraction that, in turn, is required for force production and successful dorsal closure. The authors support their findings by knocking down two separate MGC subunits, which also leads to a failure to generate sufficient force for dorsal closure. This study paves the way for investigating MGCs in other morphogenic processes, for example during wound repair.
Dorsal closure is a morphogenic process that involves the interplay of mechanical forces as two opposing epithelial sheets come together and fuse. These forces impact cell shape and the rate of morphogenesis, but the molecular pathways that translate mechanical force into phenotype are not well understood. Now, on p. 325, Daniel Kiehart and colleagues demonstrate a role for calcium signalling via mechanically gated ion channels (MGCs) in Drosophila dorsal closure. Using UV-induced calcium release, the authors show that increased calcium levels stimulate contractility during dorsal closure, whereas treatment with a calcium-chelating agent disrupts closure. Via a series of pharmacological perturbations, the authors demonstrate that MGCs regulate actomyosin contraction that, in turn, is required for force production and successful dorsal closure. The authors support their findings by knocking down two separate MGC subunits, which also leads to a failure to generate sufficient force for dorsal closure. This study paves the way for investigating MGCs in other morphogenic processes, for example during wound repair.
PLUS…
How to make a primordial germ cell
 Primordial germ cells (PGCs) are the precursors of sperm and eggs, which generate a new organism that is capable of creating endless new generations through germ cells. Here, Magnúsdóttir and Surani summarise the fundamental principles of PGC specification during early development and discuss how it is now possible to make mouse PGCs from pluripotent embryonic stem cells, and indeed somatic cells if they are first rendered pluripotent in culture. See the Primer on p. 245
Primordial germ cells (PGCs) are the precursors of sperm and eggs, which generate a new organism that is capable of creating endless new generations through germ cells. Here, Magnúsdóttir and Surani summarise the fundamental principles of PGC specification during early development and discuss how it is now possible to make mouse PGCs from pluripotent embryonic stem cells, and indeed somatic cells if they are first rendered pluripotent in culture. See the Primer on p. 245
Retinal neurogenesis
 In their Development at a Glance article, Centanin and Wittbrodt provide an overview of retinal neurogenesis in vertebrates and discuss implications of the developmental mechanisms involved for regenerative therapy approaches. See the poster article on p. 241
In their Development at a Glance article, Centanin and Wittbrodt provide an overview of retinal neurogenesis in vertebrates and discuss implications of the developmental mechanisms involved for regenerative therapy approaches. See the poster article on p. 241
 (No Ratings Yet)
(No Ratings Yet)Get involved
Create an account or log in to post your story on the Node.
Sign up for emails
Subscribe to our mailing lists.