A day in the life of a Termite lab
Posted by Kohei Oguchi, on 31 May 2019
How do genes and their environment interact during development and evolution to generate phenotypic diversity? To answer these questions in the Miura lab, by focusing on diverse animal taxa, we are studying physiological and developmental mechanisms of phenotypic changes in animal life cycles in response to environmental shifts. By the way, I’m Kohei Oguchi, a third year PhD Candidate and a member of Toru Miura’s lab at Tokyo University. Among diverse animal taxa, we have been especially focusing on social insects, “termites”.
Termites
Termites are insects belonging to a major group of social insects with divisions of labors among morphologically and behaviorally differentiated individuals (i.e., castes). In termite colonies, although all individuals share the same genetic background, there are several types of castes such as reproductives, workers and soldiers (Fig. 1A, B). These caste differentiations and caste ratios are regulated by individual interactions among colony members to organize highly sophisticated societies. Therefore, researches on termites help us to understand how genes and environments interact during development, i.e., caste differentiation, and how this interaction generates novel phenotypes, i.e., castes.
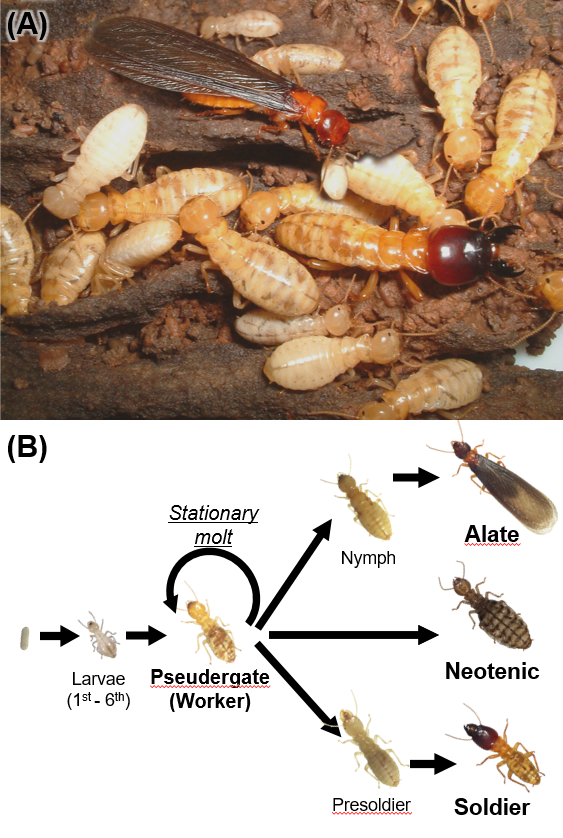
Recently, we identified a single gene which is responsible for the mandibular elongation that are seen specifically during soldier differentiation (Fig. 2A). To find genes specifically expressed in mandibles during the soldier differentiation processes, we performed gene expression analysis by RT-qPCR and immunostaining, and finally unraveled that dachshund (dac) was the gene we wanted to know (Fig. 2B). Consistently, RNA interference of dac expression inhibited the soldier-specific mandibular elongation (Fig. 2C).
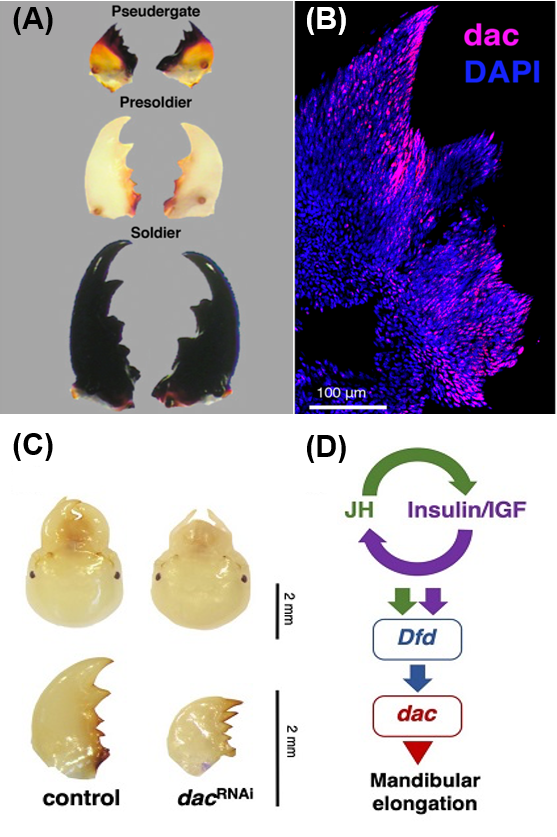
Moreover, we revealed that dac expression was regulated under hormonal factors and Hox genes (Sugime et al., 2019; Fig. 2D). In some previous research, interactions among colony members are known to trigger the physiological changes such as the elevation of juvenile hormone titers leading to soldier differentiation (e.g. Cornette et al., 2008). In addition to JH, insulin signaling is also shown to be responsible for soldier morphogenesis (Hattori et al., 2013). Depletion of juvenile hormone receptor, insulin receptor or particular Hox genes all lead to reduced dac expression, shedding light on the epistatic relationships regulating the dac activity. Therefore, our study strongly suggests that the cross talks between temporal factors, i.e., hormonal pathways, and spatial factors, i.e., Hox genes, lead the caste-specific developmental modifications that are seen during caste differentiation in termites.
Typical day
To achieve these findings, in a typical day, we spend a lot of time on the workbench with several types of termite samples. For gene expression and immunostaining analysis we use several pieces of equipment such as RT-qPCR and a confocal laser microscope (Fig. 3A, B). We also spend on time with live termite individuals for behavioral observations and experimental manipulations such as caste induction and micro injection (Fig. 3C).

In our focal termite species damp-wood termite Hodotermopsis sjostedti, several castes can easily be induced. In the reproductive caste induction, we manipulate sex and caste ratio of experimental colonies. On the other hand, soldier differentiation can be easily induced by the application of juvenile hormone. In addition, we often have discussions and seminars to discuss our research progresses and experimental plans (Fig. 3D). In every May, we go for collection trips for termite colonies to the Yakushima Island, which is a subtropical island located south of Kyushu, Japan.
Field day in the forest
Our focal termite species is the damp-wood termite Hodotermopsis sjostedti, that lives in rotten trees as its common name suggests. In Japan, this species is distributed in a subtropical forest of southern islands. My supervisor, Prof. Toru Miura, has been sampling termite colonies at Yakushima island every year for nearly 20 years. Soon after arrival Yakushima island, we discuss about schedules and searching area with a spread map. After discussion, we entry the forest and search the rotten trees. We walk and climb through the forest carefully to search for termite colonies, in addition to avoid any dangerous animals such as Mamushi pit vipers (Fig. 4A). When we break rotten trees with a hammer, we occasionally find termite individuals (Fig. 4B). Then, we cut and split the log to find a whole termite colony, that is then separated into containers, by cutting the wood with a saw and an axe. Sometimes, giant centipedes appear from the log (Fig. 4C). During the field sampling we spend a whole day as if we are lumberjacks. To avoid mixing colonies, we carefully labeled the colony number and identification, and send them to our laboratory. After such physical labour, we have fun by bathing in hot springs and drinking “Mitake”, traditional distilled spirit.

Field day in the ocean
Recently, our lab moved to Misaki Marine Biological Station at the University of Tokyo (Fig. 5A, B) from Hokkaido. Two years ago, our environment was largely changed to marine laboratory, so the focus of our studies has expanded to fascinating marine animals which provide us tremendous implications on development and evolution. We are now studying diverse taxa across animal phyla, and trying to establish rearing systems for future analysis to reveal physiological and developmental aspects of flexible phenotypic changes in animal life cycles.

Misaki Marine Biological Station is one of the oldest marine stations in the world (since 1886). It is located at the tip of the Miura Peninsula in Kanagawa Prefecture, facing the Sagami Bay that shows interesting geological features and a rich diversity of marine fauna (Fig. 5A, C). We are constantly sampling diverse animal species from the ocean throughout a year. We collect a number of marine creatures by walking with waders, snorkeling or scuba diving (Fig. 6A). After sampling, we maintain those creatures in the lab to use for experiments. Now, we are working on ten species of animals belonging to eight phyla (Fig. 6B).

References
Cornette R, Gotoh H, Koshikawa S, Miura T (2008) Juvenile hormone titers and caste differentiation in the damp-wood termite Hodotermopsis sjostedti (Isoptera, Termopsidae). Journal of Insect Physiology 54:6
Hattori, A, Sugime, Y, Sasa, C, Miyakawa, H, Ishikawa, Y, Miyazaki, S, Okada, Y, Cornette, R, Lavine, LC, Emlen, DJ, Koshikawa, S, Miura, T (2013) Soldier morphogenesis in the damp‐wood termite is regulated by the insulin signaling pathway. J. Exp. Zool. (Mol. Dev. Evol.) 320B: 295– 306.
Sugime Y, Oguchi K, Gotoh H, Hayashi Y, Matsunami M, Shigenobu S, Koshikawa S, Miura T (2019) Termite soldier mandibles are elongated by dachshund under hormonal and Hox gene controls. Development 146: dev171942.
Be sure to check out the other 40 (!) articles in our ‘A Day in the Life‘ series
 (4 votes)
(4 votes)Get involved
Create an account or log in to post your story on the Node.
Sign up for emails
Subscribe to our mailing lists.