(or: how to avoid misleading representations of statistical data)
Recently, a kickstarter project raised more than 3000€ in one month to campaign for banning the wrong usage of bar plots in scientific journals. This demonstrates two important points: a lot of the plots in scientific journals are quite misleading, and, a growing number of people feel very uneasy about this!
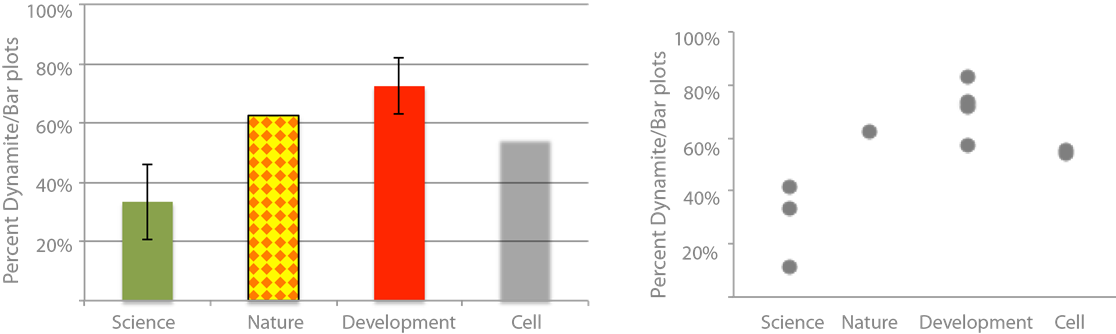
What exactly is wrong about bar plots? Nothing per se, but everything goes wrong if you use a bar plot for statistical data – this kind of plot species is also referred to as the dynamite plot: the bar being the detonator and the error range the firing cable (see figure)! We are talking about the famous vertical or horizontal boxes that often come in a dazzling array of colors or patterns, with big fat black outlines and overly prominent error bars.
Dynamite Plot Data Plot
Are they common? Very much so! My personal survey (footnote 1) of dynamite plots in scientific journals revealed that on average 30-60% of articles use them (see figure). These journals cover a wide range of subjects that include physics, meteorology or psychology where authors typically have rigorous training in applied mathematics. The prevalence of dynamite plots increases as we go towards more life science journals, where 50- 70% of articles are accompanied by a dynamite plot showing a statistical summary (footnote 2).
Most of us are completely accustomed to dynamite plots and happily use them, that is, until we see the light. From then on it is impossible to not hate them! Because it is so obvious they are misleading and they just make it harder than necessary to understand the data! And, as scientists, we thrive for clear and concise information!
The top reasons to avoid dynamite plots
They hide the real distribution of data. Do all samples cluster closely? Do they form two groups? Or is there one drastic outlier? Generally, we assume a normal distribution of the data around the mean where there might not be one! In my survey of dynamite plots per journal they were more or less normally distributed.
They hide the sample size. From the bar plot you would not have known that I probed one issue of Nature, two issues of Cell and four issues of Development! But for judging scientific data knowledge of the sample size is essential for a proper evaluation of the data! Too often we have to search for the n in axis labeling, figure text, the results, or the methods section to finally find this information. And sometimes it is omitted entirely. A clear understanding of sample size in my opinion is also critical for the review process of a paper and should be demanded by the reviewers! Not showing data, or only showing summary data, should be treated equally to cropping Western blot bands!
Many different data distributions lead to the same bar! See also the Anscombe quartet. Bar plots are not intended to show statistic distributions, they are for absolute numbers. And, by plotting the real data we also learn more about the biology!
Not quite convinced? Seeing is believing, check out this figure:
(c) Page Piccinini and the #barbarplots campain
Further information: watch the video of the kickstarter campaign (humor alert!) – ideally with your entire lab! And read this seminal paper on wrong usage of bar charts and this survey of their prevalence in biomedical journals!
Practical advice to avoid dynamite plots
Plot charts with statistical programing tool R. You have to either learn it, or be really nice to someone who knows it – if your PhD requires 3 boxplots, maybe invest in a friendly relationship with the bioinformatic geek in your department, a couple of coffees go a long way!
Learn how to make box plots in excel! (Hereand here is how, but it’s a bit tedious).
1) I probed the top10-articles of Nature in July, the three most recent volumes of Science (August), four issues of Development (Vol 138, 1:3(2011) and Jan 2016), and two issues of Cell journal from 2016 (Jan and August). I was very relaxed in my judgement and gave the benefit of doubt when I wasn’t sure. But I was rigorous when authors mixed right and wrong usage of bar plots. How does this even happen? Mix of co-authors and some know better than others?
2) Disclaimer: this does not mean the other articles have great figure design in any of the journals! I saw multiple uses of 3-dimensional pie charts, rainbow color schemes, other instances of unintentional usage of color, incomprehensible spider graphs and 3-dimensional heat maps! Maybe I will devote another blog post to those.
Here are the highlights from the current issue of Development:
Coordinating neuronal specification and differentiation
Neurogenesis – the process of making new neurons – is indispensable for normal development and for adult homeostasis and repair. Many of the signalling and transcriptional events that regulate the specification and differentiation of neural progenitor cells (NPCs) into neurons have been uncovered; however, how these events are coordinated at a post-translational level is not well defined. Now, on p.3085, Miho Matsuda, Ajay Chitnis and colleagues identify the protein Epb41l5 – an adaptor protein that links cytoplasmic proteins to specific membrane compartments – as a new regulator of neuronal differentiation in the developing zebrafish hindbrain. The authors first identify Epb41l5 in a yeast two-hybrid screen against Mib1, a key component of the Notch signalling pathway. Using Epb41l5-deficient embryos, the authors show that loss of Epb41l5 impairs neuronal differentiation, but that this can be partially rescued by knockdown of N-cadherin expression, suggesting a possible role for Epb41l5 in the disassembly of apical adherens junctions. In support of this, the authors further demonstrate that Mib binding to Epb41l5 facilitates its degradation and thus promotes apical adhesions, which may impair proper delamination and differentiation. The authors conclude by proposing a model whereby changes in Notch ligand levels that occur during neuronal differentiation protect Epb41l5 from Mib1-mediated degradation, thereby facilitating neuroepithelial detachment and subsequent differentiation.
Sall4 is dispensable for mouse pluripotency
In order to specify the correct lineage at the correct time, the developing embryo must maintain tight control over the gene regulatory networks that enact these changes. Sall4 has long been associated with the regulation of embryonic stem cell (ESC) self-renewal and differentiation; however, teasing out its precise role has been difficult. Now, on p. 3074, Brian Hendrich and colleagues present a comprehensive analysis of the role of Sall4 in self-renewal and differentiation, and shed light on the nature of its interaction with the NuRD complex during these events. Using a series of phenotypical and transcriptional analyses of double Sall4/1 knockout mouse ESCs (mESCs), the authors show that Sall1 and Sall4 are dispensable for ESC pluripotency but are required to repress neuronal differentiation. Remarkably, the authors observed the spontaneous production of neurons alongside self-renewing mESCs in the double knockout mESCs. Genome-wide analyses demonstrate that, although a small proportion of Sall4 does indeed interact with NuRD, Sall4 neither recruits nor functions through the NuRD complex. Rather, Sall4 is seen to bind to enhancer sequences along with the pluripotency-associated transcription factors Pou5f1, Nanog, Klf4 and Esrrb, which can result in either gene activation or repression. Together, these data shed light on a number of previously unresolved issues with regard to the function of Sall4 in mammalian development.
The asymmetry of asynchrony: new roles for CYB-3 in cell division
Regulation of the cell cycle is a crucial component of development, and has been linked to the execution of cell fate decisions in a wide range of developmental contexts. Although many of the molecular components involved in cell cycle progression have been identified, how these proteins are regulated and how their distribution and abundance can influence cell fate remains unclear. In this issue (p. 3119), Matthew Michael identifies cyclin B3 (CYB-3) as a key regulator of cell cycle timing in the developing C. elegans embryo. Using RNAi to knockdown CYB-3, Michael demonstrates that in the one-cell embryo CYB-3 controls not only mitotic entry but S-phase entry as well – a dual-action that is unique among cyclins. At the two-cell stage, the author shows that CYB-3 is asymmetrically distributed in a par-dependant manner such that somatic precursor cells inherit ∼2.5-fold more CYB-3 than do their germline precursor sister cells. The author uses maternal strains with varying copy numbers of cyb-3 to show how variations in the level of CYB-3 can affect the speed and synchrony of the cell cycle at the two-cell stage, suggesting a novel role for CYB-3 in regulating asynchronous cell cycling in the developing embryo. Together, these data advance our understanding of how the timing of cell division is differentially regulated in the early embryo.
PLUS:
Introducing cross-referee commenting in peer review
Following our recent community survey on priorities in peer review and online publishing, we are making changes to the journal, including some changes to our peer review process. Find out more by reading the Editorial on p. 3035
Slit-Robo signaling
Slits are secreted proteins that bind to Roundabout (Robo) receptors. Slit-Robo signaling is best known for mediating axon repulsion in the developing nervous system. However, in recent years the functional repertoire of Slits and Robo has expanded tremendously and Slit-Robo signaling has been linked to roles in neurogenesis, angiogenesis and cancer progression among other processes. Here, Heike Blockus andAlain Chédotalsummarize new insights into Slit-Robo evolutionary and system-dependent diversity, receptor-ligand interactions, signaling crosstalk and receptor activation. See the Development at a Glance poster article on p. 3037
Metabolism meets development at Wiston House
It is becoming increasingly clear that cellular metabolite levels regulate the activity of signaling pathways, and conversely that signaling pathways affect cellular physiology and growth via metabolic pathways. Thus, metabolism and signaling mutually influence each other. The recent Company of Biologists’ Workshop ‘Metabolism in Development and Disease’ brought together people studying signaling and development with people studying metabolism, particularly in a cancer context. Here, Aurelio Teleman discusses examples of talks that illustrated this principle. See the Meeting Review on p. 3045
Proliferation, survival and metabolism: the role of PI3K/AKT/mTOR signalling in pluripotency and cell fate determination
Phosphatidylinositide 3 kinases (PI3Ks) and their downstream mediators AKT and mammalian target of rapamycin (mTOR) constitute the core components of the PI3K/AKT/mTOR signalling cascade, regulating cell proliferation, survival and metabolism. Although these functions are well-defined in the context of tumorigenesis, recent studies – in particular those using pluripotent stem cells – have highlighted the importance of this pathway to development and cellular differentiation. Here, Jason Yu and Wei Cui review the role PI3K/AKT/mTOR signalling plays in the control of pluripotency and differentiation, with a particular focus on the molecular mechanisms underlying these functions. See the Review on p. 3050
The roles of microRNAs and siRNAs in mammalian spermatogenesis
MicroRNAs and siRNAs, both of which are AGO-bound small RNAs, are essential for mammalian spermatogenesis. Although their precise germline roles remain largely uncharacterized, recent discoveries suggest that they function in mechanisms beyond microRNA-mediated post-transcriptional control, playing roles in DNA repair and transcriptional regulation within the nucleus. Here, Andrew Grimson and colleagues discuss the latest findings regarding roles for AGO proteins and their associated small RNAs in the male germline. They also evaluate the emerging and differing roles for AGOs and AGO-bound small RNAs in the male and female germlines, suggesting potential reasons for these sexual dimorphisms. See the Review on p. 3061
Sense & Sensibility: niche signals regulate neural stem cells in an age-dependent manner
paper feature by Thomas Schwarz-Romond
Tissue specific stem cell niches provide lifelong support for adult stem cells. The cell-biological dissection of (adult neural) stem cell – niche interactions uncovered unexpected regulatory functions. These new results imply that stem cell niches actively sense (age-dependent) physiological changes and translate them into molecular cues to instruct stem cell activity.
The physiological control of stem cell activity is an intensely studied subject. Stem cells reside in so called niches, specialized compartments to nurture and protect the remarkable cellular properties of tissue-resident, totipotent cells. From a morphological perspective, the form and composition of tissue specific stem cell niches varies tremendously, being neatly adapted to accommodate the functional requirements in any given tissue. Clonal lineage-tracing strategies and conditional deletion experiments from many labs have accelerated our insights into stem cell hierarchies, best illustrated in the hematopoietic system and a growing list of epithelia stem cell compartments such as the skin, the mammary gland, the intestine or prostate1,2. At the same time, we are just beginning to capture the structural, cellular and micro-environmental components constituting tissue-specific stem cell niches3,4.
Another exciting stream of research suggests that stem cells in their respective niches are able to respond dynamically to changes in their physiological environment. In fact, GDF11 was characterized as a systemic factor to modulate muscle and neuronal stem cell function in an age- dependent manner. The same authors also revealed that GDF11 elicited some of these effects by functionally improving the vascular neurogenic niche5,6. Such data hint to contributions of stem cell niches beyond sole structural support and motivate ongoing research (i) addressing the complex cellular composition of tissue-specific stem cell niches, (ii) investigating active roles in sensing the physiological environment, and (iii) inquiries into niche components able to transform systemic signals into molecular cues to regulate niche-resident stem cells.
Along such conceptual preposition, Fiona Doetsch and her team7 explore the lateral ventricle choroid plexus (LVCP) as a putative novel component of the neural ventricular stem cell niche. A simple system to harvest conditioned medium from LVCP explants enables the authors to dissect the specific, cell-biological effects of the LVCP-secretome on neural stem cells as well as their progeny. Transcript- and proteomic evaluation of the LVCP and its secretome determines a rich
reservoir of factors known to promote neural stem cell (NSC)-quiescence, stem cell activation and proliferation, respectively. This includes chemokines, lymphokines, growth factors, hormones, ECM-components and their remodelers. Some of these (e.g. IL1B, NT3 and IGF2) had been implicated in the regulation of sub-ventricular zone (V-SVZ) stem cells before, functionally establishing the LVCP as new, and so far neglected part of the V-SVZ adult stem cell niche. Reaching further, the studies performed by Silva-Vargas et al. surface new regulators of V-SVZ stem cell activity, namely BMP5 and IGF1. Addressing the physiological significance of BMP5 and IGF1 in the control of neural stem cell activity, reveals their enrichment in young, compared to aged LVCP-secretomes, a crucial hint to age-associated fluctuations. Backed by these findings, the authors nominate the LVCP as new component of the V-SVZ neural stem cell niche. They conclude from the age-dependent changes in secretome composition that the LVCP acts as a sensor of physiological change, in turn adopting the composition of the secreted ‘cocktail’ as to accommodate environmental conditions. By extension, the results imply that niche-components in general could monitor systemic change and instruct stem cell activity in a context-dependent manner.
How do these ideas integrate with previous knowledge? It could easily be argued that the LVCP, as part of the choroid plexus, would morphological be in a prime spot to control NSC-behavior8. It was also described that the choroid plexus, a vascularized epithelium within the brain ventricle, produces the majority of the cerebrospinal fluid (CSF) and constitutes the blood-CSF barrier9. The fluid CSF-compartment had been reported to produce migratory cues for emerging neurons and factors maintaining stem cell quiescence10, while the choroid plexus itself was shown to dynamically respond to physiological inputs11. Finally, NSCs adjacent to the lateral ventricles extend projections into the CSF on one hand, and physically connect to blood vessels on the other12. Those data already provided a general scheme for the make-up of the tentative V-SVZ niche: highly connected NSCs, positioned in close proximity to the choroid plexus – the blood- cerebrospinal barrier, and hence at the interface to systemic circulation. Though seemingly distinct in morphology, such a conceptual composition appears reminiscent of the features previously described for the perivascular niche in the hematopoietic system3. Therefore, the new findings from Silva-Vargas et al.7 advance current views on the V-SVZ NSC-niche and help to generalize our understanding of stem cell-niche interactions.
Specifically, the far-reaching proposal of a niche-encoded ‘sensory- and molecular instructor’ function to control (neural) stem cell behavior significantly advances newly emerging concepts in contemporary stem cell biology and inspire eminent questions: could BMP5 and IGF1, characterized here as V-SVZ niche signals, have much broader functions? Do they operate in other stem cell niches, or even systemically, like GD11? Appreciating that specific stem cell niches employ various sets of instructive signals, new cell-biology might soon be disclosed, as so often when venturing into unknown territories13. For illustration, Wnt3, produced by (niche) paneth cells and acting in a gradient to control stemness, has recently be reported to spread in a membrane- bound fashion, instead of traveling by simple diffusion14. Further, similar-focused investigations assessing the inventory of niche components might accelerate new therapeutic interventions. This route has been elegantly exemplified in a study on muscle stem cells, with an artificially designed stem cell niche extending the quiescence of (cultured) stem cells and improving stem cell engraftment after transplantation15. In sum, continued efforts, which capitalize on the conceptual similarity and appreciate tissue specific differences in the cell-biological and molecular make up of stem cells niches will not only add exciting new chapters to basic biology textbooks but also generate tangible knowledge to inspire future therapies.
Wabik, A. & Jones, P.H. EMBO J. 34, 1164-79 (2015).
Part of the fascinating potential of stem cells is their ability to provide replacement cells and tissues to treat diseases. In order to do this most effectively, scientists need to be able to create differentiated cells quickly and accurately. However, making differentiated cells can take weeks to months and the resulting cells might be a mixture of the desired type of cell and others. This is because the steps between pluripotency and the differentiated state are not fully understood.
To tackle these obstacles, Irving L. Weissman and his team based at Stanford University studied how the cells of the middle layer of the embryo – the mesoderm – turn into 12 differentiated cell types. They mapped out 1) the intermediate cell types formed as a cell becomes more and more differentiated, 2) how these intermediates are formed via alternate paths at each branching point and 3) the signals that encourage and discourage cells to follow a path at each branching point.
Identifying the signals that cells receive to determine which path they follow allowed the researchers to make differentiated cells that were 80-99% the desired cell type in days rather than weeks or months.
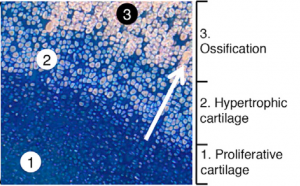
To show that these human cells behave as expected in a living model organism (‘in vivo’), they transplanted the cells that can produce bone, cartilage or smooth muscle (‘ventral somite progenitors’), into mice without an immune system.
The cells shown in the image are human cells inside of a mouse. The blue indicates cartilage and the light pink indicates bone. The process of cartilage turning into bone occurs as the blue cells secrete a scaffold of cartilage (‘proliferative cartilage’). The blue cells then enlarge and die (‘hypertrophic cartilage’), and the cartilage they leave behind turns into bone (‘ossification’). The light pink ossified cells are lined in diagonal rows, showing the structure of bone on a microscopic level.
The process developed by Weissman and his team mimics natural bone development and shows that the cells produced via differentiation can integrate into a living system properly. This suggests that, with further research, these cells could be transplanted into humans. With this faster, more efficient technique, therapies utilizing such differentiated cells could be significantly improved.
We are seeking outstanding postdoctoral candidates to join the Campas lab (Morphogenesis and Self-organization of Living Matter lab) at the University of California, Santa Barbara (UCSB). Our group combines theoretical and experimental approaches to study the spatiotemporal control of tissue mechanics during morphogenesis, using zebrafish as model organism. We have recently developed two microdroplet-based techniques that enable direct measurements of forces and mechanical properties (such as stiffness and fluidity) within developing 3D tissues, as well as allowing the application of controlled forces. Using these techniques, we are studying the molecular control of spatiotemporal variations in tissue mechanics and the role of mechanical feedback on cell behavior, all within developing embryos.
We are specifically seeking independent, passionate, and motivated applicants for a postdoctoral position to work on the mechanics of embryonic development in zebrafish. The candidate will be able to work in a collaborative manner with a highly interdisciplinary group of researchers, including theoretical physicists and engineers. A Ph.D. in the biological sciences, biophysics or related fields and at least 3 years of laboratory research experience in zebrafish development are required. Applicants with quantitative biology or biophysics backgrounds, in addition to experience in zebrafish development, will be considered positively.
This is a renewable, two-year position with full benefits, reappointed annually according to the performance of the candidate. Salary will be competitive and dependent on the level of experience of the candidate. Applicants should email a CV and a description of research interests to campas@engineering.ucsb.edu, and should also arrange for at least two references to submit letters of recommendation of their behalf. Applications submitted by October 31st 2016 will receive priority consideration, but the position will remain open until filled. Start date is flexible and could be as early as November 2016.
The University of California, Santa Barbara (UCSB) provides an exceptional, interdisciplinary and collaborative environment for scientists interested in quantitative biology and systems biology. Researchers at UCSB enjoy regular visits from world-leading scientists and workshops on quantitative biology and biophysics through the Kavli Institute for Theoretical Physics, in addition to exposure to the Summer School on Quantitative Biology.
The Campàs Lab (“Morphogenesis and Self-Organization of Living Matter” lab) at the University of California, Santa Barbara, works on tissue and organ morphogenesis in zebrafish. The lab focuses on understanding the role of mechanical cues in the development of embryonic structures using novel in vivo force transducers and actuators.
Our lab is seeking independent, passionate, and motivated applicants for a Research Specialist position. This position will assist with experiments, training, and general lab duties, as well as participate in multiple research projects. The candidate will be able to work in a collaborative manner with a highly interdisciplinary group of researchers, and will have many opportunities for co-authorships on published manuscripts. A master’s degree in the biological sciences or related field, and at least 3 years of laboratory research experience in zebrafish are required (lab experience can include graduate training). Knowledge of zebrafish molecular biology, imaging, microinjection, genetics and husbandry are necessary. Applicants with a Ph.D. will be considered positively.
This is a long-term, renewable, full-time position with benefits, reappointed annually. Anticipated begin date may be as soon as November 2016, but the start date is flexible. Salary will be competitive and dependent on the level of experience of the candidate. Applications must be submitted electronically at https://recruit.ap.ucsb.edu/apply/JPF00759 and must include a CV and research statement. Applicants will also need to arrange for at least 3 references to submit letters of recommendation of their behalf via the recruitment website. Applications submitted by October 31st 2016 will receive priority consideration, but the position will remain open until filled.
The department is especially interested in candidates who can contribute to the diversity and excellence of the academic community through research, teaching, and service. The University of California is an Equal Opportunity/Affirmative Action Employer and all qualified applicants will receive consideration for employment without regard to race, color, religion, sex, sexual orientation, gender identity, national origin, disability status, protected veteran status, or any other characteristic protected by law.
Forget about those large amounts of bottles containing thousands of flies, those huge piles of boxes containing different lineages of mice or large tanks filled with happy-hopping frogs. Also, forget about transgenic, mutant, knockout litters… what I am going to tell you is the routine of an emergent lab working (or, better, trying to) with emergent models that you probably haven’t heard about for a while.
My name is Emilio Lanna and I am a young faculty at the Biology Institute in Federal University of Bahia (UFBA) in Salvador, Bahia, Brazil. I coordinate the Embryology and Reproductive Biology lab (LEBR), which currently consists of four grad students and four undergrads; all of them focused in understand the evolution of sponge development and life history.
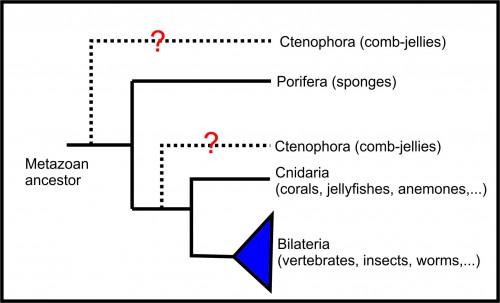
Sponges (phylum Porifera) are aquatic animals living in the bottom of the oceans (you can find some species in freshwater, too). They are morphologically very simple, having a body adapted to pump as much water as possible to get their aliment, get rid of toxic metabolites, and even reproduce. However, although morphologically simple, their genomes are complex and similar to those of other animals. It is very important to study the development of sponges if we are to understand the evolution of animals as whole, as this group is considered the sister-group of all other metazoans (Figure 1, BUT you may find divergent opinions about that, especially those that put ctenophorans as the sister-group of all metazoans, but this is a history for another time). Their development may also shed light on the ecology and conservation of those animals. And, from an anthropocentric perspective, sponges are a rich source of natural products with a broad range of potential application in pharmaceutics, cosmetics and bioproducts. Last, but not least, the sponges’ cells have the ability to differentiate into other cell types in any stage of their life cycle. Therefore, understanding the mechanisms of cell differentiation in these animals could help us to treat cancer and advance tissue regeneration therapies. Therefore, although they are mainly interesting from an evo-devo perspective, there is a potential for medical application for the knowledge generated studying sponge development.
Figure 1 – Metazoan phylogeny (see Lanna, 2015 for a better discussion about the phylogenetic relationship of the non-bilaterian phyla)
As stated before, we are working with emerging models. We are currently investigating ten different species belonging to two different classes and six orders of Porifera (Figure 2). Before starting our work, we knew nothing about the reproductive biology of these species. Even simple questions such as: “when?”, “how often?”, “how much?” and “how do they reproduce?” were never addressed by other investigators before. So, we are doing a lot of early and basic research here. To do that, the work in our lab always starts with the students heading to a gorgeous beach close to the campus (Figure 3). We usually plan the sampling looking at a tide table, in order to choose the time when the distance of the surface and the bottom is the lowest possible, because we are going to skin dive. No special gear is necessary, as the seawater in Salvador is always around 24-26 ºC (75-79 ºF). Diving is not always necessary, as there are some of the species that hang on nautical ropes in nearby harbour and others that are directly collected in a flat rocky substrate during the low tide. IMPORTANT! Before any sponge collection we have to register the sampling campaign in an online platform of the Brazilian Environmental Ministry.
Figure 2 – In situ photographs of some of the species of sponges investigated by our lab.
Figure 3 – Porto da Barra beach is located at the mouth of the Todos os Santos Bay, the 3rd largest bay in Brazil. It has warm and transparent waters throughout the year (credits: Creative Commons).
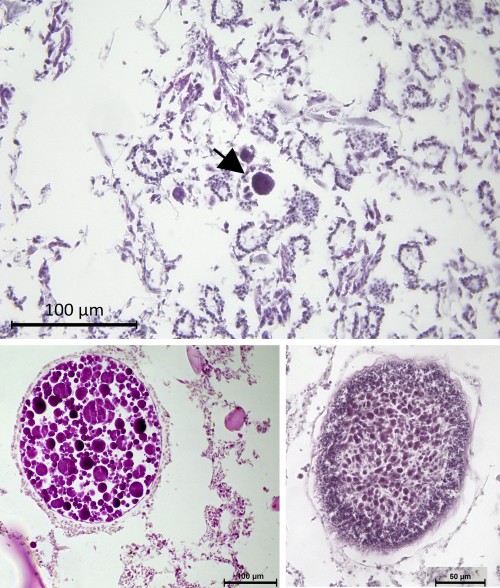
The dive can have different objectives and the after-dive work will depend on these. The most usual is to collect pieces of sponges for histological procedures. In this case, the samples are fixed in saline formaline and brought to the lab, where they are processed through standard protocols. The material is then observed with a compound microscope to search for the reproductive elements (eggs, sperm cells, embryos and larvae). Sponges lack any organs, and gonads or reproductive compartments are always absent. Gametes and embryos, when present, are found spread throughout the somatic tissue of the organism and the researcher has to quickly learn how to differentiate a choanocyte (a typical somatic cell) from spermatozoa and to find embryos among the choanocyte chambers (Figure 4). We can count these elements to answer the questions on when, how often and how much the species reproduce, but this procedure tells us little about the gametogenesis and embryogenesis.
Figure 4 – Histological preparations to study the reproduction of sponges. The upper panel shows an egg (arrow) among the choanocyte chambers. The right bottom panel is an embryo in late cleavages and the left bottom panels shows a parenchymella larvae.
To describe gametogenesis and embryogenesis, we are doing electron microscopy and planning three-dimensional reconstruction using a confocal microscope (using DAPI). For the electron microscopy (EM), we take two pieces of the same sponge: one is fixed for EM (with standard fixative cocktails) and other for histology. We first confirm that the reproductive elements are present through the histological slides, because it’s cheaper and routine in our lab. To prepare the samples for EM we have to take them to another lab in another institution (Fiocruz-BA). There, the wonderful crew of technicians do the hard work for us and we have only to check the semithin sections and then go to spend productive hours sitting in front of the screens of the scanning or transmission EM.
In the last few months we started another approach, which has been, at the same time, both challenging and very interesting. We are putting larval traps above the sponges in the field to collect the free-swimming larvae of these animals. We go to this beach few hours before the sunset and set the traps around the sponge. The traps lay there till the next morning when we dive again to collect conical tubes filled with 0.1-0.5 mm long larvae. These tubes are taken to the lab, where we observe the ciliated larvae swimming in circular pattern for hours. Then, with a lot of patience, we observe them to settle, metamorphose and start their development. We do that with direct observation, fixing in different periods and through time-lapse imaging. The larvae lack a functional aquiferous system (the one used to pump water). As the development of this trait is little understood, we are going to investigate the morphogenesis and dynamics of development of this system. We are currently staging the development of three species and are also planning to disturb their development applying drugs and physical disruptors.
As you could see till here, our lab is not what a canonical student of developmental biology might expect to find in a developmental biology lab. We are getting there, though. In a few months, we expect to sequence the transcriptome of different stages and parts of one of the species. After that, we are also planning to do some in situ hybridizations to check which cell type express key developmental genes. Until those issues become routine in our lab, they cannot be described here… so, I may come back in the future.
If you have any question or comment, please send me a message. I´ll be glad to chat with you!
Controlling differentiation using biophysical cues from development
Embryonic stem cells have the potential to become any cell type in the adult organism, but coaxing them to a specific fate continues to be a challenge for researchers. While many of the soluble signals involved in patterning the early embryo are well-established, only recently have tools been available to study how biophysical cues synchronize with other signals in the microenvironment to specify cell fate during development. Mechanical signals have been implicated in controlling stem cell fate in a number of contexts, but the molecular mechanisms involved are often not well-defined. By taking cues from embryonic development, we have identified a key role for mechanical signals in driving differentiation of human embryonic stem cells (hESCs) toward mesoderm, work that was recently published in Cell Stem Cell (Przybyla et al., 2016a). We used genetic and pharmacologic manipulation to identify molecular mechanisms whereby tissue tension directs stem cell fate and demonstrated how hESC differentiation protocols can be optimized to direct early progenitor specification.
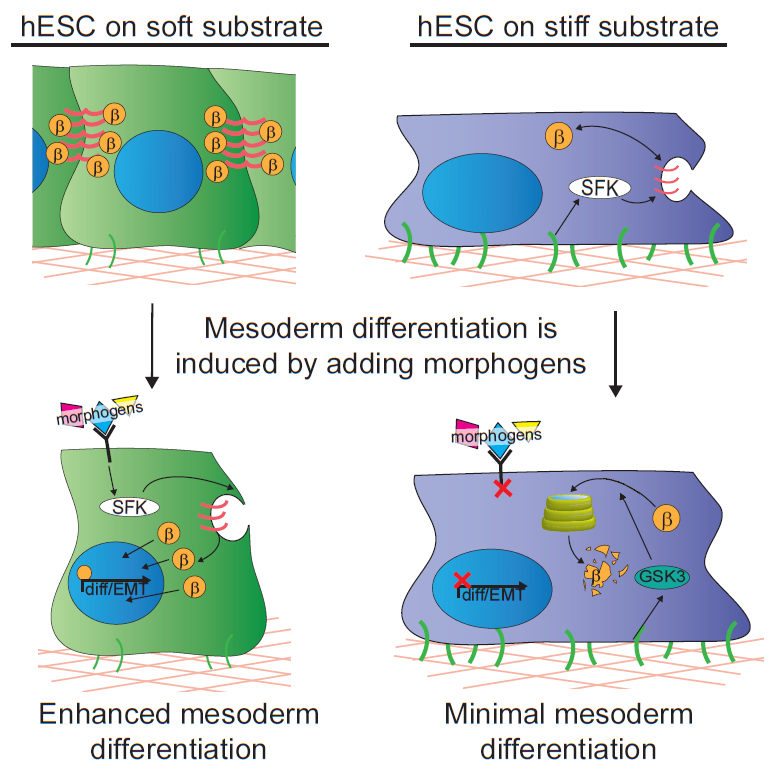
Substrate compliance can be tuned to alter the ability of hESCs to differentiate toward mesoderm.
Whether scientists are working to regenerate damaged organs or fix genetic defects in existing organs, a major bottleneck in regenerative medicine is generating differentiated cells that exhibit a structure/function that faithfully recapitulates that of the cells within the tissues in healthy adult organs. One approach has been to mimic the soluble cues and transcriptional signaling cells receive and integrate during embryonic development as they transition to tissue-specific differentiation in the adult organism. Yet cells in tissue also receive spatial-mechanical cues that are critical for directing their fate and maintaining the tissue function.
For more than a decade the Weaver laboratoryhas been exploring the role of biophysical cues in adult tissue development and homeostasis and malignancy, in order to clarify how the physical context of a cell in a tissue drives embryogenesis and influences stem cell fate specification. To this end, our experimental efforts have focused on examining how cell-cell and cell-extracellular matrix interactions and tissue geometry regulate early developmental decisions. We have been studying early gastrulation and mesoderm specification using human embryonic stem cells and a simplified polyacrylamide gel system combined with three dimensional cell printers. Our findings revealed that the biophysical microenvironment helps to synchronize embryogenesis to facilitate mesoderm progenitor differentiation.
The complexity of embryonic patterning
The first symmetry-breaking process in embryogenesis is gastrulation, when cells are reorganized and divided up into three germ layers that go on to derive all the structures and organs of the body. This process also marks the first epithelial-mesenchymal transition (EMT), as cells in the epiblast ingress into the newly formed primitive streak and lose their epithelial characteristics to form the mesoderm layer. A major question remains as to how cells know where to go and when, and what to do when they get there. Cells derive this information from the inputs they receive from their cellular and noncellular microenvironment. A major goal of our work is to begin to unravel the molecular mechanisms whereby spatial-mechanical cues regulate gastrulation.
Epithelial versus mesenchymal cell organization can affect the cell’s response to extrinsic cues
When cells switch between epithelial and mesenchymal states, their organization with respect to each other becomes dramatically altered. Cells in an epithelial state have strong cell-cell contacts and exhibit collective cell behaviors such as coordinated migration and mechanical connectivity, whereas cells in a mesenchymal state acquire a motile phenotype that is linked in part to stronger cell-extracellular matrix interactions. After the initial EMT during gastrulation that forms the mesoderm layer, mesoderm cells destined to different downstream fates undergo additional mesenchymal-epithelial transitions (METs) and EMTs as tissues are formed and cells migrate to their appropriate embryonic positions. The objective of our recent studies was to clarify how cell arrangements and rearrangements influenced the cellular response to mesoderm-inducing cues in the context of early embryonic gastrulation. To do this, we first developed a method to control both the hESC plating geometry and density and the mechanical signals presented to hESCs through their extracellular matrix substrate (Lakins et al., 2012; Przybyla et al., 2015). We then used this system to interrogate how cell context influences early mesoderm specification and is able to orchestrate a program of differentiation with striking parallels to embryonic gastrulation.
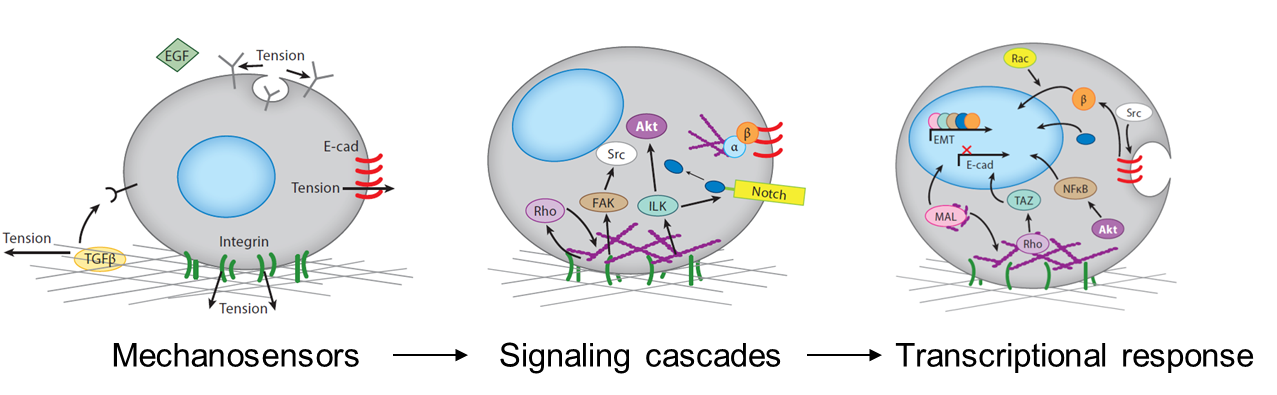
Mechanical signals can control transcriptional responses
The mechanical stimuli sensed by cells in the developing embryo include the stiffness and topography of their extracellular matrix, the forces exerted between cells, the shear stress applied by surrounding fluid flow, and mechanical strains resulting from interstitial and osmotic pressures. Cells respond to mechanical signals with integrated mechanosensitive machinery, including molecules that hold tension at the cell-cell and cell-substrate interfaces. Cell organization within tissues affects what mechanosensitive machinery is expressed and activated, thereby dictating how cells respond to other environmental cues (Przybyla et al., 2016b). For example, mesenchymal cells may initiate signaling pathways downstream of strong cell-matrix contacts at focal adhesions, whereas strong cell-cell junctions in epithelial cells may allow for efficient transmission of contact-based signals. Mechanical signals can also alter ligand (Giacomini et al., 2012; Wipff et al., 2007) or receptor (Boulant et al., 2011; Gauthier et al., 2011) accessibility, thereby enhancing or inhibiting downstream signaling pathways.
Mechanisms by which mechanical cues can be translated by cells into functional responses.
Tissues and organs all have an intrinsic stiffness that is likely to affect their functionality. When tissue stiffness is altered, as in the case of heart or liver fibrosis, the ability of the organ to function is dramatically compromised. Similarly, developing embryonic tissues have an intrinsic stiffness that rapidly changes during differentiation (Majkut et al., 2013). Though we believe that these intrinsic properties contribute to development as part of the signaling microenvironment that drives cell fate, this has not been demonstrated in a controlled way. Using engineered matrices to control substrate stiffness as a mechanical parameter allowed us to explore this phenomenon during stem cell differentiation in vitro, and relate this to processes that occur in vivo during embryonic development.
Stem cell fate can be controlled by substrate stiffness as a mechanical input
In 2006, Dennis Discher’s group reported that mesenchymal stem cell fate could be controlled by altering substrate stiffness (Engler et al., 2006). Since then, many studies have demonstrated that substrate compliance is able to influence mesenchymal, neuronal, and liver stem cell differentiation (Cozzolino et al., 2016; Gobaa et al., 2015; Saha et al., 2008) as well as stem cell self-renewal (Chowdhury et al., 2010; Gilbert et al., 2010) and they have implicated RhoGTPases and myosins as key regulators of these behaviors. Nevertheless, the relevance of these observations to human embryonic stem cell fate and the molecular mechanisms critical for eliciting these phenotypes remained ill defined.
In our study, we cultured human embryonic stem cells on soft substrates similar in compliance to a gastrulation-stage embryo (400Pa) and substrates that were stiffer by two orders of magnitude. We were able to show that hESCs maintain their ability to grow and self-renew regardless of substrate stiffness, but surprisingly we failed to observe any direct impact of substrate stiffness on hESC differentiation. Nevertheless, we did note a remarkable increase in their propensity to differentiate into mesoderm progenitors when they were stimulated with differentiation-promoting morphogens if grown on the extracellular substrates that mimicked the microenvironment of the embryo. Mesoderm differentiation occurs concomitant with the first EMT of development, so we concluded that it could represent a stage at which cells respond to mechanical inputs by altering their fate.
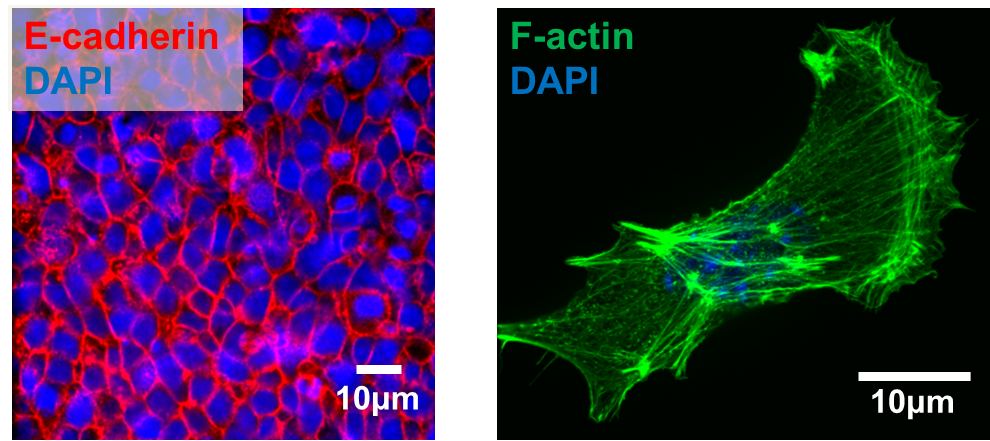
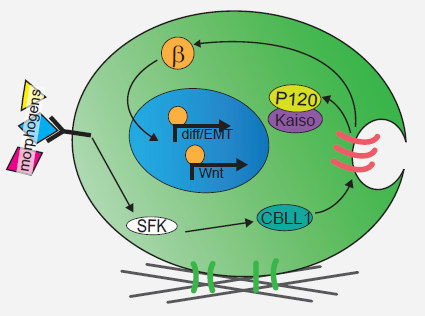
We next explored the molecular mechanisms underlying the enhanced mesoderm progenitor specification, finding that hESCs on embryo-like substrates coordinated several complementary pathways to initiate and stabilize mesoderm progenitor differentiation. First, β-catenin was not degraded in cells cultured on soft substrates, while rapid proteasomal degradation occurred in cells on stiff substrates due to increased Src and GSK3 activity. Second, cell-cell junctions were stabilized in cells on soft but not stiff substrates, creating colonies with enhanced epithelial characteristics, including circumferential actin and high levels of E-cadherin and β-catenin localized at these junctions. Third, the induction of differentiation caused controlled activation of signals, including P120-catenin-mediated Kaiso sequestration, that coordinately enhanced the ability of β-catenin to enter the nucleus and begin a program of transcription to initiate and reinforce mesoderm differentiation. By upregulating Wnts, this system propagated a feed-forward loop that mitigated expression of secreted Wnt inhibitors to ensure robust, efficient mesoderm differentiation, but only under circumstances where cells were primed on soft, embryo-like substrates. Our findings obtained using this simplified hESC system not only build on prior knowledge of Wnt-dependent mesoderm specification during embryogenesis derived from model organisms, but also definitively link Wnt-stimulated developmental programs to a tightly coordinated cytoskeletal-linked adhesion-directed mechanosignaling pathway.
Mechanism for how cells cultured on soft gels respond to differentiation cues to coordinately initiate mesoderm differentiation
Using an in vivo model of gastrulation, the chick embryo, we also demonstrated that some of the same signaling pathways were upregulated in cells undergoing gastrulation and differentiating toward mesoderm. This study therefore demonstrates a means by which soluble signals synchronize with mechanical cues to drive hESC fate decisions, implying that biophysical parameters sensed by cells in vivo are an important part of the extracellular microenvironment that contributes to cell specification during development.
Understanding the cues required for differentiation during development is a crucial step towards tissue engineering-based cell therapies
Stem cell-based regenerative therapies require generation of specific cell types that perform their physiological function and are arranged properly within tissues. Pluripotent stem cells are particularly important in regenerative medicine because patient-specific induced pluripotent stem cells (iPSCs) can be derived and used to fix genetic defects or regenerate diseased or injured organs without risk of immune rejection. Although protocols exist to differentiate pluripotent cells into specific cell types in vitro, many such protocols generate immature cells in which tissue-specific marker expression is much lower than levels found in corresponding primary adult cell cultures, and these cells often lack the ability to adequately perform the functions required in fully functional tissues.
By demonstrating how one aspect of the biophysical environment, substrate compliance, can be tuned to influence embryonic stem cell fate in a developmentally relevant way, we have taken a step towards recapitulating developmental processes in the dish, and we plan to extend these findings to explore how mesoderm cells incorporate biophysical cues to further differentiate toward a cardiac fate. We hope that recapitulating more aspects of the biophysical microenvironment of the embryo, including shear stresses and mechanical strains, will allow us to develop differentiation protocols that result in functional cells that will be useful in regenerative therapies. To this end, our objective is to further characterize the mechanical properties of the early embryo and develop additional innovative techniques to recapitulate the in vivo embryonic microenvironment in a dish.
References
Boulant, S., Kural, C., Zeeh, J.-C., Ubelmann, F., and Kirchhausen, T. (2011). Actin dynamics counteract membrane tension during clathrin-mediated endocytosis. Nat. Cell Biol. 13, 1124–1131.
Chowdhury, F., Li, Y., Poh, Y.-C., Yokohama-Tamaki, T., Wang, N., and Tanaka, T.S. (2010). Soft Substrates Promote Homogeneous Self-Renewal of Embryonic Stem Cells via Downregulating Cell-Matrix Tractions. PLoS ONE 5, e15655.
Cozzolino, A.M., Noce, V., Battistelli, C., Marchetti, A., Grassi, G., Cicchini, C., Tripodi, M., and Amicone, L. (2016). Modulating the Substrate Stiffness to Manipulate Differentiation of Resident Liver Stem Cells and to Improve the Differentiation State of Hepatocytes. Stem Cells Int. 2016, 5481493.
Engler, A.J., Sen, S., Sweeney, H.L., and Discher, D.E. (2006). Matrix Elasticity Directs Stem Cell Lineage Specification. Cell 126, 677–689.
Gauthier, N.C., Fardin, M.A., Roca-Cusachs, P., and Sheetz, M.P. (2011). Temporary increase in plasma membrane tension coordinates the activation of exocytosis and contraction during cell spreading. Proc. Natl. Acad. Sci. U. S. A. 108, 14467–14472.
Giacomini, M.M., Travis, M.A., Kudo, M., and Sheppard, D. (2012). Epithelial cells utilize cortical actin/myosin to activate latent TGF-β through integrin α(v)β(6)-dependent physical force. Exp. Cell Res. 318, 716–722.
Gobaa, S., Hoehnel, S., and Lutolf, M.P. (2015). Substrate elasticity modulates the responsiveness of mesenchymal stem cells to commitment cues. Integr. Biol. Quant. Biosci. Nano Macro 7, 1135–1142.
Lakins, J.N., Chin, A.R., and Weaver, V.M. (2012). Exploring the link between human embryonic stem cell organization and fate using tension-calibrated extracellular matrix functionalized polyacrylamide gels. Methods Mol. Biol. Clifton NJ 916, 317–350.
Majkut, S., Idema, T., Swift, J., Krieger, C., Liu, A., and Discher, D.E. (2013). Heart-specific stiffening in early embryos parallels matrix and myosin expression to optimize beating. Curr. Biol. CB 23, 2434–2439.
Przybyla, L., Lakins, J.N., Sunyer, R., Trepat, X., and Weaver, V.M. (2015). Monitoring developmental force distributions in reconstituted embryonic epithelia. Methods 94, 101-113.
Przybyla, L., Lakins, J.N., and Weaver, V.M. (2016a). Tissue Mechanics Orchestrate Wnt-Dependent Human Embryonic Stem Cell Differentiation. Cell Stem Cell, in press.
Przybyla, L., Muncie, J.M., and Weaver, V.M. (2016b). Mechanical Control of Epithelial-to-Mesenchymal Transitions in Development and Cancer. Annu. Rev. Cell Dev. Biol. 32, in press.
Saha, K., Keung, A.J., Irwin, E.F., Li, Y., Little, L., Schaffer, D.V., and Healy, K.E. (2008). Substrate modulus directs neural stem cell behavior. Biophys. J. 95, 4426–4438.
Wipff, P.-J., Rifkin, D.B., Meister, J.-J., and Hinz, B. (2007). Myofibroblast contraction activates latent TGF-beta1 from the extracellular matrix. J. Cell Biol. 179, 1311–1323.
Mathew Tata was the winner of the BSDB poster competition at the BSDB-BSCB 2016 meeting in Warwick. His prize was a trip to Boston to attend the SDB-ISD meeting in August, where he caught up with the SDB poster prize winner Yusuff Abdu to continue the BSDB-SDB interview chain (have a look at our previous interviews, which started way back in 2012). Yusuff’s prize in turn is to attend the BSDB-BSCB meeting in April, 2017.
The Node listened in as the two prize-winning graduate students chatted over a drink on the final night in Boston.
Yusuff, congratulations on winning the SDB poster prize! Can you tell us a little about the lab you work in?
Thank you very much. I’m inJeremy Nance’s lab, he’s a developmental biologist at NYU, and the lab studies a lot of events in the c. elegans embryo. Some people work on polarity and PAR proteins, but I study the development of the primordial germ cells.
And how long have you been in Jeremy’s lab?
Five years, just starting my sixth.
Can you tell us about your poster?
One common feature in development is this intimate interaction between germ cells and the endoderm, and I feel like we haven’t really fully appreciated it. My work has been looking at a particular form of germ cell remodelling driven by the endoderm: we basically found that parts of the germ cells are eaten by the endoderm, and we’re trying to figure out why that is. It appears to be important in some way for germ cell development.
In essence, the poster is about the active remodelling of germ cells by endodermal cells.
What would say is the single experiment or finding that you are most proud of?
That’s a tough one…This process we study occurs when the embryo is moving rapidly within its eggshell, so the imaging is tough and challenging. We were interested in these huge protrusions the germ cells send out, called lobes, and whether some of them remained attached to the cell body or not. To do this, we basically had to paralyse these embryos and do FRAP on them, photobleaching the lobes and seeing if there is any recovery from the cell body…so I call it imaging acrobatics: you have to keep them alive, but still enough, and do fancy imaging on top of that. I didn’t think it was going to work, but it did!
So what’s next for you?
After I get my doctorate, a postdoc: I think I’ll stay with worms, they’re a great organism to work with and we’re getting new tools everyday. There’s still a lot of weird stuff going on in worms that is yet to be understood: I’d encourage anyone who doesn’t work on them to consider a worm lab in the future.
And are you looking forward to the BSDB next year?
I am, yes: I’ve heard some great things about it.
Well it was a pleasure to meet you, and congratulations again!
We invite applications to this permanent full time appointment in the School of Biological Sciences. The successful candidate will be expected to make a significant contribution to biological research in the School and College, and to develop a research programme in their specialist area that will attract external funding and international recognition. They will also contribute to the teaching of biology and related subjects at undergraduate and postgraduate levels.
The successful candidate will be educated to PhD standard or equivalent and have previous experience across a broad range of sub-disciplines within Biological Sciences. To match our vision for the development of biology in the school we would particularly welcome candidates to carry out research to complement our existing areas of expertise (Animal Physiology; Behaviour and Conservation; Wetlands, Biogeochemistry and Plant Science; Microbiology, Parasitology and Biotechnology). From a teaching perspective, applicants should be prepared to contribute to areas that could include: physiology, immunology, biochemistry, human/primate biology, biotechnology, systems biology, quantitative biology, bioveterinary science or forensic biology.
In addition, the post holder will be expected to make a strong contribution to our existing ethos of inter-disciplinarity and team work in research and teaching, enhancing and complementing our existing area of expertise.
The appointment will be made in the range of Lecturer 1 £31,656 – £37,768 (Grade 7) or Lecturer 2 £38,896 – £46,414 (Grade 8) per annum, depending on previous experience.
Informal enquiries can be made by contacting Prof Chris Freeman (tel: +44 (0) 1248 382353, e-mail: c.freeman@bangor.ac.uk
Closing date for applications: 9th September 2016. Interviews will be scheduled shortly after 21st September 2016.

 (11 votes)
(11 votes)


 Slits are secreted proteins that bind to Roundabout (Robo) receptors. Slit-Robo signaling is best known for mediating axon repulsion in the developing nervous system. However, in recent years the functional repertoire of Slits and Robo has expanded tremendously and Slit-Robo signaling has been linked to roles in neurogenesis, angiogenesis and cancer progression among other processes. Here,
Slits are secreted proteins that bind to Roundabout (Robo) receptors. Slit-Robo signaling is best known for mediating axon repulsion in the developing nervous system. However, in recent years the functional repertoire of Slits and Robo has expanded tremendously and Slit-Robo signaling has been linked to roles in neurogenesis, angiogenesis and cancer progression among other processes. Here,  It is becoming increasingly clear that cellular metabolite levels regulate the activity of signaling pathways, and conversely that signaling pathways affect cellular physiology and growth via metabolic pathways. Thus, metabolism and signaling mutually influence each other. The recent Company of Biologists’ Workshop ‘Metabolism in Development and Disease’ brought together people studying signaling and development with people studying metabolism, particularly in a cancer context. Here, Aurelio Teleman discusses examples of talks that illustrated this principle. See the Meeting Review on p.
It is becoming increasingly clear that cellular metabolite levels regulate the activity of signaling pathways, and conversely that signaling pathways affect cellular physiology and growth via metabolic pathways. Thus, metabolism and signaling mutually influence each other. The recent Company of Biologists’ Workshop ‘Metabolism in Development and Disease’ brought together people studying signaling and development with people studying metabolism, particularly in a cancer context. Here, Aurelio Teleman discusses examples of talks that illustrated this principle. See the Meeting Review on p.  Phosphatidylinositide 3 kinases (PI3Ks) and their downstream mediators AKT and mammalian target of rapamycin (mTOR) constitute the core components of the PI3K/AKT/mTOR signalling cascade, regulating cell proliferation, survival and metabolism. Although these functions are well-defined in the context of tumorigenesis, recent studies – in particular those using pluripotent stem cells – have highlighted the importance of this pathway to development and cellular differentiation. Here,
Phosphatidylinositide 3 kinases (PI3Ks) and their downstream mediators AKT and mammalian target of rapamycin (mTOR) constitute the core components of the PI3K/AKT/mTOR signalling cascade, regulating cell proliferation, survival and metabolism. Although these functions are well-defined in the context of tumorigenesis, recent studies – in particular those using pluripotent stem cells – have highlighted the importance of this pathway to development and cellular differentiation. Here,  MicroRNAs and siRNAs, both of which are AGO-bound small RNAs, are essential for mammalian spermatogenesis. Although their precise germline roles remain largely uncharacterized, recent discoveries suggest that they function in mechanisms beyond microRNA-mediated post-transcriptional control, playing roles in DNA repair and transcriptional regulation within the nucleus. Here, Andrew Grimson and colleagues discuss the latest findings regarding roles for AGO proteins and their associated small RNAs in the male germline. They also evaluate the emerging and differing roles for AGOs and AGO-bound small RNAs in the male and female germlines, suggesting potential reasons for these sexual dimorphisms. See the Review on p.
MicroRNAs and siRNAs, both of which are AGO-bound small RNAs, are essential for mammalian spermatogenesis. Although their precise germline roles remain largely uncharacterized, recent discoveries suggest that they function in mechanisms beyond microRNA-mediated post-transcriptional control, playing roles in DNA repair and transcriptional regulation within the nucleus. Here, Andrew Grimson and colleagues discuss the latest findings regarding roles for AGO proteins and their associated small RNAs in the male germline. They also evaluate the emerging and differing roles for AGOs and AGO-bound small RNAs in the male and female germlines, suggesting potential reasons for these sexual dimorphisms. See the Review on p.  (No Ratings Yet)
(No Ratings Yet)


 (3 votes)
(3 votes)