Forgotten classics: Cut and paste embryology
Posted by the Node, on 2 February 2017
Hörstadius, S. 1939. The mechanics of sea urchin development, studied by operative methods. Biological Reviews 14(2):132-179.
Recommended by Bob Goldstein, University of North Carolina at Chapel Hill
Sven Hörstadius stands alongside the likes of Boveri, Spemann, Mangold and Driesch as a giant of experimental embryology in the first half of the twentieth century. While his 1950 treatise on the neural crest in vertebrate head development became an early bible for the field, he is probably best known for his work on sea urchins. His 1939 manuscript summarises a whole suite of work from himself and others dealing with various kinds of determination: of the axes of the embryo, of cell fate by instruction or position, and of ‘species character’ by the cytoplasm or the nucleus.

The focus is on the purple sea urchin, Paracentrotus lividus, which Hörstadius studied during trips to the Stazione Zoologica in Naples, once a haunt of Driesch and Boveri and still an active centre of research. P. lividus was a particularly useful urchin species because the eggs and early embryos have a vegetal pigment band, allowing polarity to be easily traced. In previous decades the basics of its development had been sketched out, including where successive cleavage planes lie in the cell, the movements of gastrulation, and fate of some blastomeres. However, while fundamental rules of development could be inferred from observation alone, Hörstadius was an experimentalist, and intervened in development in a variety of creative ways to investigate determination. Like Rosa Beddington, subject of the previous post in this series, he was known as an expert dissector: he could isolate blastomeres with fine glass needles, and create chimeras by fusing part of one animal to part of another and placing a small glass ball on top “to give the necessary pressure”. His 1939 paper is full of this sort of cut and paste embryology, and the outcomes are often unexpected (“The following is a very strange phenomenon…”).
The paper also devotes a lot of time accounting for contradictory results from other researchers. In particular, there seems to be a long standing beef with one Leopold von Ubisch, with some wonderfully formal put-downs:
“von Ubisch does not admit the possibility that halves of equatorial and subequatorial eggs are different. This is strange…” (p162)
“von Ubisch concludes that the cytoplasm has no influence on the species character of the skeleton. I do not find this conclusion convincing” (p169)
“The diagram of Fig. 9 has been criticized by von Ubisch. But his objections were, it seems to me, anticipated in the original paper” (p158)
As a bleak reminder of the historical context of the work, von Ubisch was forced out of his chair at Münster University for being non-Aryan four years before Hörstadius’ paper came out.
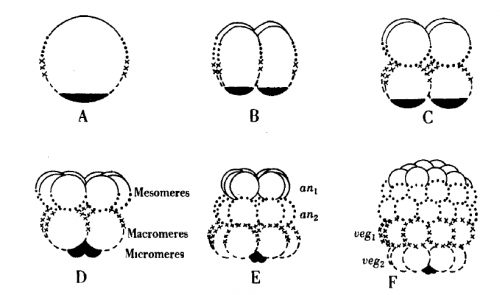
Among the flood of experiments and ideas that Hörstadius describes, some stand out. The first results section addresses the control of spindle position and orientation in the divisions that make the 16 cell embryo with its characteristic micromeres at the vegetal pole. These divisions can be delayed by shaking or adding diluted sea water; once this delay is relieved, the embryo starts dividing again. However, instead of picking up where they left off, the embryos would often skip a cell division such that, for instance, a delayed 4 cell embryo would divide to give micromeres (column B in the figure below).
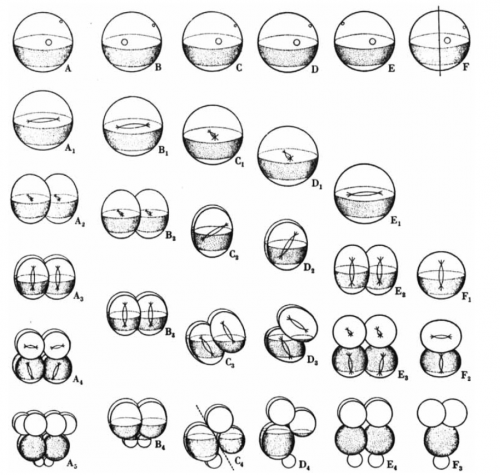
Hörstadius proposed that the micromere division is determined by the ‘activation’ of material in the vegetal cytoplasm, and that this activation occurs independently of how many cell divisions had occurred. After a certain amount of time, the embryo wants to make micromeres, irrespective of the number of divisions completed, and changes spindle position to do so. There is a kind of ‘cleavage clock’ which sets the type of cell division that occurs next.
The next three sections concern the embryonic axes. Hörstadius began his career under the supervision of John Runnström, who proposed that the sea urchin egg held two gradients, one emanating from the animal and one from the vegetal pole. The gradients “interact mutually and are partially hostile to each other”, and a cell’s fate is dependent on the relative strength of each of the gradients in the cytoplasm it inherits. Before the molecular biology revolution, before experimental and what was then called ‘chemical’ embryology were united, the cytoplasm had “qualities” or “forces” that influenced development.
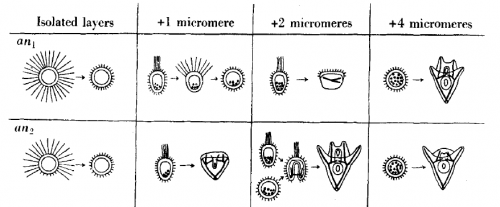
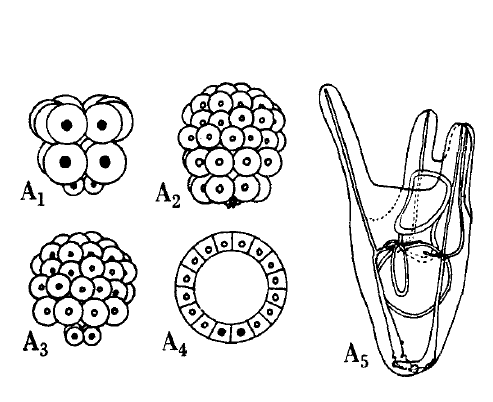
One of the ways Hörstadius investigated the double gradient hypothesis was to add micromeres (the inheritors of the most vegetal, and hence most ‘active’ cytoplasm) to layers of animal cells. These layers, if isolated and left alone, go on to form useless ciliated balls. Remarkably, add enough micromeres and you could rescue normal development, and gradually increasing the micromere number led to progressively ‘better’ development. Along with the results of various other experiments involving adding bits of one embryo to bits of another, this implied that the amount of animal and vegetal material you start with is crucial to the outcome. To explain these results, Hörstadius
“…assume[d] an animal and a vegetative gradient, both reaching the opposite pole and progressively diminishing. The animal and the vegetative qualities or forces have to interact in order to bring about normal differentiations, e.g. vegetative influences are necessary for the formation of ciliated band and stomodaeum, animal ones for gastrulation and skeleton formation, and so on. The differentiation depends – within wide limits – upon the relative amounts of animal or vegetative material present.” (p173)
While Lewis Wolpert used these experiments to inform his models of positional information, half a century later the picture had changed as Eric Davidson and others challenged the idea of a double gradient. In the updated model, specification is conditional, and mediated by successive interactions between micromeres and blastomeres, the micromeres having been initially autonomously specified by maternally supplied factors. The model combines modern data on signalling pathways and gene expression with a reinterpretation of Hörstadius’ classic experiments and their insight that cell interactions could direct development.
The paper ends with the question of the role of the nucleus in inheritance. Hörstadius took up work originally started by Boveri half a century earlier, removing the nucleus from eggs of one species and fertilising them with the sperm of another to make ‘heterosperm merogones’. The nucleus coming from one species, and the cytoplasm the other, he could ask which species the larva ended up looking like, and hence where ‘character’ was determined. In one combination, the merogone followed the characteristics of the species from which the nucleus was taken, but due to high variability in other combinations, Hörstadius had to be cautious in his intepretation:
“…the nucleus obviously has a positive effect on the species character. It is to be regretted that the characters in question are not defined sharply enough to permit of a conclusion as to the possible role of the cytoplasm.” (p172)
Even treading this lightly, the results are consistent with the work of Boveri, Spemann, Waddington, Wilson and Stevens, all of which put the nucleus, and the mysterious substance contained within it, in the driving seat of development and heredity.
The paper is testament to the power of cut and paste embryology, and the rich potential of the sea urchin embryo as a model. We can leave it Carl Olaf Jacobsen, one of only two graduate students trained by Hörstadius during his long time as a Professor in Uppsala, to summarise his supervisor’s legacy:
“…his experiments on sea urchin larvae shed light on a couple of the most central findings in developmental biology, namely that the uneven distribution of the egg-cell contents give rise to early embryo cells with shifting qualities, and that communication between these cells has an essential role in the differentiation process.”
Thoughts from the field
Bob Goldstein, University of North Carolina, Chapel Hill
This paper’s a fun read because it’s dense with simple and clever experiments. And many of the experiments built a foundation for our current understanding of how animal development works. But I love it most of all because it’s a great example of old literature that includes questions that we, as a field, forgot were fundamental questions. How does Hörstadius’ cleavage clock work? I think we still don’t know.
David McClay, Duke University, North Carolina
Hörstadius’ work was the inspiration for much of what I have done over the past 30 years. The following information was circulated over the years first by my mentor who had met Hörstadius, and more stories were circulated years ago when there was a festscrhriften thrown in honor of Hörstadius’ life in Stockholm. Unfortunately it was years after his death. Apparently until after his death the Swedes didn’t realize how well known Hörstadius actually was.
The post mentions that Hörstadius worked under Runnström. That was true and it was the cause for several things in his life. The Swedish system had very few professors so Horstadius worked under Runnström for most, if not his entire career. For that reason the Swedes never considered Hörstadius as particularly famous because he was not in charge. Ruunström was also involved in Hörstadius’ science so I was told. Runnström was a major proponent of morphogenetic gradients, especially double gradients. Consequently, the double gradients that are part of Hörstadius’ sea urchin work fit the ideas of Runnström – or perhaps were strongly suggested by him. Perhaps it was only gossip, but I heard from several senior scientists that Hörstadius never really believed that double gradient nonsense but since he was in Runnström’s lab that was part of the cost. He was later admitted to the Swedish Academy and to the Royal Academy largely for his work on neural crest, but the sea urchin work has stood the test of time with a greater presence than the neural crest work.
Höstadius was praised for his microsurgical abilities. He was the only one in the world who could do those remarkable dissections of the tiny sea urchin embryos and the cut and paste experiments were considered amazing feats. That was how I was initially taught about his work. He taught himself to dissect on the stage of a compound microscope. That meant he had to teach himself to move his needles in the opposite direction relative to what was seeing through the objective. His dissection tray was a piece of photographic film on the bottom of a shallow petri dish. That was because the photographic film had a coating of gelatin on it which was just thick enough to score a trough in which his embryos would be placed. Then, all of his dissections were done by hand. By contrast, I do all the same dissections but in my case I have major help from micro manipulators, a dissection microscope which shows movements in the same direction as what I can see though the objective, and I can record everything with a camera whereas he had to draw everything. As I indicated, he had great hands in addition to a perceptive brain.
† Further usage of any Wiley content that appears on this website is strictly prohibited without permission from John Wiley & Sons, Inc. Please contact Wiley’s Permissions Department either via email: permissions@wiley.com or use the RightsLink service by clicking on the ‘Request Permission’ link accompanying this article on Wiley Online Library (www.onlinelibrary.wiley.com)”
Aidan Maartens
This post is part of a series on forgotten classics of developmental biology. You can read the introduction to the series here and read other posts in this series here. We also would love to hear suggestions for future Forgotten Classics – let us know in the comments box.
 (8 votes)
(8 votes)Get involved
Create an account or log in to post your story on the Node.
Sign up for emails
Subscribe to our mailing lists.