We invite applications to this permanent full time appointment in the School of Biological Sciences. The successful candidate will be expected to make a significant contribution to biological research in the School and College, and to develop a research programme in their specialist area that will attract external funding and international recognition. They will also contribute to the teaching of biology and related subjects at undergraduate and postgraduate levels.
The successful candidate will be educated to PhD standard or equivalent and have previous experience across a broad range of sub-disciplines within Biological Sciences. To match our vision for the development of biology in the school we would particularly welcome candidates to carry out research to complement our existing areas of expertise (Animal Physiology; Behaviour and Conservation; Wetlands, Biogeochemistry and Plant Science; Microbiology, Parasitology and Biotechnology). From a teaching perspective, applicants should be prepared to contribute to areas that could include: physiology, immunology, biochemistry, human/primate biology, biotechnology, systems biology, quantitative biology, bioveterinary science or forensic biology.
In addition, the post holder will be expected to make a strong contribution to our existing ethos of inter-disciplinarity and team work in research and teaching, enhancing and complementing our existing area of expertise.
The appointment will be made in the range of Lecturer 1 £31,656 – £37,768 (Grade 7) or Lecturer 2 £38,896 – £46,414 (Grade 8) per annum, depending on previous experience.
Informal enquiries can be made by contacting Prof Chris Freeman (tel: +44 (0) 1248 382353, e-mail: c.freeman@bangor.ac.uk
Closing date for applications: 9th September 2016. Interviews will be scheduled shortly after 21st September 2016.
Today’s paper is from the latest issue of Development and investigates the mechanisms of lizard tail regeneration, revealing distinct cell behaviours in the proximal versus distal regenerating tail. Today’s person is Thomas Lozito, Research Assistant Professor at the Center for Cellular & Molecular Engineering, University of Pittsburgh, and co-author with Rocky Tuan.
So Thomas, tell us a little bit about yourself: how did you come to work in the Center for Cellular & Molecular Engineering (CCME) in Pittsburgh?
I actually came to the CCME with my mentor, Dr. Rocky Tuan, when he moved from the National Institutes of Health (NIH) to the University of Pittsburgh in 2009. I attended the graduate partnership program between the National Institutes of Health and the University of Cambridge. I spent the first part of graduate school at Cambridge, and finished up in Rocky’s lab at the NIH. As I was writing up, Rocky accepted a position as the director of the CCME at the University of Pittsburgh, and asked if I would like to come with him as a Post Doc. So I moved to Pittsburgh, first as a Post Doc, and then as a Research Assistant Professor.
What is the general focus of your work within the CCME?
My work focuses on lizard tail regeneration. I try to understand how lizards regenerate their tails, how the regenerated lizard tail tissues are patterned, as well as the cellular origins of regenerated tail tissues. A large part of my work also involves comparing lizard tails with the tails of other regenerative species, such as salamanders, as well as tails of non-regenerative animals, such as mice, in the hopes of finding that special mix of factors that allow for regeneration.
“Turns out no one knew why lizards regenerate cartilage instead of bone, so that’s where I started.”
How was this particular project conceived?
I’ve always been interested in lizards. In fact, herpetology has been a hobby of mine since I was 4 years old. I started with box turtles, and I maintained a small colony in my back yard. Over the years, I’ve kept and bred many different kinds of reptiles and amphibians, including poison dart frogs, monitor lizards, and geckos. As I was approaching the end of my Post Doc, I started to think about what type of research direction I would like to build a career on. Coming from Rocky’s lab and studying cartilage biology, I immediately thought of lizard tail regeneration because the regenerated lizard tail skeleton is almost completely cartilaginous, not bone like the original tail skeleton. Turns out no one knew why lizards regenerate cartilage instead of bone, so that’s where I started.
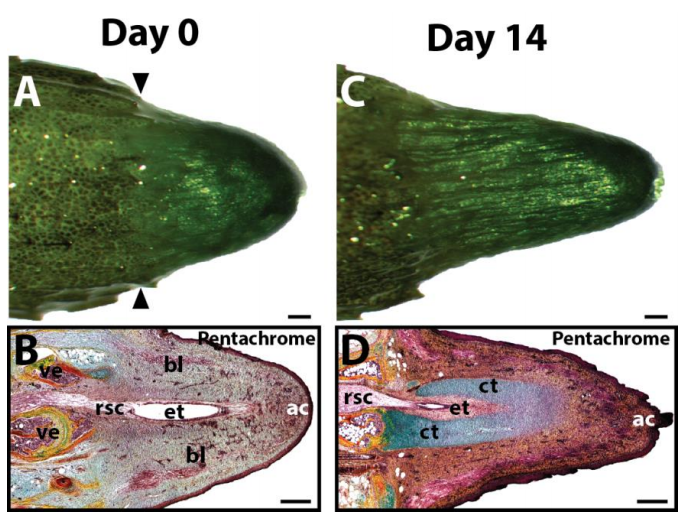
Isolated lizard tail blastemas, at day 0 and 14 after explant culture. From Fig. S1 in the paper.
People will be familiar with salamanders as models for regeneration, but perhaps not so familiar with their lizard cousins. Why use lizards?
Lizards represent an interesting, yet underused, model organism with potential applications to regenerative medicine. As amniotes, lizards are more closely related to mammals, and yet retain impressive regenerative capabilities. Thus, studies involving lizards may be more relevant to improving human regeneration than work done with urodeles (newts and salamanders), the model organisms traditionally used to study vertebrate regeneration. Indeed, several aspects of the urodelian (salamander) life cycle, particularly those pertaining to the larval stage of metamorphosis, are at odds with those of higher vertebrates. In fact, the urodele species most commonly used in regeneration studies, the axolotl salamander, exhibits neoteny, typically never metamorphosing from its larval form. The absence of a larval form in mammals makes meaningful comparisons with salamanders difficult. Lizards, on the other hand, follow a similar developmental plan as mammals and are able to regenerate their tails as adults, making lizard regeneration particularly attractive from a biological and developmental standpoint.
“Lizards are very alert and surprisingly social. They constantly display to one another for territories and dominance, and they definitely watch you as you work and during maintenance”
And what are lizards like to work with as a model organism?
As a hobbyist, working with lizards is second nature to me, and they are very interesting animals. Most only eat live insect prey, so keeping lizards means also keeping feeder insects like crickets. And the lizards are very alert and surprisingly social. They constantly display to one another for territories and dominance, and they definitely watch you as you work and during maintenance. On the science side, working with lizards comes with challenges, since many primers and antibodies aren’t validated for lizards or reptiles. But the challenges make for an exciting research project, I think.
Could you sum up the key results of the paper in a few sentences?
In investigating regenerated lizard tail cartilage, we noticed that the extreme proximal cartilage in contact with the original tail skeleton ossifies, while the rest of the regenerated cartilaginous skeleton does not. In this paper, we described that these differences in cartilage development between the two areas are due to differences in cell sources and signalling. Proximal cartilage forms from periosteal cells in response to signals from the original tail bone, similar to a cartilage callus during fracture repair, while the distal cartilage forms from blastemal cells in response to signals from the regenerated spinal cord.
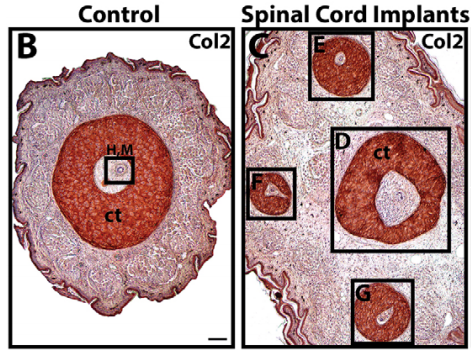
Implanting exogenous spinal cords leads to the formation of multiple cartilage tubes, from Fig. 1 in the paper.
When doing this research, was there a particularly exciting result or eureka moment that stuck with you?
We did some experiments with lizard spinal cord implants. Spinal cords pieces were subcutaneously implanted along the tails of lizards. Wherever a spinal cord piece was implanted, an ectopic tail grew. That was fascinating, and really showed that, in terms of lizard tail regeneration, it’s the spinal cord that provides the regeneration spark.
And what about the flipside: any particular moments of frustration?
Isolating primary cells from lizards, which harbour a lot of bacteria and fungi on their scales, always runs of risk of contamination. We try to be as careful as possible, and take the necessary precautions, but there was a period of several months that every lizard cell isolation got contaminated. That was very frustrating.
“I hope that lizards can teach us new ways to improve mammalian regeneration.”
What can lizards teach us about regeneration in general?
I hope that lizards can teach us new ways to improve mammalian regeneration. As I mentioned, in sparking regeneration in lizards, it basically comes down to the spinal cord, which provides the regeneration spark, so to speak. Mammals like mice don’t have spinal cords in their tails as adults, and can’t regenerate their tails. Is getting a mouse to regenerate its tail as simple as making a mouse with a spinal cord in its adult tail? Probably not, but we can start looking to lizards to see what else they have that is lacking in mammals and that is involved in regeneration.
Is there a particular loose end or unexpected result that you’d like to get to the bottom of?
Of course. In this paper, we’ve determined why proximal regenerated lizard tail cartilage ossifies. But we still don’t know why distal regenerated cartilage does not ossify. Yes, distal cartilage is derived from a different cell source than proximal cartilage. But I’d like to know if distal cartilage cells are even capable of ossification. So far our results say no, but I’d like to know why not.
And what are you working on now?
I just got an R01 grant from NIGMS to answer the question “Why don’t lizards regenerate perfect tails like salamanders?”. Regenerated salamander tails are near-perfect copies of their tails, complete with dorsoventral patterning and segmentation. “Imperfect” regenerated lizard tails, on the other hand, lack dorsoventral patterning and do not segment. As part of the grant, I’m trying to identify the molecular and cellular reasons behind these differences, and ultimately use genome-editing technologies like CRIPR/Cas9 to correct some of the “imperfections” in regenerated lizard tails by introducing patterning and segmentation.
What do you like to do when you’re not surrounded by lizards?
Since my hobby is still herpetology, a good portion of my spare time is also spent surrounded by lizards and frogs! I currently keep several types of geckos and frogs as pets. But I am basically interested in all animals. Wherever I travel, I try to visit the local zoo, and most of my vacations are of the “ecotourism” sort where I try to observe some exotic animals in their natural habitats. For example, I went to Costa Rica and Panama a couple years ago to see some of their endangered frogs. And this year I visited Crystal River in Florida to see and swim with the manatees. Closer to home, I enjoy movies, especially super hero movies, and spending time with friends and family, especially when it involves good food.
The joint meeting of the Society for Developmental Biology and the International Society for Differentiation was held in Boston between August 4th and 8th. It was a meeting of firsts for me: first meeting representing the Node, first time at the SDB (having regularly enjoyed the Brit version, I was looking forward to hearing the US angle on what’s exciting at the moment), and first time in Boston. The venue was the gargantuan Copley Place Marriott, which offered a stunning view of the city (from my hotel room at least) as well as a central base to explore. Boston turned out to be beautiful and walkable. Great local beer too.
Boston, looking south from the Marriott
True to our diverse field, the meeting covered a lot of bases, from cell mechanics to genome editing, patterning to computational modelling, as well as life away from the bench (education, publishing, grant-writing) and the planet (designing experiments for NASA’s Genelab). What follows is a selection of my personal highlights from the talks I saw. Let me know if I missed anything or have mangled things beyond comprehension.
Regenerating parts and patterns
I began the meeting in a satellite symposium, Evolution of Regenerative Capacities: recapitulation of development or novel mechanisms? The symposium set the tone for the rest of the meeting in terms of speaker composition: a mix of postdocs, young PIs and established leaders in the field, plus the odd grad student, and an even gender balance.
Most of the speakers started with the same observation: some animals are great at regeneration, some (like us) not so great, and even within the same animals, some tissues regenerate better than others (though axolotls apparently can regenerate everything!). However, we really lack a mechanistic explanation for this spectrum of regenerative capacity; such an explanation might inform our efforts to coax currently intransigent human tissues to regenerate. The evolution part came from the range of species under investigation, which allowed for a fairly broad phylogenetic sample.
For someone fairly naïve to this field, I was struck by the diversity of mechanisms used in the different contexts, as illustrated by a rough list the proteins discussed: piwi piRNA pathway members, various transcription factors, second messengers, growth factors, cytokines, cell adhesion molecules, glycosylation enzymes, and matrix proteins. (I guess this shouldn’t really have been surprising given the tasks at hand for the wounded tissues.) The extent to which regeneration redeploys developmental pathways also seems to depend on the tissue: for instance, Ken Muneoka described how digit regeneration in the mouse shared some features with normal bone development, but not others.
One of the more remarkable stories involved the moon jellyfish, described by Lea Goentoro. Ephyra (juvenile medusa) stage animals typically have 8 evenly-spaced arms which pulsate synchronously to drive propulsion and feeding. Cut off some of the arms, and the ephyra proceeds not to regenerate them, but to reorganise its remaining limbs to regain radial symmetry, in a manner requiring muscle contractility but not cell division. The ephyra regenerates not body parts but a feature of the body pattern, perhaps because repaired animals have still got a reasonable chance of surviving. Got to love nature sometimes.
Gene expression and chromosome topology
After the regeneration fest, KenZaret kicked off the Presidential Symposium by celebrating developmental biology, and insisting on the continued need for basic research in model systems (his reaction, perhaps, to recent events inthe US and Canada). He then stated that, for him, changes in gene expression are the real heart of development (the symposium did have an epigenetics theme, after all). Ali Shilatifard, in his exploration of histone methylation and childhood leukaemia, agreed. Coming from a bit more of a cell mechanics background, I wanted to come up with a counter argument, but by that point didn’t have the brain power for it.
One of the most remarkable epigenetic feats is the inactivation of an entire X chromosome in female mammals, providing almost complete gene silencing from early development onwards. The long non-coding RNA Xist is key to inactivation, and Jeannie Lee described how RNA proteomics had identified the proteins that interact with Xist (it turns out there are a surprisingly large number of them), andhow super-resolution imaging had visualised individual Xist molecules decorating the inactive X (it turns out there are a surprisingly small (~100) number per X). An analysis of Xist mutants then implicated other silencing mechanisms acting synergistically with Xist, and showed that even modest changes in X gene dosage can have profound effects on viability.
Revelations about the X chromosome resulting from a combination of cutting-edge technology and genetics…history repeated itself the next morning, in Job Dekker’s talk, part of a fascinating Chromosome Topology and Gene Regulation session. Revealing the organisation of the inactive X is complicated by the fact that the two copies of the chromosome are too similar to be distinguishable by HiC. The solution: cross two mice lines, creating a (frankly rather shabby looking) F1 hybrid, which contains enough SNPs to distinguish maternal and paternal chromosomes, and then derive ES cells from these mice and induce X inactivation in them. Dekker showed howthese cells revealed the fine-grained structure of the inactive X, with its two highly compact mega-domains, mysterious boundary region between them, and islands of topologically associating domains around the few genes that remain active.
After a delightful interview with Dave McClay (look out for it in an upcoming Development issue and on the Node), and an hour jaywalking around Boston for lunch, I was back for the Hilde Mangold Postdoctoral Symposium.
From a very strong field (the eight talks were picked from over a hundred applicants), two really stuck with me. Yaniv Elkouby, from Mary Mullins’ lab at UPenn, documented theintricate relationship between the centrosome, telomeres, a chromosomal ‘bouquet’, microtubules, and the Balbiani Body in the polarisation of the fish oocyte. Beautiful in vivo cell biology. Jacqueline Tabler (John Wallingford lab at UTexas), pointed out that given how critical vocalisation is for vertebrates, it’s striking how little we know about the development of the organ that creates vocalisations, the larynx. Aided by her own whistling and beautiful images, she told us how interfering with Hedgehog signalling led to an expansion of neural crest-derived tissues in the larynx, and defective vocalisations in newborn pups. (Her previous work has looked at various aspects of craniofacial development.)
Friday afternoon brought another plenary: Biology at the level of single cells. Single cell sequencing is a remarkable advance, but sequencing for the sake of sequencing does not necessarily advance the field; you need a strong biological question. It was therefore wonderful to hear Mark Krasnow reconstruct the program of alveolar development by sequencing individual cells of various stages from a dissociated lung. With exquisite sensitivity (‘a single RNA in a single cell’), the technique revealed new players in the developmental program by correlating expression with differentiation markers, providing a quick, non-genetic screen.
Tony Hyman then took us back to basics and asked: how does the cell organise chemical reactions without membranes? Part of the answer appears to beliquid-liquid phase separations. Unstructured protein regions (for instance in prion-like domains) promote phase transitions, and changing the sequence of these regions could alter the properties of the resulting agglomeration, from liquids that have fast turnover and promote chemical reactions, to gel-like, stable, dense matrices prohibitive of reactions (such as the Balbiani body we encountered in Yaniv’s talk earlier in the day). The talk really made me think differently about the way cells work.
The Charles River
Fat fish and organoids
I started Saturday in the Emerging model organisms and evo-devo session. The general feeling was that techniques like CRISPR were opening up many organisms to developmental analysis; your question might now dictate your model, rather than the other way round. (Lo and behold:a recent piece in Nature.)
Nicolas Rohner introduced us to Astyanax mexicanus, the blind cave fish, which has emerged as a model for metabolic diseases. These fish are always hungry, get really fat (without any clear health disadvantages), and are remarkably resistant to starvation. They are also an evolutionary windfall: not only do river forms persist (providing an ‘ancestral’ population that blind forms can still breed with), but the cave forms themselves have evolved multiple times independently, so they are a great model for convergent evolution.
During the coffee break, I interviewed Doug Melton – another wonderful half hour, look out for the write-up in the future – prior to his Jean Brachet lecture, in which he promoted the idea of applied developmental biology in harnessing stem cells to treat diabetes. Applied developmental biology is also being advanced by organoids, which were celebrated in a special plenary session in honour of Yoshiki Sasai. One recurrent problem with organoids is the variation you get between samples, and Jürgen Knoblich described how growing brain organoids on strands of a synthetic polymer reduces variability, and promotes development of a cortical plate-like structure . Hans Clevers discussed modelling cancer with tumour organoids (somewhat counterintuitively, tumour-derived organoids always grow slowly), and promoted the idea that, in many cases, stem cells are not a rigid cell fate, but a labile cell state.
Downtown Boston, looking like a toy town
Circumventing Mendel and celebrating scientists
Sunday: the final day; bleary-eyed and near-saturated, but not dead yet. Before the SDB award lectures, I heard from Ethan Bier on the ‘mutagenic chain reaction’, a CRISPR/Cas9 technique which gives a rapid form of non-Mendelian inheritance. Not only does this drastically cut the time it takes to make mutants (hypothetically, to make a mouse quadruple mutant: 1/64 mice in the 4th generation of a standard cross; 64/64 in the 2nd generation using MCR), and provide a tool for organisms without established genetic techniques, it also allows you to rapidly spread a genetic element through a population (a gene drive). Stunning, revolutionary genetics.
The afternoon Education Symposium was a change of gears, as educators discussed how to provide biological training in institutions with shrinking budgets. The general consensus was that inquiry-driven learning could provide a more engaging route for biology students than your standard rote lab work (this report sums up a lot of the speakers’ ideas). Getting undergrads to feel a sense of ownership over the projects also helps: Sarah Elgin described how hundreds of undergrads had contributed to the annotation of the oft-forgotten dot chromosome of Drosophila melanogaster (leading to a paper with over a thousand authors!).
And finally, the SDB awards. Ida Chow (Viktor Hamburger Outstanding Educator Prize) encouraged all of us to fight for science and science education, and got a raucous standing ovation (from a show of hands, many in the audience had directly benefited from one of her many initiatives). We got to see old pictures of Dave McClay (Lifetime Achievement Award) as a high school football player and an underwater diver, before he described how each of the EMT events in the early sea urchin is driven by a separate gene regulatory network. And finally, we heard from Kathryn Anderson (Edwin G. Conklin medal; I also had the pleasure of interviewing Kathryn, keep an eye out for it) who described the beauty and mystery of gastrulation in the mouse.
The evening ended with drinks, poster awards, dinner (including waiters impatient to get you on to your next course: “coffee with your chicken, Sir?”), drinks, dancing, watching-of-the-dancing, and then final drinks in a dive bar (Cambridge really lacks places like these).
Pre-dinner drinks in the Marriott
I left the meeting with the feeling that developmental biology is in a good state. This was not just down to the general positivity of Americans: new technologies are unlocking doors everywhere, and the increasing applicability of developmental research to human health (with stem cells, organoids) gives a public persona that I hope will draw in more money without side-lining all of the strong non-applied research that sits under the DB umbrella. Coming from England, you also forget how vast the US is and how much wonderful research is being carried out there.
I would also like to commend all of the speakers who suffered technical difficulties and kept their poise to finish their talks, rather than melt down and destroy the equipment (in one of the meeting’s funniest moments, Mina Bissell walked the length of the hall to fix her frozen computer at the back AV desk, all the while giving running commentary).
And finally: the posters. More than 500, each up for one night only, they were excellent too, and included many with added extras (3D printed models, interactive whiteboard sections, and built-in videos!).
Quotes of the meeting (paraphrased, potentially misremembered)
“Three cheers for developmental biology!”
Ken Zaret opening the meeting, cue cheers and applause.
“This is for you, Sarah Palin…”
Ali Shilatifard on how research in flies helped to inform children’s cancer treatment
“Like Republicans, Aedes aegypti ‘repeal and replace’”
Marc Halfon on mosquito GRN evolution (not that I immediately got the reference)
“It is not birth, marriage, or death, or gastrulation, but epithelium formation that is truly the most important time in your life”
Matthew Gibson takes Wolpert back a step
“If your grandfather was a fish, he’d have no problem hearing you”
Shawn Burger on the amazing regenerating ear hair cells of zebrafish
“I justlove the spiny mouse!”
Overheard at the Regeneration symposium (before I found out that they were a species, not a Pokemon)
“Blind cavefish are fat”
Nicolas Rohner pulls no punches in describing his beloved model organism
“We must build the embryo to understand it”
Eric Siggia goes a bit Feynman
“If you’re not arrogant, you can look at your data and see what it’s trying to tell you”
Mina Bissel warning against over-confidence
“There are no ‘higher’ or ‘lower’ organisms”
Richard Berhinger on Twitter
“Other hashtag is … different”
James Gagnon on Twitter, after confusion over #2016SDB (the correct one) versus #SDB2016 (Seventh Day Baptists)
We are a major international research centre at the forefront of studies in mouse genetics and functional genomics. We are investigating a wide variety of disease models in mice, in order to enhance our understanding of the molecular and genetic basis of disease.
About the Role
We require a leader for the newly-formed Gene Delivery group. Central to the generation of new mouse models is the technical process of delivering genetic material into mouse germ cells (usually embryos). You will manage the delivery of this service and establish/develop new techniques of embryo manipulation in order to exploit the potential of technologies such as genome editing.
We provide a dynamic and progressive atmosphere for science-led service delivery, supporting the generation and characterisation of mouse lines for the study of human diseases. With a leading role in the International Mouse Phenotyping Consortium (IMPC) and co-ordinating the UK Genome Editing Mice for Medicine (GEMM) initiative, MRC Harwell is at the forefront of the provision of high-quality mouse lines and phenotyping data to the international genomics community.
About You
It is essential to have previous experience with transgenic techniques and a PhD or Masters in molecular embryology/biology. You will have experience leading a team effectively. You will also have experience in Vivo experimental work, including statistical analysis of the experimental data.
Benefits
The MRC is a unique working environment where our researchers are rewarded by world class innovation and collaboration opportunities that the MRC name brings. Choosing to come to work at the MRC means that you will have access to a whole host of benefits from a final salary pension scheme and excellent holiday entitlement to access to employee shopping/travel discounts and salary sacrifice cycle to work scheme and childcare vouchers, as well as the chance to put the MRC on your CV in the future. We are strongly committed to your personal development and well-being, with extensive opportunities for training and flexible working.
Heart development is a complex process involving numerous cell types and different morphogenetic events to make an organ that begins its vital function long before it reaches its final, intricate shape. That this process is error prone is borne out by the high prevalence of congenital heart defects in humans. For de la Cruz, a Cuban cardiac embryologist who spent most of her career in Mexico City, a precondition for understanding the aetiology of congenital defects was a comprehensive description of normal heart development. She was particularly preoccupied with the question of where the various parts of the final (or ‘definitive’) heart mapped onto earlier developmental stages.
Her 1989 paper begins with a nod to an earlier age: the descriptive work of Carl Davis, who in the 1920s used human embryo samples from the Carnegie Collection to infer the lineage of the compartments of the heart. Davis had described one particular embryo, #3709, the type specimen for stage 9, at which point the heart is a straight, symmetrical tube, yet to undergo looping and formation of the chambers (the same embryohas recently been reconstructed in 3D).
Davis inferred that each of the regions of the straight tube heart is the primordium of a definitive cardiac cavity; that is, by stage 9, the atria and ventricles of the heart are mapped out onto the straight tube heart, and subsequent development involves morphogenesis of these pre-patterned regions.
For de la Cruz, decades later, purely descriptive embryology on fixed and sectioned samples was inadequate to really test lineage, as there was no means of following regions in the same heart. She thus turned to in vivo labelling, which allowed the “study of the cardiac zones up to their anatomical expression in the mature heart in a continuous and uninterrupted sequence”. Of course she required a non-human model, and used the easily accessible eggs of leghorn chickens (like mammals, birds have a four-chambered heart). Regions of the heart could be injected with iron oxide particles, and the embryo left to develop either in vitro or in ovo until the desired stage. Over the decade preceding the paper, she and others had gathered evidence which questioned the existence of Davis’ primitive cardiac cavities; for instance, the primordial atria were not apparent in the heart tube tissue, but only later in the loop stage heart.
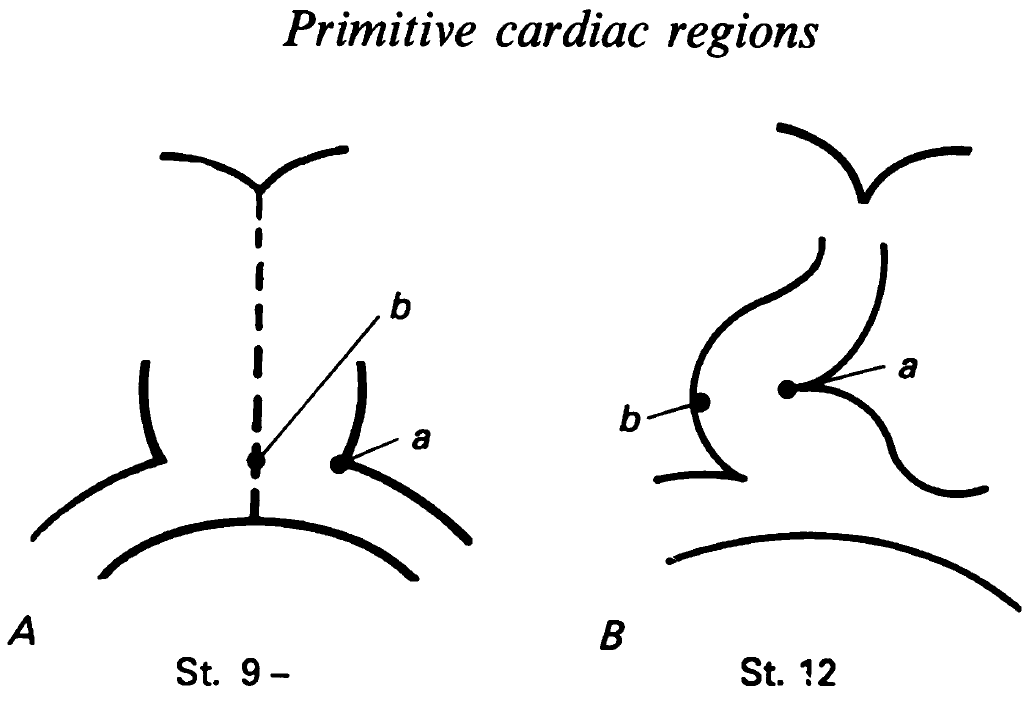
Excerpt from Figure 2 showing labelled regions in St. 9 hearts, and where they ended up in St. 12. de la Cruz, et al. 1989. J Anat. 165:121-131. Reproduced with permission of Wiley.
The 1989 paper was her latest attempt to map out where the regions of the heart tube ended up, and would she hoped “allow us finally to discard the term ‘primitive cardiac cavities’.” To start with, the straight tube heart was labelled in two caudal regions (a and b in the figure excerpt), and then the label was observed in the loop heart stage. Label a, at the left border of the heart tube, ended up in the left border of the loop stage heart, consistent with previous descriptive embryology. But label b showed something different: cells at the tube’s midline ended up at the right border of the loop, which dismisses a simple ‘one to one’ correlation between the regions of the tube and the loop. A final caudal label, at venous edge of the tube, ended up in the border between the left ventricle and left aorta in the mature heart. These labelling experiments were complemented by SEM and histology to compare the morphologies of the straight tube heart with later stages.
This paper, along with de la Cruz’s previous and later work, demonstrated that the straight tube heart does not contain single primordia for each of the definitive cavities. As the initial heart tube only contributes to a subset of the final heart (mainly the left ventricle), the rest of the heart must be added during later development. This work predated the molecular definition of additional regions of the mesoderm that give rise to the heart (the so-called second heart field) by more than a decade, but appears not to have been widely appreciated, perhaps due to the methods employed. It could however be argued that the power of the method is its simplicity: label a cell or set of cells and see where they end up, and deduce from there how the final organ is built. Indeed lineage tracing is still being used to great effect to this day.
Thoughts from the field:
Benoit Bruneau (Gladstone Institute for Cardiovascular Disease)
“Victoria de la Cruz used somewhat crude methods to map out where segments of the early heart tube end up in the more developed heart, and the results were not what people might have expected based on preconceived notions. Her work went largely ignored as people examined gene expression patterns and made erroneous conclusions about the chambers of the heart being already patterned and present as primordia in the linear heart tube. The discovery of the second heart field brought back to the forefront her work, which suggested that there might be an additional source of heart cells.”
“The chick embryo is easily accessible and has been used extensively for fate mapping studies. This paper by Maria Victoria de la Cruz is a prime example of classic mapping experiments that contributed to changing the thinking in the field. Her work using labeling of the linear heart tube with iron oxide particles, indicated that new heart segments are added successively, in particular to generate outflow myocardium. She also concluded that precursors for the right and left primitive atria are not yet present in the early straight heart tube but become incorporated later during loop stages. This was not fully appreciated until the origins of secondarily added cell populations were discovered; in the chick using essentially similar approaches, and in mouse using genetic labelling.”
“This 1989 paper was an important step in a series of cell labeling studies from Maria Victoria de la Cruz and colleagues that demonstrated the dynamic nature of early heart development. Her purely embryological approach was initially underappreciated yet set the scene for many of the molecular and genetic studies underway today. MV de la Cruz was one of the first to realise that there was a myocardium-forming region outside the linear heart tube, a critical advance in emergence of the second heart field model of vertebrate heart development. Indeed, the concepts that certain parts of the heart are late added components and that primitive cardiac regions only contribute to parts of the definitive cardiac chambers stem from this work and have had significant impact on our understanding of congenital heart defects. Her book Living Morphogenesis of the Heart is highly recommended further reading for those interested in the major embryological questions, including heart tube growth, septation, coronary development and the mechanisms underlying anomalous cardiac development, that continue to drive the field today”
A little more reading
I learned a little about Maria de la Cruz’s life from two obituaries (1, 2)
This post is part of a series on forgotten classics of developmental biology. You can read the introduction to the series here and read other posts in this series here.
We are seeking an enthusiastic and outstanding postdoctoral researcher to join a multidisciplinary team led by Prof. Chris Thompson at the University of Manchester. You will use single cell RNA sequencing to identify groups of heterogeneously expressed genes within normal populations of cells, and study the role of these genes in cell fate choice.
Cell fate choice and proportioning are typically considered to be ordered, robust and reproducible. However, noise and stochasticity can lead to heterogeneous gene network activity. Consequently, it has been proposed that gene networks may be ‘wired’ to buffer these fluctuations. Alternatively, heterogeneity may be functionally important to prime cells or increase the spectrum of differentiation capabilities. Addressing these questions represents one of the greatest challenges in Developmental and Stem Cell Biology. However, to date it has been impossible to follow entire gene network behaviour in individual cells, or to follow their temporal changes in activity in individual cells as cells commit to differentiation along different lineages. Single cell gene expression analysis, together with novel computational reconstruction of gene network dynamics provides this opportunity.
This work builds upon our recent finding (Chattwood et al, eLife 2014) that the interplay between dynamic heterogeneity in Ras-GTPase activity and nutritional status is required for normal lineage priming and robust running of an ultradian cell fate oscillator. Computational approaches will be used to identify putative genes involved in lineage priming and cell fate choice. In addition, the role played by these genes will be tested in the lab through the analysis of gene knockout strains, live cell imaging and molecular genetics.
Candidates with extensive experience of using either computational approaches or wet lab approaches to understand the molecular basis and gene networks will be considered.
You should currently hold or be about to obtain a PhD in a relevant field.
The post funded by the Wellcome Trust and is available for up to 3 years.
In 2009, FaceBase was launched in response to the need for more comprehensive analysis of craniofacial development: with so much craniofacial data being generated, there is a danger of relevant datasets being buried in the avalanche of genomic and other data. FaceBase is a curated, one-stop shop for facial development and research offering the community input and access to datasets that can bring their research to the next level. After the first 5 years (known as FaceBase 1) resulting in almost 600 datasets and over 100 publications, the next phase of FaceBase (FaceBase 2) began in August 2014 with a new Hub that developed an updated data model allowing for more data integration and faceted searches with a new server interface. The FaceBase website (http://www.facebase.org/) continues to be a resource for the community.
What is FaceBase? A collaborative NIDCR-funded consortium to generate data in support of advancing research into craniofacial development and malformation.
Serves as a community resource by generating large datasets of a variety of types and making them available to the wider research community at http://www.facebase.org/.
Emphasizes a comprehensive and multidisciplinary approach to understanding the developmental processes that create the face.
Spotlights high-throughput genetic, molecular, biological, imaging and computational techniques.
Facilitates cooperation and collaboration between projects and beyond.
Find datasets by various filters: organism (e.g., human, mouse, zebrafish), investigator, data type, various developmental ages, mutation, genotype, gene, chromosome and much more.
New data are now available from the FaceBase 2 projects as well as continued uploads of FaceBase 1 datasets
What you’ll find:
Global and specific gene expression patterns
Regulatory elements and sequencing
Anatomical and molecular atlases
Human normative facial data and other phenotypes
Genetics of craniofacial development
Repositories of animal models and of human samples and data
Software tools and animal models for analyzing and testing and integrating these data
Resources for researchers who may be new to the craniofacial field
Finding the data you need:
Start your search by clicking the Data Browser button on the homepage or in the top navigation. The video below provides a tutorial on the site’s search functions. There is also extensive documentation on the website.
3D skull anatomy:
Here is a video which highlights every bone in the craniofacial region and provides internal and external views of the skull as well as measurements of selected bones.
FaceBase 1 projects (2009-2014) and their contributions:
Contributed MR Image data of normal and a mouse model of cleft palate (tgfß KO) presented as orthogonal sections through fetal embryo heads (stages 13.5-18.5).
Mapped on a genome-wide scale distant-acting gene regulatory sequences (enhancers) involved in craniofacial development and examined the precise activity pattern of individual enhancers in transgenic mouse assays.
Used laser capture micro dissection to isolate specific compartments of the developing face, which were then used for gene expression profiling with microarrays and/or RNA-seq.
Produced software – CranioGUI – that can be used by the craniofacial research community to help analyze facial abnormalities. The software works on 3D head meshes and can be used for multiple conditions.
FaceBase 2 projects (2014-present) and data currently being deposited:
Advance the use of zebrafish in the study of skull development, and facilitate comparative studies with mammals that will advance treatment options in human patients.
Characterize epigenetic landscapes and transcriptomes of human and chimpanzee cranial neural crest cells and analyze candidate human-specific craniofacial enhancer activity in vivo.
Develop and maintain an infrastructure that will store, represent, and serve craniofacial data to the research community and develop tools for visualizing, integrating, annotating, linking and analyzing the data
Characterize the gene regulatory landscape of craniofacial development using epigenomic profiling of developmental mouse and human facial tissues, coupled to characterization of craniofacial enhancers in transgenic mice.
Develop a genomics analysis interface that makes analysis of pertinent genomics data available to FaceBase users without releasing the individual level data.
Generate comprehensive gene expression atlases of the major and functionally important craniofacial sutures of the mouse.
References:
The FaceBase Consortium: a comprehensive resource for craniofacial researchers.Brinkley JF, Fisher S, Harris MP, Holmes G, Hooper JE, Jabs EW, Jones KL, Kesselman C, Klein OD, Maas RL, Marazita ML, Selleri L, Spritz RA, van Bakel H, Visel A, Williams TJ, Wysocka J, FaceBase Consortium, Chai Y. Development 2016 143:2677-2688. doi: 10.1242/dev.135434
The FaceBase Consortium: a comprehensive program to facilitate craniofacial research. Hochheiser H, Aronow BJ, Artinger K, Beaty TH, Brinkley JF, Chai Y, Clouthier D, Cunningham ML, Dixon M, Donahue LR, Fraser SE, Hallgrimsson B, Iwata J, Klein O, Marazita ML, Murray JC, Murray S, de Villena FP, Postlethwait J, Potter S, Shapiro L, Spritz R, Visel A, Weinberg SM, Trainor PA. Dev Biol. 2011 355(2):175-82. doi: 10.1016/j.ydbio.2011.02.033
In this new series, we interview the people behind some of the most exciting recent papers in developmental biology and related fields, to give context to the work and find out how the story came together.
To inaugurate the series, we start with a paper that came out recently in Cell, and uncovered a mechanism for how nuclear pore complexes are inserted into the nuclear envelope in early Drosophila development.
We hear from five of the people behind the paper, all of whom are based at the EMBL in Heidelberg: lead author Bernhard Hampoelz, his PI Martin Beck, and their EM collaborators Yannick Schwab (PI and head of the EM facility), Nicole Schrieber and Paolo Ronchi.
They gave us their perspectives on this collaborative, multi-disciplinary project.
Introducing the players, from left to right: Nicole, Bernhard, Yannick, Paulo and Martin
We’ll start with Martin: can you tell us the brief history of the Beck lab, and what key questions the group is trying to answer?
MB I have a mixed training in structural biology (PhD with Wolfgang Baumeister at the MPI Martinsried) and systems biology (postdoc with Ruedi Aebersold at ETH Zurich). I had already worked on nuclear pores as a student and when I started my own laboratory at EMBL in 2010, I felt that my training positioned me well to attempt to understand nuclear pore complex architecture. We have combined electron microscopic with mass spectrometric approaches to structurally analyse nuclear pores in situ.
I was always intrigued by two biological aspects of this: i) that understanding the assembly pathways of large macromolecular machines can help us to understand their architecture because nature essentially has broken down the problem into smaller pieces for us that are more feasible to approach. And ii), that in order to understand the function of large macromolecular machines in situ, one needs to understand how their structure is spatiotemporally modulated, e.g. across cell types or the cell cycle.
From my perspective, the beauty of Bernhard’s project is that it brings all of this together to elucidate a new phenomenon. It builds on the methodological strength and quantifies a compositional variation of nuclear pores across space and time to discover an unanticipated way to get a nuclear pore complex (NPC) in to the nuclear envelope.
And Bernhard, how did you come to join the Beck lab? Am I right in thinking you brought flies to the lab?
BH Exactly – I think flies were not very popular in Martin’s group before I entered. Also my background was perhaps unusual for the lab, since I came from developmental biology. When working at the IBDM with Thomas Lecuit, my focus was on nuclear morphology and its developmental control. Thomas gave me a lot of freedom in investigating things that popped up and it happened that I observed annulate lamellae (AL; stacked cytoplasmic membranes that are a subset of the ER and decorated with NPCs) insertion into the nuclear envelope by imaging. Aware of the potential of this finding, I knew that in order to nail this down mechanistically I had to convince somebody that works on NPCs at the ultrastructural level to give me the chance to pursue this project. I came to EMBL as a visiting scientist and soon presented my findings to Martin. I am very glad that Martin, although the project was not in the direct focus of his lab at that time, agreed on supporting me to continue this work in his group.
“I knew that in order to nail this down mechanistically I had to convince somebody that works on NPCs at the ultrastructural level to give me the chance to pursue this project.”
So Yannick, you’re neighbours in Heidelberg, but did you know Martin well beforehand? How did you get involved in Bernhard’s project?
YS Martin and I work on the same floor at EMBL. In fact, I head the electron microscopy core facility (EMCF) which is sharing the space where Martin’s Unit (Structural and Computational Biology) has its set of cryo electron microscopes. Therefore, we see each other very often. Even though Martin’s group is focused on structural biology, they do not hesitate to cross the border towards cell biology which is the field of expertise of the facility. Whilst Bernhard had already solid background in EM and was already an advanced user, his project required advanced expertise both in volume EM and in Correlative Light Electron Microscopy (CLEM). For this, he teamed up with some of the EMCF staff (Pedro Machado, Paolo Ronchi and Rachel Mellwig) and with Nicole Schieber a specialist in Focussed Ion Beam Scanning Electron Microscopy (FIB-SEM) from my team. My involvement in this project was mostly at the level of organizing this collaborative work and setting priorities when the last set of experiment had to be done.
Finally, Nicole and Paulo: how were you recruited to this story?
NSI started with the Schwab team when it was just beginning as the research technician. My background has had a strong focus on Electron Microscopy for the past 9 years since my undergraduate degree got me hooked at the University of Queensland, Australia. For the past 3 years here at the EMBL I have shifted more towards the 3D EM techniques, especially FIB-SEM.
My role in the lab means that I try to connect the team to the facility and on some occasions I can step in to help with projects that I either find interesting or see I can add some expertise. For this story, my colleagues Rachel and Pedro from the EMCF were already working on this project together with Bernhard and I had been following its progress in our regular meetings. They had managed to solve some difficulties with the EM sample preparation and we all quickly realised that FIB-SEM would add to the three dimensional picture of the story. Since this would be a demanding task for the FIB-SEM I put my hand up to acquire the data and really enjoyed the challenge as well as being able to team up and work more closely with my colleagues than we normally would.
PRI have been working in the EMCF for 2 years after a postdoc experience at EMBL, working on membrane trafficking. Since I’ve been here, I have always been very keen on pushing CLEM methods in the facility and Bernhard’s project was a great opportunity. But it was a big challenge as well.
I got an email from Yannick asking whether I would be interested in helping Bernhard with a CLEM experiment. I knew his project and I had always found it very interesting, but it was on short notice (it was for the revision of the paper) and I was on holiday! The project required to adapt a high accuracy CLEM method that had been previously developed on yeast cells to the Drosophila embryos. Luckily, I had been setting up the best conditions to treat Drosophila ovaries for another collaboration and thought that Bernhard’s system was similar enough. Therefore I used the same protocol and, for once, everything worked smoothly at the first attempt.
Where did the interest in nuclear pores come from, Bernhard? And what was the key problem you wanted to address with this paper?
BH I have to admit that my interest in NPCs came by accident. In Thomas Lecuit’s group I worked on nuclear morphology and used fluorescently labelled Nucleoporins as means to outline the nucleus in imaging. Naively I realized that these Nups do not only label the nuclear envelope (NE) but also foci in the cytoplasm and I learned about AL, which have been known for decades actually. Curious about their function, I imaged them live and saw that they insert into the NE. Puzzled by the fact that this has not been observed before I started to think how to further develop this project.
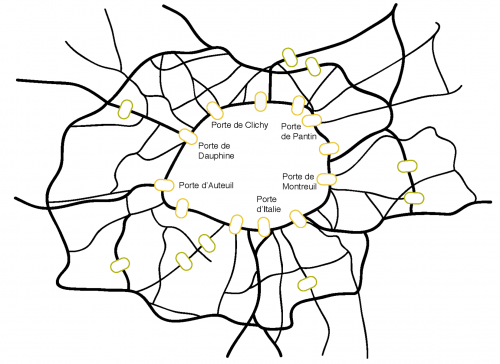
A dense network of roads feeds into the Boulevard Périphérique, which surrounds the city of Paris. Major junctions, some of which are depicted in this image, bridge the Périphérique and link the city to the metropolitan region. Likewise Nuclear Pore Complexes (NPCs) pierce the nuclear envelope (NE) and control transport between the cell nucleus and the cytoplasm. The NE is continuous with the surrounding ER network that feeds nuclear expansion during interphase. Credit: Bernhard Hamploez.
What makes the early Drosophila embryo an ideal model for the question of nuclear pore insertion? What are the model’s challenges?
BH The fly embryo offers a couple of advantages. The most intriguing is probably its ease to do live imaging. Basically you just glue an embryo expressing your fluorescently labelled protein of interest onto a coverslip and start to image. Moreover the embryo is susceptible to injection of drugs or genetic manipulation that could interfere with your process of interest. Conceptually, I like that it allows you to put a cell biological question into the context of animal development. A challenge is the short cell cycles, especially for EM – we sectioned numerous embryos that happened to be frozen during mitosis and were thus useless for our purpose.
“Basically you just glue an embryo expressing your fluorescently labelled protein of interest onto a coverslip and start to image.”
Can you briefly describe what you found out about the relationship between annulate lamellae, nuclear pore complexes, and the nuclear envelope?
BH AL are sub-compartments of the endoplasmic reticulum that contain stockpiled NPCs. Because AL are in particular enriched in oocytes and early embryos across species, they were always considered as storage pools of maternally derived NPC material. Whether they could somehow contribute to the NPC pool at the nuclear envelope had remained elusive. Our study proves that AL are indeed inserting into the nuclear envelope when the nucleus expands during interphase in Drosophila embryos at the blastoderm stage. This can work because unlike in differentiated cells NPCs in the early fly embryo are laterally mobile within the NE and thus can redistribute. We reveal that NPCs at AL are pore scaffolds that only mature to the full NPC complement once inserted into the nuclear envelope. And, based on EM, we suggest a topological model how such insertion could happen.
So what makes a nuclear pore at the AL different from one in the nuclear envelope?
MB Its composition. Certain nucleoporin subcomplexes are missing in AL. This includes for example the Nup214 complex that is important for mRNA export out of the nucleus in differentiated cells but also for some import pathways that might play already a role in the early embryo. We believe that it is assembled into AL-originated pores only after they inserted into the nuclear envelope.
And why was EM necessary? I understand three different techniques were used (CLEM, FIB-SEM and tomography): were any of them a particular challenge?
YS EM in general is a challenging set of techniques. From the sample preparation to the image analysis, specific skills and dedicated protocols are required. Fortunately, the EMCF has accumulated a great deal of experience since its creation more than 10 years ago. Benefiting from a good integration within the research scene, it has, among others, developed specific methods for preparing Drosophila embryos (by high pressure freezing and freeze substitution) and for imaging them with 3D EM (tomography and FIBSEM).
NS FIB-SEM is a difficult technique that requires a lot of patience but is extremely rewarding in the results you can obtain. For this project one of the main difficulties came after finding the correct stage of the embryo and an interesting event by TEM thin sections. We then wanted to immediately image this same embryo in the FIB-SEM by looking at the block face itself and use the focussed ion beam (FIB) to ablate very thin slices to allow consecutive images that build the 3D data. During standard preparations for FIB-SEM you have a buffer where you can allow the sample to stabilise during the imaging so that the slice thickness is consistent, and can also test the imaging conditions to get the best possible resolution. In this case we didn’t have such luxuries and this was a challenge especially since the event we were looking for required the best possible resolution. The other challenge came from remounting the sample from how it was sectioned for TEM to the stub for FIB-SEM, here we were dealing with a piece of resin containing the sample that was very small (in the range of several hundred microns) and the orientation of this block was critical.
PR The CLEM method we used was developed by the Briggs and Kaksonen groups at EMBL to study endocytosis on yeast and mammalian cell cultures. Their work has made this technique very popular in the community and therefore our facility in the last 2 years has dealt with a number of projects using this method, adapting it to different organisms and different subcellular structures. Bernhard’s case was more challenging because we had to look for embryos of the right developmental stage and in the right stage of the cell cycle (interphase). After preparing the samples for EM, we therefore sectioned a few embryos and inspect them by conventional EM to identify the ones that displayed interphase nuclei, before proceeding with the CLEM workflow.
What was the key insight given by EM?
YS EM techniques enabled us to visualize for real what a bundle of other techniques could only suggest. For example, live fluorescence imaging strongly suggested the physical connection, continuity, between the AL and the nuclear envelope. Thanks to 3D EM (tomography and FIB-SEM) this continuity appeared obvious. FIB-SEM furthermore demonstrated the organization of the ER sheets relative to the nuclear envelope, an observation that led us to propose the model exposed in the paper. It was very important as well to demonstrate that the very dynamic fluorescent patches observed in vivo were indeed AL. CLEM clearly confirmed this.
“EM techniques enabled us to visualize for real what a bundle of other techniques could only suggest.”
Does the mechanism of AL-NPC insertion change during development, and is this important?
BH Yes, indeed. Insertion of entire AL is a maternal program and declines starting with zygotic induction. One reason might be simply the temporal constraint. AL insertion happens fast and could overcome the slow kinetics of classical interphase NPC assembly, as we know it from tissue culture cells. In the prolonged interphase 14 of Drosophila embryogenesis, AL diminish from the nuclear layer at the embryos’ cortex. The vast abundance of AL in oocytes and early embryos of many animal species also argues that they have a general role in the earliest stages of development.
Did you have a single ‘eureka!’ moment when everything came together or you got a particularly stunning result?
BH I would say two of them: First the photoconversion experiments where I could see that converted Nucleoporins distributed from an extranuclear spot into the nuclear envelope and there dissipated laterally. This really proved to me that AL do insert. And secondly when we could reveal insertion ultrastructurally in the FIB-SEM analysis.
Is there a loose end or surprising result in the paper you would particularly like to get to the bottom of?
MB Various experiments described in our paper show that nuclear envelope organisation before the start of transcription is very different from well-studied conditions in differentiated cells. That overexpression of the lamin B receptor is sufficient to make nuclei in the early embryo look as their counterparts at later stages I personally found very surprising.
And what are you working on now?
BH As always many questions emerge: for example how do the NE openings that seem critical for AL insertion form? How are they stabilised? To get a handle on this on a molecular level is my next goal.
Martin and Yannick, any planned future collaborations?
MB YES!
YS Yes definitely. This collaboration was one of these great and rewarding moments when we witness how the expertise from a service facility helps our colleagues to progress in their science. Martin’s and Bernhard’s enthusiasm and collaborative spirit successfully engaged everyone in this fantastic story. I can tell you they have plenty more, as exciting as this one!
“This collaboration was one of these great and rewarding moments when we witness how the expertise from a service facility helps our colleagues to progress in their science.”
Bernhard Hampoelz, Marie-Therese Mackmull, Pedro Machado, Paolo Ronchi, Khanh Huy Bui5, Nicole Schieber, Rachel Santarella-Mellwig, Aleksandar Necakov, Amparo Andrés-Pons, Jean Marc Philippe, Thomas Lecuit, Yannick Schwab, Martin Beck. Pre-assembled Nuclear Pores Insert into the Nuclear Envelope during Early Development. 2016. Cell. Volume 166, Issue 3, p664–678
You can catch up with the latest People behind the Papers here
A postdoctoral researcher position is available in the Plageman lab in the College of Optometry at the Ohio State University to elucidate mechanisms of vertebrate epithelial morphogenesis. The project will utilize the invaginating lens placode of the mouse as a model in combination with live-fluorescent microscopy. Candidates should have a Ph.D., a strong publication record and ideally have experience with mouse genetics, immunofluorescent imaging, and a background in molecular biology, biochemistry, embryology, and/or cell biology. Expertise in other model systems are also welcome. To apply, follow the instructions at the following link: https://www.jobsatosu.com/postings/72726
More information about the lab can be found at: https://u.osu.edu/plageman.3/ and inquiries about the position can be made directly to: plageman.3@osu.edu
 (No Ratings Yet)
(No Ratings Yet)
 (3 votes)
(3 votes)