A PhD studentship is available in Dr. Claudia Barros’ group, Peninsula School of Medicine, Plymouth University, UK. The project is devoted to characterise the role of novel candidate genes in brain tumour initiation identified in the laboratory. It will focus on investigating pathways acting in mitochondria, the cells’ power plants, but it is not restricted to it. Drosophila will be used to examine candidate genes in the conversion of normal neural stem lineage cells into brain tumour initiation cells via gain and loss of function assays, providing a unique opportunity to unravel their role in live brains at single-cell resolution. Translation of findings will be performed using glioma tumour samples and patient-derived glioblastoma stem cell lines.
We are looking for a candidate with experience in genetics, molecular biology, immunohistochemistry, cell culture, biochemistry and/or confocal imaging. A high interest in neural stem cell and brain tumour biology is a must. Applications are invited from qualified graduates (1st class or 2:1, equivalent), and preferably a relevant Masters or extended laboratory research experience. International students must have an IELTS score of min 7.0 (with no less than 6.5 in any element).
For further information regarding the project, please contact Dr. Claudia Barros (claudia.barros@plymouth.ac.uk). Applications must be made in accordance with the details shown below.
Funding Notes:
The studentship is supported for 3 years and includes full Home/EU tuition fees plus a stipend of £14,553 per annum. The studentship will only fully fund those applicants who are eligible for Home/EU fees with relevant qualifications. Applicants required to cover overseas fees will have to cover the difference between the Home/EU and the overseas tuition fee rates. For the 17/18 academic year the difference in fee is £10350. If you are selected you will be required to provide financial assurances.
Shortlisted candidates will be invited for interview. We regret that we may not be able to respond to all applications. Applicants who have not received an offer of a place by end of May 2018 should consider their application has been unsuccessful on this occasion.
Skin is a fascinating organ that dynamically adjusts its size throughout our lifetimes and does so perfectly while, at the same time, acting as a robust bi-directional barrier protecting us from the insults of the outside world. Of particular interest is the epidermis, which is the outermost, barrier- and load-bearing layer of the skin. Every time we bend our fingers, undergo cycles of weight loss or gain, or are a subject to life-altering events, such as pregnancy, the epidermis has to shrink, expand, and withstand various amounts of tension while still maintaining integrity! From a point of view of an engineer, skin is a marvel – how can this be? What is the structure of this stunning tissue allowing it to perform the many functions that it has?
The epidermis is a highly ordered, multi-layered stratified epithelium. The bottom-most sheet is a tightly packed layer of proliferating stem cells attached to both the underlying matrix as well as to the cells of the terminally differentiated, postmitotic suprabasal layer. Upon differentiation, cells delaminate from the basal stem cell layer and move upwards, filling the many layers of the skin. During this upward motility, the cells need to maintain tight cell-cell junctions in order not to compromise the barrier function of this epithelium. When reaching the most superficial layer, the cells lose their nuclei and dye, forming the dead cornified layer, from where the cells are slowly sloughed off. This turnover is quite rapid, and the entire epithelium is fully renewed every 7-10 days.
The basal layer is the hotspot of most of this activity in the epidermis where some stem cells divide and continue residing within the layer while others differentiate, delaminate, move upwards, and eventually die. In the homeostatic state, epidermis must maintain a constant tissue size and this means a relatively constant basal cell number. Intuitively, the rate of proliferation then must equal to the rate of departure and delamination of the differentiating cells. On a population-level this behavior is completely logical, but the decision-making occurs on cellular level. Then, what are the drivers of cellular fate decisions within a tissue and how do cells coordinate their cell fate with their position? This is the exactly the question that we tackled in our recent publication! It turned out that it is all about differential cell and tissue mechanics!
Let the force guide you – how to robustly couple position with fate
It is intuitively apparent why one may think about external forces, such as compression and tension, in the context of the basal layer of the epidermis as it is an incredibly crowded and mechanically-loaded tissue. It is also instinctive to consider differential cellular tension in the context of cellular extrusion as we now from ample developmental biology literature that cell sorting is largely based on differential cortical tension. To put it simply, it would be more likely for a soft cell to squeeze upwards from a tightly jammed space than it would be for a stiff one. Thus, we started off with the very basic hypothesis that differential cell mechanics would be an excellent cue guiding cells to either remain within the crowded layer or change their position and move upwards. This was especially compelling in the context of highly mitotic areas with highly elevated lateral cellular compression and cell shape deformation. Our hypothesis was that crowding could alter cell shape and cortical tension. Thus, the first question that we asked was whether there was mechanical difference between cells that start differentiating versus ones that remain stem cells within the basal layer. Using atomic force microscopy we observed a biphasic evolution of cortical tension of differentiating cells, where initially upon differentiation cells indeed became softer as they made the decision to detach from the layer and, upon successful escape from the basal layer, they became stiffer, while at the same time changing their adhesion profiles. By measuring adhesion forces we further realized that the more differentiated the cell, the stronger the cell-cell adhesion forces. This was exciting as it suggested that indeed cell fate changes were accompanied by changes in cell mechanics. It further suggested that perhaps the differentiated cell was not only being squeezed out of the stem cell layer, it could also be partially pulling itself out using the strong cell-cell adhesion forces.
Time-lapse DIC video and smoothed velocity map of EPC monolayers after Ca2 + treatment. Motility vectors (left panel) and smoothed velocity maps (right panel) are shown. Asterisks demarcate examples of 2 dividing cells and arrowhead an example of a delaminating cell. Supplementary Video 3 from Miroshnikova, et al. 2018.
The path to finding answers: when physics meets cell biology
The next step was to ask the classical chicken-and-egg question of what comes first. Does the altered cell mechanics trigger differentiation or does differentiation trigger altered cell mechanics. To this end, we utilized a compression bioreactor as well as micro-patterned surfaces to precisely control crowding and cell shape anisotropy. The answers provided by these experiments made the engineers working on the project quite ecstatic as by artificially inducing crowding or restricting cell adhesion area and forcing cells to assume anisotropic shapes we were able to sufficiently to trigger differentiation. In addition, crowding was further, not surprisingly, sufficient to alter cell mechanics. So indeed, the stem cell layer crowding state seemed to control cell fate, and the cell fate change was coupled to a change in cell mechanics, providing an elegant mechanisms by which the tissue could monitor and react the changes in cell density as well as to couple cell fate to positioning.
As all of the initial observations were done using various in vitro manipulations, we subsequently turned to in vivo imaging of whole embryos to ensure that the in vitro principles are conserved and relevant to the state of the epidermis in vivo. Upon careful visual inspection of the in vivo epidermis, it was immediately apparent that the basal layer existed in a solid-like, jammed state. Interestingly, we subsequently noted that jamming coincided with the onset of delamination in our in vitro monolayers, implying that delamination of differentiating cells could be a specific feature of jammed epithelia. The biological effects of these imaging experiments were clear but their quantification was a bit more of a challenge that we overcame via a fruitful collaboration with Dr. Irene Wang and Dr. Martial Balland from Grenoble, France that allowed us to model tissue strains and monolayer flows in a quantitative manner.
Finally, it was important to test whether crowding-induced stochastic decrease of cortical tension coupled to increased cell-cell adhesion was sufficient to induce delamination. This was challenging, as any chemical or genetic perturbation would affect the mechanics of all cells, whereas we needed to induce mechanical heterogeneity. So to get an initial hint that we were really on to something relevant, we initiated an collaboration with Dr. Joerg Galle and Dr. Torsten Thalheim from Leipzig, Germany who had in the past beautifully simulated intestinal stem cell homeostasis in silico. They set out to simulate monolayer crowding, where increasing cell density through proliferation would, as shown by our experiments, reduce cell surface area, reduce cortical tension and increase cell-cell adhesion.
Time-lapse video of a 3D model epidermis simulation, side view is shown. Supplementary Video 5 from Miroshnikova, et al. 2018.
As a matter of fact, we were blown away by the results that the system maintained epidermal homeostasis at rates comparable to what had previously been observed using live imaging of intact epidermis of mice, and was completely buffered against changes in proliferation rates within the basal layer. Further, only cells with low cortical tension and high cell-cell adhesion delaminated, showing that this could indeed be an extremely robust mechanism to couple cell fate with position. We further went on to validate key aspects of the model by cell mixing experiments in epidermal organotypic cultures, where we observed that single E-cadherin-deficient cells, that showed lower cell-cell adhesion forces and high cortical tension, had severely compromised ability to delaminate, even if they had terminally differentiated.
Taken together, we conclude that individual cells tune their fates based on the activities of the neighboring cells, and by changing their mechanical properties as part of this fate switch, change their position within the tissue. This is quite remarkable, and we are now thinking about the implications of this phenomenon in terms of tissue pathologies. For example, recent findings by the lab of Dr. Phil Jones demonstrated that sun-exposed skin of healthy individuals contains clonal patches of cells with cancer-causing mutations such as mutations in Ras, but these clones co-exist with normal cells without any perturbations to the physiological functions or structure of the epidermis. Our skin is seems thus to be buffered against perturbations such as local differences in proliferative activity or differentiation rates.
I am grateful for having had the chance to work together in a fantastic team of engineers, physicists, biochemists, and cell biologists and am particularly proud to have been leading a study that employed such a wide spectrum of techniques from in vivo imaging of whole organisms to developing new quantitative algorithms and agent-based simulations to explore and understand the wonders of biology.
This editorial by Katherine Brown and Olivier Pourquié was published in Development on 22/02/18.
We are excited to announce the launch of preLights (https://prelights.biologists.com/), a new service from The Company of Biologists. preLights is a community platform for selecting, highlighting and commenting on recent preprints from across the biological sciences. With this service, we aim to help researchers discover the most interesting and relevant preprints from the growing archive of manuscripts deposited on bioRxiv and other preprint servers.
So what is the rationale behind preLights, and how will it work? While preprints have been around for a long time in the physics community, it was only when bioRxiv (https://www.biorxiv.org/) came on the scene in late 2013 that they began to take off in the biological sciences. Since then, we have seen an almost exponential growth in the number of preprints posted, as more and more researchers recognise the value in making their research available at an early stage, and as journals become increasingly open to considering papers that have already been posted on a preprint server. Here at Development, we quickly changed our policy on preprint deposition after the launch of bioRxiv, and have allowed authors to submit papers that were already available on preprint servers since Spring 2014. Since early 2016, all of the Company’s journals have been open to considering manuscripts deposited as preprints. Not only that, but we actively facilitate posting of submitted manuscripts through our bidirectional transfer portal with bioRxiv (see http://dev.biologists.org/content/news#biorxiv for more details). Overall, The Company of Biologists and Development see a synergy between preprint servers and the more traditional peer review and publishing model we operate. Preprints allow authors to get their work out quickly and readers to access the latest research, pre-publication peer review helps to ensure the rigour and quality of the work, and publication in a field-specific journal like Development helps the community find and digest the most relevant research in their area in an accessible format.
Since 2016, our community blog the Node has played an active role in helping developmental biologists find the most relevant preprints – providing a monthly round-up of the latest preprints in developmental biology and related fields (see https://thenode.biologists.com/tag/preprints/). This initiative has proved hugely popular: the post is generally our most-read post each month, and many people have told us how valuable they find the listing. Moreover, the list is getting longer each month, and is in danger of becoming unwieldy, even with the careful curation from our community manager. Given these trends, we started thinking about what more we could do to help people to navigate the ever-growing preprint literature – across the range of fields covered by the Company’s journals.
The preLighter community as of 22/02/18. Meet them all here
From these discussions, the idea of preLights was born. At its heart, preLights is a community of around 80 researchers, mainly (but not exclusively) postdocs and early stage PIs, many of whom have been nominated by our editors or editorial board members. Their research interests span the range of fields covered by our journals and beyond – from morphogenesis to neuroethology, from autophagy to cancer immunology. Our preLights team members will be selecting each month the preprint or preprints that they feel most worthy of comment, and will provide a personal perspective on why they have chosen each article. Through our online platform, we also hope to encourage other members of the community to comment on those preprints selected by our team, and to engage the authors of the selected preprints in the discussion as well – thus facilitating the exchange of ideas and opinions. Hopefully, this will also help authors improve and revise their papers as they make their way towards formal publication. Although the platform has been designed and will be hosted by The Company of Biologists, we see preLights as a community-run service, where we provide logistical support, but the content is driven by our team of selectors and the broader community.
Posts will be categorised and tagged by topic to make it easy for you to search for those preprints most relevant to you, and we’ll also highlight the most popular posts across all fields to give you a flavour of the latest work that’s attracting attention. We’re also planning to feature content from the preLights site in the journal, further helping the developmental biology community to discover the newest research in the field. And to reassure those of you who have become used to browsing the Node’s monthly list, we’re not planning to stop doing this. Finally, an additional aim is to support and promote our team of selectors – giving them the platform and profile to get their opinions ‘out there’ and helping them to expand their networks, both within their specific fields and beyond.
We are of course aware that commenting on preprints and papers has not really taken off in a big way in the biological sciences. preLights is therefore something of an experiment, but we’ve been hugely encouraged by the enthusiasm with which the idea was met by those we approached as potential contributors. While we hope that preLights will provide a venue where such discussion can happen more freely, we also believe that there is significant value in the selection and highlighting of a subset of preprints that will be of particular interest to our communities. Over time, we expect that the preLights site, and the team of selectors, will evolve – and we welcome your suggestions and feedback on how we can make it better. For now, though, we invite you to browse the first set of posts on preLights and to join in the discussion there. We hope you find this new initiative valuable!
Are you interested in applying mathematics, statistics or machine learning to biomedical problems? Apply now for a MRC WIMM Centre for Computational Biology Studentship, to start in October 2018. The studentship is fully-funded for four years, including a stipend of £18,000 p.a. and all University and College fees paid.
The studentship is open to applicants with a background in Physics, Mathematics, Engineering, Statistics or Computer Science. To be eligible for a full award, applicants must have no restrictions on how long they can stay in the UK and must have been ordinarily resident in the UK for at least 3 years prior to the start of the studentship. Further details about residence requirements may be obtained here.
For further information on how to apply can be found here.
All applications must be received by 12 noon (UK time) on Friday, 23 March 2018.
Interviews will take place the week commencing 16 April 2018.
Approaching the problem of muscle disease with cell replacement therapy
We will all be affected by a muscle wasting disease in our lifetime. The ability to transplant a stem cell into skeletal muscle that can self-renew to make more of itself, and respond to injuries as they arise, could provide a continuous source of cells to replace damaged muscle tissue throughout our life span.
Stem cell replacement is especially imperative to the largest childhood muscle disorder Duchenne Muscular Dystrophy, where a genetic mutation in muscle cells results in muscle death and progressive weakness for about 20,000 new cases each year worldwide. The muscle stem cells of Duchenne patients also contain the genetic mutation rendering them ineffective at repairing the muscle as it dies.
Human pluripotent stem cells (hPSCs) have the potential to differentiate into any cell type making them attractive candidates for drug screening and cell replacement therapies. Indeed, hPSCs are already being used in the clinical setting and in preclinical models for diseases ranging from macular degeneration to cardiac myopathy, diabetes, and Parkinson’s disease.
Using induced pluripotent stem cells taken from Duchenne patients, our team at UCLA has devised a strategy to permanently correct the genetic mutation in Duchenne cells with CRISPR/Cas9 (Young et al., 2016). Our current challenge is delivering the corrected cells back to the affected patients. Correction of patient-derived induced pluripotent stem cells ex vivo and subsequent differentiation to human muscle stem cells offers one potential route for restoration of dystrophin after transplantation.
Pluripotent stem cells must mature through stages of development similar to natural cells to function like in vivo cell counterparts
When we first started this project, the only way to make skeletal muscle from human pluripotent stem cells was using virus to overexpress muscle transcription factors, which lacks precision in truly recapitulating the cells created from human development. For example, during regeneration a muscle stem cell must be able to simultaneously make more of itself and differentiate to make new muscle fibers. Forced viral overexpression of genes may limit proper stem cell dynamics.
Directed differentiation is a process that follows developmental cues to allow hPSCs to form any cell lineage. It has been well established that for many cell lineages, including cardiac muscle cells, the progeny made from directed differentiation of hPSCs are immature and do not function as well as adult cells. While there are a number of recent papers that use directed differentiation to produce muscle cells from hPSCs, no papers have benchmarked the cells being made to those derived from human muscle tissue. In this study we set out to follow human development of muscle more closely to create a muscle cell similar to those naturally found within the body as a means to obtain a cell for replacement therapy.
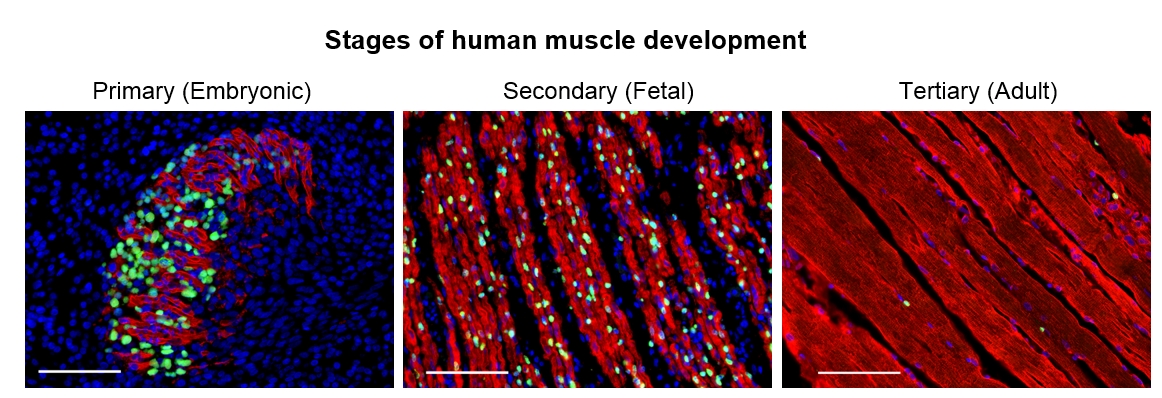
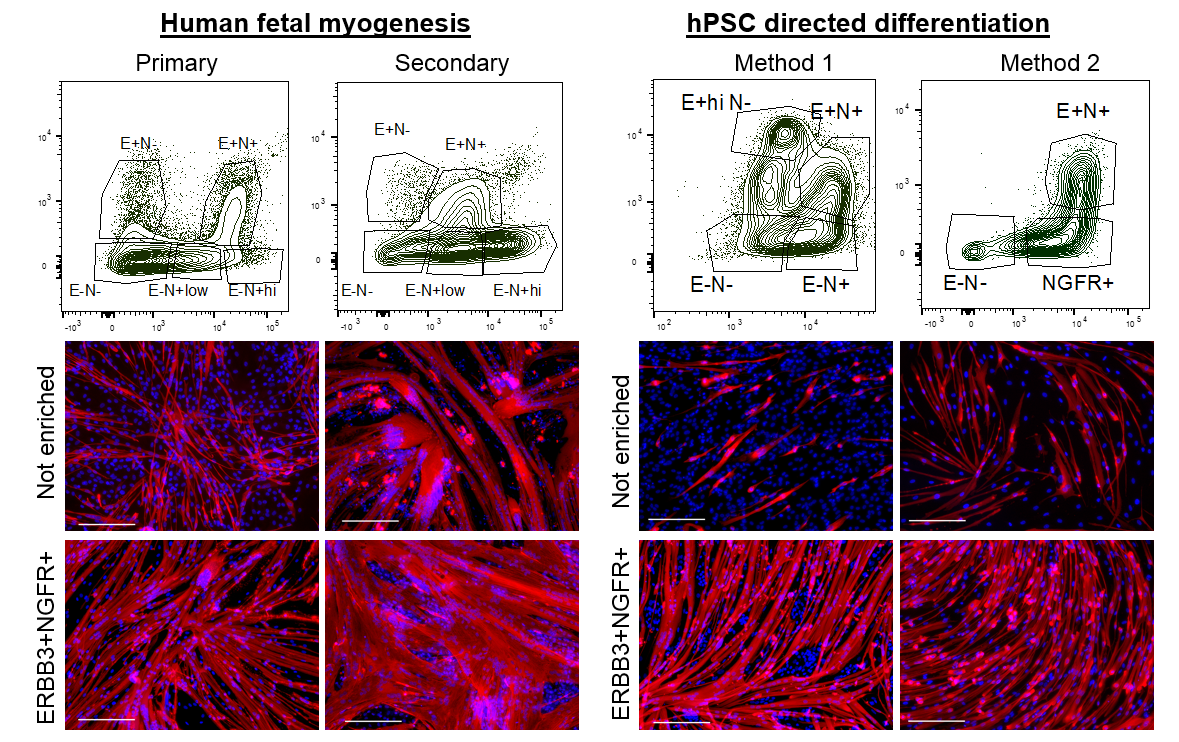
Figure 1. Human developmental myogenesis occurs in three successive waves, each fulfilling different functional needs. Primary myogenesis establishes the basic muscle pattern. Secondary myogenesis is critical for growth and maturation of the muscle. Adult myogenesis allows for postnatal growth and repair of damaged muscle. Myosin staining (Red) shows the differences in myofiber morphology at each stage of human development. DAPI staining (Blue) shows the number of nuclei within and outside of myofibers. While myogenic transcription factors such as PAX7 (Green) are expressed by both skeletal muscle progenitor cells (SMPCs) in fetal development and by satellite cells (SCs) in adult, the number of SMPCs and SCs per myofiber and ability to make new myofibers may widely vary. Scale bar equals 100μm.
We set out to determine the myogenic potential of muscle cells created from hPSCs (termed skeletal muscle progenitor cells or hPSC-SMPCs), compared to the muscle cells from human embryonic, fetal, and adult myogenesis (Figure 1). Cells were dissociated from human tissues and their ability to fuse to form myotubes were compared to hPSC-SMPCs. We found that hPSC-SMPCs made from two commonly used directed differentiation protocols created muscle in a dish that looked and fused similarly to the embryonic muscle from primary myogenesis.
When we transplanted hPSC-SMPCs into mice, we found they did not engraft very well. Alternatively, human fetal cells from secondary myogenesis were very capable of forming new muscle in vivo.
Directed differentiation to skeletal muscle produces a number of heterogeneous cell types, and we thought that enrichment for the best-known adult satellite cell marker, known as NCAM or CD56, would improve our ability to engraft cells. However, NCAM enrichment failed, and if anything was slightly worse than unsorted hPSC-SMPCs. This was an example of the failure of applying adult tissue paradigms to immature hPSC progeny from directed differentiation. It was like comparing apples and oranges. After many months, we were no closer to making an engraftable cell from hPSCs.
Development guides the enrichment of functional muscle progenitors from hPSCs
We thought about how muscle cells created from hPSCs functionally resembled the morphology of muscle from embryonic myogenesis, but how muscle cells from fetal myogenesis were able to engraft very well. We hypothesized rather than trying to immediately create an adult cell from hPSC-SMPCs, we still had to direct their differentiation through more stages of human myogenesis and maturation.
The first thing we needed to do was determine the maturational identity of our cells. We performed RNA sequencing of NCAM+ hPSC-SMPCs, and compared gene expression to NCAM+ fetal week 17 SMPCs. As predicted, we found that hPSC-SMPCs were more embryonic than fetal SMPCs, but they were also more differentiated with increased expression of genes such as myogenin. This genetic profile supported the idea that the cells we were making more closely resembled the primary myogenic wave in which founder cells differentiate to establish the first muscle fibers, before a second wave of muscle progenitors arise that go on to make the adult satellite cells. Because NCAM is expressed by many cells in directed differentiation, including immature mesoderm cells, neurons, and more differentiated myoblasts, we hypothesized this was also a reason for the lack of improved enrichment and engraftment in vivo by NCAM+ hPSC-SMPCs.
We turned to our colleagues, Drs. Denis Evseenko and Ben Van Handel at the University of Southern California, who were facing a similar challenge with directed differentiation of hPSCs to chondrocytes. To identify unique genes of human chondrocytes, Dennis and Ben had performed a Weighed Gene Analyses (WGCNA) on five types of human fetal tissues including bone, cartilage, tendon, ligament, and skeletal muscle. In the fetal muscle dataset, they identified over 400 cell surface genes and transcription factors unique to skeletal muscle (Furgeson et al., 2018). A fun part of this project was pilfering through the gene data, making candidate lists, performing PubMed searches and refining cell surface genes to about 20 candidates. We then set up directed differentiations to evaluate new and known cell surface marker expression from the candidates in hPSC-SMPCs. From this screen, we found enrichment for ERBB3 or NGFR produced the most myogenic cell in vitro with robust fusion ability.
Eureka! ERBB3 or NGFR demarcate myogenic cells in human development
We wondered about the relevance of ERBB3 or NGFR in human development and whether these two cell surface markers demarcated the same cell or different cells at stages of development. We decided to analyze and enrich for cells based on ERBB3 or NGFR co-expression in primary and secondary myogenesis. We identified multiple subpopulations based on the expression profiles of these two cell surface receptors. In all cases, double positive ERBB3+NGFR+ cells resulted the highest PAX7 gene expression. Furthermore, primary and secondary myogenesis could be distinguished based on these markers, where very high expression marked a more immature progenitor (Figure 2). ERBB3 was not expressed by adult satellite cells.
We next applied this same enrichment strategy to hPSC-SMPCs. As expected, in every directed differentiation protocol tested double positive ERBB3+NGFR+ SMPCs could be induced to form homogeneous myotubes. These data demonstrate a striking similarity between the surface marker phenotypes of fetal myogenic progenitors and hPSC-SMPCs undergoing directed differentiation.
Figure 2. ERBB3 and NGFR mark a distinct skeletal muscle progenitor cell in human development and hPSCs. Top. Using flow cytometry, subpopulations of human myogenic cells were identified based on ERBB3 and NGFR expression during primary myogenesis (fetal weeks 8–10), secondary myogenesis (fetal weeks 16–18), and two hPSC directed differentiation protocols (Shelton et al., 2014 and Chal et al., 2015), Bottom. When fetal or hPSC-SMPCs were dissociated/replated or enriched for ERBB3+NGFR+ and induced to differentiate to myotubes, nearly 100% ERBB3+NGFR+ progenitors fused to form MYHC+ (Red) myotubes
Development guides the maturation of muscle progenitors from hPSCs
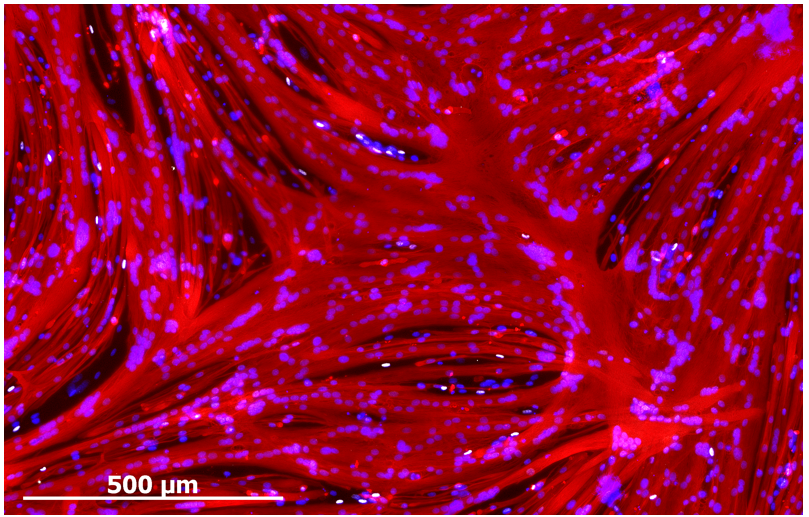
Despite enabling myogenic enrichment, differentiation of ERBB3+NGFR+ populations isolated from hPSCs yielded myotubes that were thinner and contained fewer nuclei than later stage fetal or adult counterparts. To understand the basis for these differences, we further analyzed our RNA sequencing data comparing hPSC-myotubes and fetal myotubes to their progenitors. These data revealed that activators of TGF-β signaling decreased during the course of fetal myotube differentiation. In contrast, hPSC-SMPCs had higher expression of TGF-β signaling genes and hPSC myotubes failed to down regulate TGF-β signaling, such as myostatin. When we blocked the TGF-β signaling we significantly increased hPSC-myotube fusion and produced morphology similar to late stage fetal myotubes (Figure 3). We performed multiple assays to verify that TGF-β inhibition indeed matured hPSC-myotubes.
Figure 3. TGF-β inhibition during differentiation of ERBB3+NGFR+ hPSC-SMPCs generates multinucleated (DAPI, blue) myotubes (Myosin, Red) that resemble late-stage fetal or adult myotubes in vitro. Staining also show the presence of PAX7+ cells (white) at the periphery of myotubes
ERBB3+ hPSC-SMPCs can generate new muscle fibers upon transplantation in vivo.
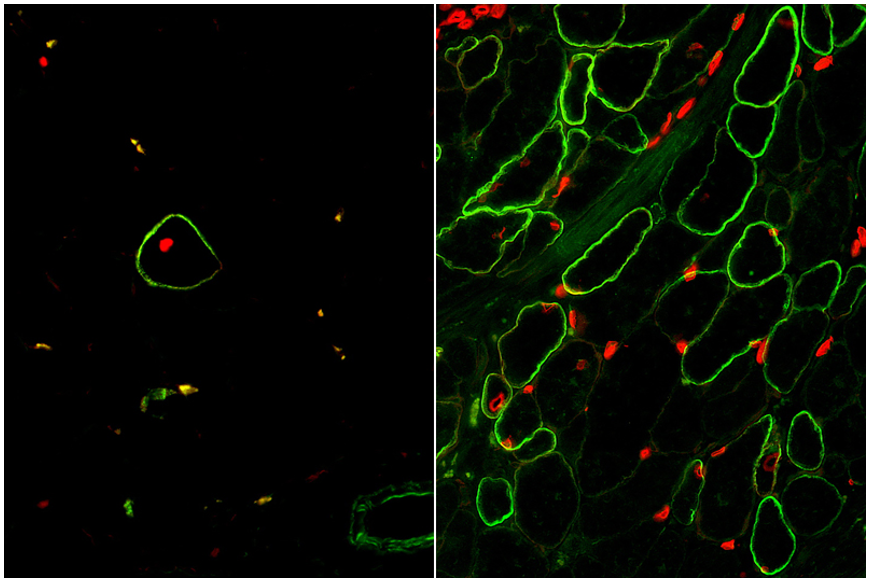
The moment of truth had arrived when our enrichment and maturation strategy was applied to hPSC-SMPC engraftment in vivo. After 30 days of waiting, mice injected with ERBB3+ hiPSC-SMPCs and treated with TGF-βi were evaluated for restoration of human dystrophin in a mouse model of DMD. We were gratified to find a greater than 50-fold improvement in number of dystrophin+ fibers using our directed differentiation strategy (Figure 4). This was especially satisfying because these cells were originally taken from boys with Duchenne muscular dystrophy, reprogrammed to induced pluripotent cells, and genetically modified with CRISPR/Cas9 to permanently correct the DMD gene mutation. What’s more, ERBB3+ hiPSC-SMPCs enabled engraftment approaching levels equivalent to directly-isolated human fetal muscle cells. By following human development, our results demonstrated a major advance in the ability to obtain engraftable SMPCs from hPSCs.
Figure 4. Skeletal muscle cells isolated using the ERBB3 and NGFR surface markers (left) restore human dystrophin (green) after transplantation significantly greater than previous methods (right).
The next phase of this research will be to test and develop strategies to mature SMPCs to satellite cells similar to adult, which can reside in the stem cell niche in vivo and continuously repopulate new muscle after damage. We hope to continue to create a more regenerative cell for use in cell replacement therapy for patients with DMD and many other muscle wasting diseases.
An exciting opportunity to be part of a pioneering biomedical research institute, dedicated to innovation and science. A postdoctoral research post in the laboratory of Dr. Alex P. Gould is now available. We are looking for a highly motivated researcher to identify the, as yet unknown, molecular mechanisms by which dietary nutrients and oxidative stress during development can profoundly alter adult metabolism and longevity. The successful applicant will choose which of the Drosophila and/or mouse model(s) established in the lab they wish to focus on (PMID: 21816278, PMID: 26451484, PMID: 29123106 and unpublished). Research techniques used in the lab include genetics, molecular biology, confocal microscopy, biochemistry, metabolomics and mass spectrometry imaging. Details of projects currently being undertaken in the lab can be seen at: THE GOULD LAB. The successful applicant will have good communication and organisational skills and a PhD in a relevant area (or be in the final stages of completion). Informal enquiries to alex.gould@crick.ac.uk.
The Francis Crick Institute is a biomedical discovery institute dedicated to understanding the fundamental biology underlying health and disease. Its work is helping to understand why disease develops and to translate discoveries into new ways to prevent, diagnose and treat illnesses such as cancer, heart disease, stroke, infections, and neurodegenerative diseases. An independent organisation, its founding partners are the Medical Research Council (MRC), Cancer Research UK, Wellcome, UCL (University College London), Imperial College London and King’s College London. The Crick was formed in 2015, and in 2016 it moved into a new state-of-the-art building in central London which brings together 1500 scientists and support staff working collaboratively across disciplines, making it the largest biomedical research facility in one building in Europe. The Francis Crick Institute will be world-class with a strong national role. Its distinctive vision for excellence includes commitments to collaboration; developing emerging talent and exporting it the rest of the UK; public engagement; and helping turn discoveries into treatments as quickly as possible to improve lives and strengthen the economy.
” If you are interested in this role, please apply via our website.
” The closing date for applications is Monday, 19 March 2018 at 23:30.
” All offers of employment are subject to successful security screening and continuous eligibility to work in the United Kingdom.
Key words
” Developmental biology
” Mouse genetics
” Drosophila genetics
” metabolism
” physiology
” developmental origins of health and disease
” mass spectrometry imaging
” metabolomics
” cell biology
” molecular biology
The droso4schools initiative is an educational outreach programme run by the Manchester Fly Facility, which I had the opportunity to be actively involved in during a month-long placement (see my previous blog). The aim of this programme is to re-introduce the fruit fly Drosophila melanogaster into school and college biology lessons. For this, the droso4schools team have developed a number of hands-on, readily prepared, interactive lessons with adjunct materials, which explain curriculum-relevant biology contents at the level of Key Stages 4 – 5. As has been explained in a recent blog and publication, these lessons make active use of Drosophila as a teaching tool – both to explain and illustrate contents and to perform micro experiments.
The Manchester Fly Facility team recently hosted a continuing professional development (CPD) day for a group of Key Stage 3, 4 and 5 science teachers, to showcase their existing lesson packages (and more are in production…). This was an important opportunity for the droso4schools team to explain their resources and gain feedback from a focus group of experienced teachers – namely, what are the lessons like to use? Could they be improved? Most importantly, are they curriculum-relevant?
The day began with a tour of the fly facility, a short activity using microscopes to identify common phenotypic markers used in Drosophila research, and an introduction to the droso4schools programme by academic lead Professor Andreas Prokop, and long-time collaborator Dr Catherine Alnuamaani, a teacher and keen Drosophila advocate from Trinity CoE High School, Manchester. Andreas and Catherine together explained the mutual benefits of using Drosophila as a teaching tool: the benefit for pupils and teachers lies in the fact that Drosophila provides uniquely detailed biology knowledge, delivering concepts, stories and experiments across biology – paired up with human examples to illustrate relevance. Researchers benefit in that students are introduced early on to the power of research with genetic model organisms as a key driver for scientific discovery, and will then hopefully be more open to such research strategies when entering university.
As Catherine highlighted, teaching practicals can become a little monotonous, and the experiments themselves are rarely very exciting. Using a live animal in the classroom puts the “life” back into the life sciences, and encourages pupils to engage with the subject content – Drosophila brings real-world relevance to the curriculum. Not only that, it also provides an opportunity for teachers to develop experimental skills, in a way that is both cost-effective and more interesting than starch-storing potatoes. But what benefit is there for researchers? As Andreas explained, many students who arrive at university to study life sciences, often don’t have much of an interest in invertebrates – they are attracted to applied vertebrate biology and clinically oriented research. Of course, this kind of research is important, but it is fundamental biology which forms the basis on which “translation” can occur. By demonstrating to school pupils the potential scope of fruit fly research, it enhances the chance that those who choose to study the life sciences at university, will have more of an appreciation for the impact of Drosophila research on modern science, and be keen to explore the field themselves.
The itinerary for the day included three full lessons in which the participating teachers took part from the student perspective. Each lesson consisted of a well-structured PowerPoint presentation guiding through the contents, interspersed with short activities and micro experiments. A couple of the participants had been using flies from the facility in their lessons already, mainly as a means of demonstrating genetic crosses; one had even been using elements from the “alcohol lesson” (see below), with great success, for two consecutive years. These teachers reported that students loved having live animals in the classroom, and so they had come to explore other options. The rest of the group were relatively new to the idea of Drosophila as a teaching tool. Some knew how they’d like to use the flies, but weren’t sure where to start. Others were curious, but not sure how successful flies could be as a learning resource, and how easy it would be to source materials.
Being in a classroom full of teachers is something of a surreal experience, but everyone was more than happy to get stuck in, ask questions, work in groups, and contribute to class discussions. The first lesson, concerning the fundamentals of nervous system organisation and function (“Principles of the Nervous System”), gave the participating teachers their first opportunity to work with live flies, through a series of micro behavioural experiments involving the shaking of epileptic flies into seizure (an experiment that is used to illustrated wiring principles and the mechanism of nerve impulses), and another causing transient paralysis of shibire mutant flies (which involves keeping vials of flies under your armpits for five minutes … always popular with pupils!).
Lesson two (“From gene to enzyme to evolution: using alcohol metabolism to illustrate fundamental concepts of biology”) is a fantastic revision lesson to use before the A-level exams, lining up a broad range of specifications, from gene expression to enzymes to evolution, into a consistent story. It contains a particularly hands-on practical, studying enzymatic reactions using the dissection of Drosophila larvae followed by a 5 minute staining reaction, which was very well received by the participants. This practical also provides a means to demonstrate the potential impact alcohol consumption can have on health, providing opportunities to discuss this topic most relevant to students of that age.
The third lesson (“Our vision: understanding light and light perception”) was of particular interest to me, as I had worked on its experimental component during my placement at the Manchester Fly Facility. I used the fact that flies are attracted to light (an innate behaviour called positive phototaxis) to test responses to different wave-lengths, using normal and sevenless ‘colour blind’ flies. This experiment is particularly effective, as taxis behaviours are simple automatic responses to a given stimulus, and therefore easy for pupils to identify and observe. I had some great feedback from the teachers, a number of whom commented on the short fly behaviour video, and how easy it would be for pupils to engage with – which was very rewarding! Most importantly, it was later highlighted that the observation of taxis behaviours is now a compulsory part of the AQA A-Level biology practical syllabus – so if any teachers are reading this, it is worth taking a look at this lesson!
A brief overview of another lesson (“The climbing assay: learning data analysis through live experiments with fruit flies”) was received with particular enthusiasm by everyone and sparked imaginations. This lesson makes use of a climbing assay that compares motor activities of young versus old flies to perform statistical analyses. Many of the teachers seemed quite excited to use this particular lesson with some of their younger (KS3) classes, as well as for the GCSE and A-Level students, as it is so easily translatable. Not only would it be straightforward to teach, but all agreed it would be a great way to engage pupils with what is traditionally not the most interesting part of scientific methodology.
To complement the lesson sessions, Sanjai Patel (manager of the Manchester Fly Facility) provided a short masterclass on basic fly handling and care techniques, in which teachers were given flies and had to tip them over into fresh vials. Everyone seemed to get the hang of it quite quickly, and there were hardly any escapees! Finally, teachers were given a “guided tour” through the facility’s droso4schools website (describing the resources and given additional support information), the teachers’ section of the Manchester Fly Facility website, and the figshare.com repository providing all the teaching resource files for download.
At lunch and during the concluding session of the day, there was a chance to consolidate the day’s information, reflect on the lessons, share thoughts and opinions on the ease of using Drosophila as a teaching tool, and provide feedback to the team. It was very inspiring, and also in many ways reassuring, to talk to teachers from different schools and areas, who were similarly passionate about their career choice, and their roles as science advocates. During the discussion, a number of excellent ideas were raised, and it was great to see how enthusiastic all the participants were about the project, and how keen they were to get back in the classroom and try the lessons for themselves! Positive feedback included:
how easily adaptable all the resources were – images, lessons, and other materials can be downloaded and utilised separately, or in whatever combination is convenient.
how easy it was to link the Drosophila practicals to current research, providing that real-world edge so often missing in everyday lessons.
that the lessons provide an excellent resource for pupils practicing the new synoptic essays (a synoptic essay assesses a pupil’s ability to demonstrate and apply their understanding of a topic, using a variety of examples on a given theme); often, this additional knowledge is obtained outside of the classroom, through private study.
that the depth and breadth of information available about Drosophila is easy to incorporate as “additional” information into revision lessons.
An area for improvement mentioned by some of the participants, was that there was something of a “language barrier”, i.e. the differences in basic terminology used in research, compared to that used in schools. Other barriers mentioned were the complex technicalities of the GSCE and A-level curriculum. As a potential solution to tackle these barriers, it was suggested that an experienced teacher could become involved with the droso4schools programme, to “fine-tune” the resources. As it was put by one participant: “You are almost there, but it now needs someone with A-level experience to fully adapt them for school use”.
Overall, the day was very uplifting, and all seemed to head for home brimming with new ideas – be it how to introduce flies to their lessons, or how to further improve available resources. Science lessons should never be boring – neither for pupils nor for teachers. Introducing Drosophila to the classroom provides a new and exciting way to observe science in action, in a way which is convenient, affordable, and manageable. Enthusing pupils about the possibilities presented by research with this powerful invertebrate could also help to inspire a new generation of fundamental life scientists and, who knows, maybe even a few more Nobel Prize winners!
Vertebrate segmentation involves the periodic formation of somites from the presomitic mesoderm, in a manner controlled by oscillating gene expression (the oscillations of the segmentation clock must be one of the marvels of nature!). While in vivo work has provided a framework for studying the process, many aspects of segmentation dynamics are obscured in the embryo. A new Techniques & Resources paper in Development describes a stem cell-derived in vitro system for studying segmentation dynamics. We caught up with first author Marina Matsumiya and her supervisor Ryoichiro Kageyamaof Kyoto University to hear more about the story.
Ryoichiro and Marina
Ryoichiro, can you give us your scientific biography and the questions your lab is trying to answer?
RK I became Assistant Professor of Kyoto University in 1989 and started a project about transcription factors that regulate mammalian neural development. We reported the basic helix-loop-helix (bHLH)-type repressor genes Hes1, Hes3, and Hes5 in 1992 and the bHLH-type proneural gene Atoh1 (Math-1) in 1995, and since then we have been characterizing the functions of these genes in neural development. We found that the antagonistic regulation between Hes and proneural genes controls neurogenesis and gliogenesis.
In December 1997, I was promoted to full professorship in the same university and continued the bHLH project. In the same month, I met Olivier Pourquié, when he visited my group in Kyoto, and we had discussion about his finding (just published in Cell) that the expression of chick hairy1, a Hes1 homologue, oscillates during somite segmentation. This discussion made me keep pondering on how such oscillatory expression with ultradian rhythms is possible and how such dynamic expression regulates the downstream events. In the 2000s, we reported that Hes1 expression oscillates in many cell types by negative feedback, and that this oscillation is important for cell proliferation. In parallel with this finding, we identified a new member of the Hes family, Hes7, and found that its expression oscillates by negative feedback in the presomitic mesoderm (PSM), thereby regulating the somite segmentation. These findings brought us to the field of the segmentation clock. Hes7 oscillations are synchronized under the control Notch signaling, and the oscillatory expression of the Notch ligand Delta-like1 is involved in the synchronized oscillations. However, the detailed mechanism of the synchronized oscillation with traveling waves in the PSM is still unknown, and we have been trying to answer this key question.
What is the status of developmental biology research in Japan?
RK Developmental biology research is well funded in Japan if a stem cell or regenerative research field is included. However, besides this field, it seems to be more difficult to get sufficient funding. I think that translational research-oriented funding is a general trend in Japan, as observed world-wide. Collaborations are very active and actually essential to many research projects of developmental biology. Grants supporting domestic and international collaborations are also available. In addition, Riken offers a service of generation of transgenic mice as a collaboration basis, and many researchers utilize this service. I do not know the exact number of labs in developmental biology in Japan, but the Japanese Society of Developmental Biologists (JSDB) has about 1200 members. This is a good number, compared to The Society of Developmental Biology (USA), which has nearly 2000 members.
However, I am a bit concerned about the recent trend. There used to be more than 300 poster presentations in the Annual Meeting of the JSDB, but last year, there were only 193, suggesting that the number of young researchers in the field is decreasing over the years. One reason is that there were only a handful of poster presentations of the stem cell or regenerative research field in the Annual Meeting of the JSDB, while many Japanese researchers participated in the ISSCR in Boston last year. I feel that (classic) developmental biology and (fancy) stem cell or regenerative research should go hand-in-hand, but in reality these two fields seem to go separately in Japan. It would be desirable to incorporate more stem cell researchers into the JSDB meetings to advance the field properly.
And Marina, how did you come to be involved with this project?
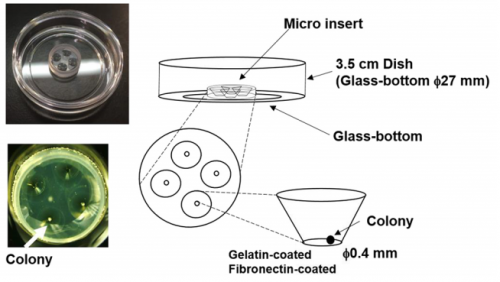
MM I was interested in the mechanism of somitogenesis, so I joined the Kageyama Lab as a master student. There were no PSM-like cell culture systems, and my colleague, Kumiko Yoshioka-Kobayashi, suggested me that such systems would be important to examine the detailed mechanism of somitogenesis. After discussion with Kumiko and RK, I decided to make a PSM-like cell culture system. At almost the same time, I found that Olivier’s group published a method to induce PSM-like cells from mouse embryonic fibroblasts (MEFs), so we first used MEFs produced from our Hes7 reporter mice. However, we were not able to induce Hes7-expressing cells, and we next tried mouse embryonic stem (ES) cells following the paper from the same group published in 2015, which focused on the induction of muscle fibers from ES cells. We also had a chance to learn an ES cell culture method from Mototsugu Eiraku at Riken CDB, who established a method to induce the eye cup from ES cells. By combining these methods, we succeeded in inducing PSM-like cells, which exhibited Hes7 oscillations.
Matsumiya, et al.’s culture method for imaging Hes7 expression in iPSM colonies, from Figure 1 in the paper.
What are the difficulties of studying the segmentation clock in embryos? And what were the limitations of previous in vitro approaches?
RK Detailed analysis of the segmentation clock has been hampered because it requires the use of live embryos. Analysis involves both genetic and pharmacological approaches: in genetic approaches, mutant animals are generated to activate or inactivate gene functions, while in pharmacological approaches, either embryos or PSM tissues prepared from embryos are used for treatment with chemicals that modulate gene activities. If in vitro cell cultures are available, both genetic and pharmacological analyses would be facilitated, and many attempts have been made to induce PSM-like tissues from ES cells. Although PSM-like tissues have been successfully induced from ES cells, there are no reports of wave-like propagation of oscillatory gene expression in such induced tissues. So, we started the in vitro PSM cell project.
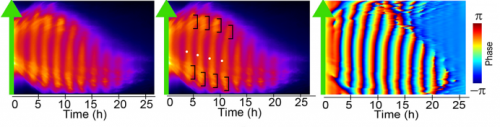
Hes7 expression in an iPSM colony cultured in a fibronectin-coated plate; Movie 2 from the paper.
Can you give us the key results of the paper in a paragraph?
MM & RK We established a simple and efficient method to generate mouse ES cell-derived PSM-like tissues, in which Hes7 expression oscillates like traveling waves. In these tissues, Hes7 oscillation is synchronized between neighboring cells, and the posterior-anterior axis is self-organized as the central-peripheral axis, generating somite-like segments at the periphery. This method is applicable to not only chemical library and RNAi screening but also CRISPR-Cas9-mediated gene modifications and will facilitate the analysis of the molecular nature of the segmentation clock.
In your system, Hes7 oscillations did show some variability: do you know what underlies this variability?
MM & RK There is some variability in the amplitude, period, and patterns of Hes7 oscillations even in the same iPSM colonies, which may hamper to detect effects of chemicals and genetic modifications. This variability may depend on the size and shape of iPSM colonies, and how iPSM colonies expand on dishes. Further improvements will be required to reduce such variability.
Kymographs of Hes-7 driven luciferase, from Figure 3 in the paper.
Is your system restricted to mouse cells, or could you use it to compare segmentation between species?
MM & RK We have not examined other species yet, but we are sure that this method may be applicable to ES/iPS cells of other species. We are interested in the mechanism of how species-specific periods of Hes7 oscillations are determined.
When doing the research, did you have any particular result or eureka moment that has stuck with you?
MM My eureka moment is when I observed Hes7 oscillation in iPSM colonies for the first time. I still remember the date, and we used the data in Figure 4 of the paper. We did not get nice data for a few years, but after changing the culture conditions, all of sudden, everything went well. We were surprised to see the Hes7 oscillations propagating like waves and leading to somite-like segment formation. When I saw the movie, we celebrated with high five. I could watch it forever. It took 6 months from the first detection of Hes7 oscillation to submit the paper. Every day in this half a year was a series of joy.
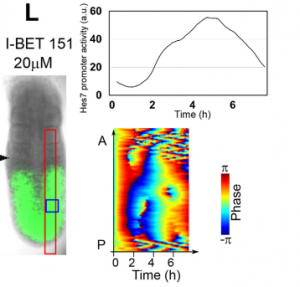
Validating in vitro results in the embryo, from Figure 7 in the paper
And what about the flipside: any moments of frustration or despair?
MM When I used MEFs, I was really nervous. I got some Hes7 signals but not oscillatory expression. I tried many different culture conditions, but none of them worked. It was like a never-ending nightmare, but now I feel that it is a very important step to improve myself mentally and technically.
What next for you Marina?
MM I have one more year to go before I finish my PhD course. Until then, I will try to reveal new factors or pathways that make Hes7 oscillations synchronized by using the iPSM method. This information will be important for the somitogenesis research. After finishing my PhD course, I would like to do research on refractory diseases of bone formation related to somitogenesis. Now, I will prepare the application of a fellowship to become a postdoc.
Where will this work take the Kageyama lab?
RK By using this system, we now started screening many chemicals to find a new pathway involved in synchronization of Hes7 oscillations. We also plan to use two-color fluorescent reporters (one from Hes7 promoter and the other from another important PSM gene) to examine the oscillation at the single-cell resolution. We are trying to find chemicals that do not repress Hes7 expression but affect synchronization. These chemicals, if found, would help reveal the mechanism of synchronized oscillation besides the Notch pathway. We have recently developed an optogenetic gene induction system and plan to introduce this system to iPSM cells. This system allows precise spatiotemporal control of gene expression, so we would like to see how light-controlled gene expression affects Hes7 oscillations.
Finally, let’s move outside the lab – what do you like to do in your spare time?
MM I would like to sleep, but I also like playing modern ballet. These are very important for me to control my condition.
RK I like jogging along the Kamo river in Kyoto, which really refreshes me. I also like walking. I used to bike, but after an accident I stopped biking now.
In spite of the difference between the cell functions responsible for giving rise to a tumour and for the metastasis of this same tumour, studies at IRB Barcelona using the fly Drosophila melanogaster reveal that some genes can drive both phenome
na.
In cancer research, there has been a tendency to differentiate between the genes responsible for tumour development and those that give rise to metastasis and to study these processes separately since the cell functions required for these two mechanisms differ radically. Researchers headed by Jordi Casanova at the Institute for Research in Biomedicine (IRB Barcelona) now demonstrate that genes activated during metastasis are also able to initiate primary tumour development, and they explain the molecular mechanism involved. Made using the fly model Drosophila melanogaster, this finding has been published in PloS Genetics this week.
“The results strengthen the notion that was already being put forward in cancer research,” explains Jordi Casanova, head of the Development and Morphogenesis in Drosophila lab and CSIC professor. He goes on to say “And that is that the same genes can activate tumour growth and metastasis, and therefore some tumours are conferred metastatic capacity from their very outset”.
Two phenomena that share genes
The study is based on the observation that benign human tumours or tumours at stages well before metastasis express some genes associated with this process. “This overlap made us question whether the so-called epithelial-mesenchymal factors—which activate genes associated with cell movement and migration, which are common in embryo development and are also present in metastasis—play a role in tumour initiation,” says Casanova.
In particular, GATA genes are overexpressed in liver metastasis in colon cancer patients, as well as in several human tumours, such as those of the breast, ovary, pancreas and colon. Jordi Casanova’s team examined various genes, among them Serpent (homologue of GATA in Drosophila), and demonstrated that the overexpression of this gene is indeed sufficient to drive tumour growth. In this regard, this gene causes overgrowth of the wing, modifies tissue structure, and activates a series of genetic changes in order to facilitate initial tumour growth.
The fly wing as a model of epithelial tumour
The research was conducted on the fruit fly Drosophila, an established model for studying epithelial tumours. “Once again, studies on Drosophila have contributed to our basic understanding of the behaviour of genes involved in cancer and metastasis, knowledge that will allow cancer researchers to pursue this line further,” says the scientist.
Kyra Campbell, first author of the study, former postdoctoral fellow at IRB Barcelona and currently at the University of Sheffield, in the UK, was supported by Xavier Franch-Marro, at the CSIC’s Institute of Evolutionary Biology.
This work was supported by the Ministry of the Economy and Competitiveness, through ERDFs, and by the Government of Catalonia.
The Beddington Medal is the BSDB’s major commendation to promising young biologists, awarded for the best PhD thesis in Developmental Biology defended in the year previous to the award. Rosa Beddington was one of the greatest talents and inspirational leaders in the field of developmental biology. Rosa made an enormous contribution to the field in general and to the BSDB in particular, so it seemed entirely appropriate that the Society should establish a lasting memorial to her. The design of the medal, mice on a stylised DNA helix, is from artwork by Rosa herself. We would like to congratulate the 2018 winner of the Beddington Medal, Emilia Favuzzi, and would like to take this opportunity to give a brief overview of her career and her PhD project that was awarded the Beddington medal.
Emilia’s career and project
Emilia started her studies in 2007 at the Sapienza University of Rome and was awarded a B.Sc. in Biological Sciences with highest marks in 2010. She stayed at the same university for her Master’s project which she performed in the laboratory of Sergio Nasi at the Institute of Molecular Biology and Pathology (CNR, Rome). She completed her M.Sc. in Neurobiology in 2011, again with highest marks. In 2011 she joined the group of Beatriz Rico at the Institute of Neuroscience in Alicante (Spain) and moved with that group to the Centre for Developmental Neurobiology at King’s College London in 2014 where she terminated her project work. Her PhD in Neuroscience was awarded in 2017 by the University Miguel Hernandez of Elche (Spain) also with summa cum laude. Since 2017 she is a postdoctoral associate in Gordon Fishell’s laboratory at the Broad Institute and Harvard Medical School.
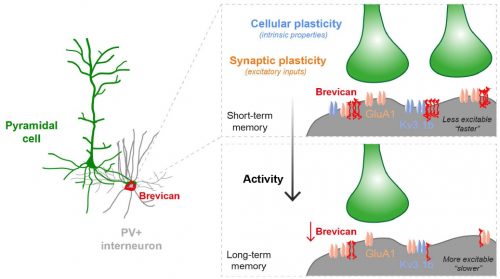
Fig.1 Activity-dependent gating of parvalbumin interneuron function by the perineuronal net protein Brevican
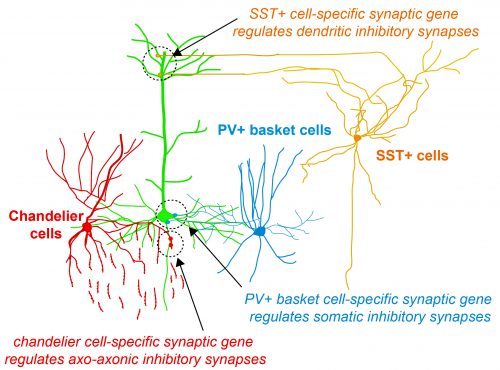
During her PhD, Emilia worked on two projects which were both based on candidate and genome-wide screen approaches aiming to identify genes that were involved in GABAergic synapse formation. In one project, she investigated the role of perineuronal nets during the synaptic development of GABAergic interneurons. She discovered that the perineuronal net component Brevican is involved in the gating of parvalbumin interneurons by controlling their intrinsic properties as well as extrinsic input through excitatory synapses (Fig.1). This paper was published as a featured article in Neuron (2017). Emilia also took ownership within a parallel project, where she collaborated with another lab member to set up protocols to isolate different populations of interneurons and screen for genes involved in the specific synaptic targeting of cortical interneurons to the different compartments of pyramidal cells. This work led to the discovery of validated candidate genes involved in specific interneuron synapse formation, as shown via loss and gain of function approaches (Fig.2). The respective manuscript is in preparation and Emilia will be shared first author.
Her PhD supervisor Beatriz Rico said about her: “Emilia is a gift for a supervisor: she goes ahead of you, technically and conceptually and pushes you forward. She is brilliant, extremely motivated and creative person and resistant to any difficulties she has found during the development of her project. She never gave up and pursues her aims with an impressive efficiency. She is extremely independent and hard worker. She is fully committed to science, a dream for a supervisor.”
Thesis abstract: Cell-type specific programs regulate the assembly and dynamics of cortical circuits
Understanding how neuronal connections are established and organized in functional networks during development is critical to understand brain function. In the mammalian cortex, GABAergic interneurons are characterized by a remarkable diversity of types and connectivity patterns. As such, they are uniquely suited to orchestrate functionally relevant circuit-specific roles and critically shape cortical function. Yet, how inhibitory circuit specificity is achieved during development is largely unknown. We revealed the transcriptional dynamics of different cortical interneurons during brain wiring and identified subtype-enriched synaptic molecules. Moreover, we showed that the functional connectivity of different interneurons relies on the cell-specific expression of such synaptic genes. Altogether, our results demonstrate that highly selective molecular programs emerging during development in cortical interneurons support their early wiring and underlie inhibitory circuit specificity. After their integration into canonical circuits, activity-dependent plasticity endows neurons with the flexibility required for adapting to sensory experience. Parvalbumin (PV+) interneurons have been shown to play a critical role in this process but the molecular mechanisms by which experience influences PV+ interneuron plasticity were poorly understood. We revealed how perineuronal net (PNN) proteins drive PV+ cell wiring as well as network adaptation to experience. We showed that the PNN protein Brevican simultaneously regulates the excitatory inputs and firing properties of PV+ interneurons by controlling the localization of AMPA receptors and potassium channels, respectively. We also showed that, by modulating Brevican levels, experience influences cellular and synaptic forms of plasticity in PV+ cells and this is required for normal cognitive function. These findings uncover a cell-specific molecular program through which a PNN protein dynamically gates PV+ interneuron function both during development and upon experience-dependent plasticity.
Papers by Emilia so far (* co-first authors)
Favuzzi E*, Deogracias R*, Marques-Smith A, Maeso P, Exposito-Alonso D, Balia M, Jezequel J, Kroon T, Hinojosa AJ, Rico B. Highly selective cell-type specific programs regulate structural synapse target specificity (manuscript in preparation)
Favuzzi E, Marques-Smith A, Deogracias R, Winterflood CM, Sánchez-Aguilera A, Mantoan L, Maeso P, Fernandes C, Ewers H, Rico B. Activity-dependent gating of parvalbumin interneuron function by perineuronal net proteins. Neuron (2017)
Marques-Smith A*, Favuzzi E* & Rico B. Shaping Early Networks To Rule Mature Circuits: Little MiRs Go A Long Way. Neuron (preview), (2016)
Annibali D*, Whitfield JR*, Favuzzi E, Jauset T, Serrano E, Cuartas I, Redondo-Campos S, et al. Myc inhibition is effective against glioma and reveals a role for Myc in proficient mitosis. Nature Communications (2014)
Savino M, Annibali D, Carucci N, Favuzzi E, Cole MD, Evan GI, Soucek L, Nasi S. The Action Mechanism of the Myc Inhibitor Termed Omomyc May Give Clues on How to Target Myc for Cancer Therapy. PLoS One (2011)
 (No Ratings Yet)
(No Ratings Yet)

 (3 votes)
(3 votes)










 Emilia started her studies in 2007 at the Sapienza University of Rome and was awarded a B.Sc. in Biological Sciences with highest marks in 2010. She stayed at the same university for her Master’s project which she performed in the laboratory of
Emilia started her studies in 2007 at the Sapienza University of Rome and was awarded a B.Sc. in Biological Sciences with highest marks in 2010. She stayed at the same university for her Master’s project which she performed in the laboratory of