Adhesion and signaling by Collagen IV intercellular concentrations (CIVICs)
Posted by Dai Jianli, on 16 October 2017
The Great Wall of Collagen IV
During the long history of evolution, the key innovation that gave rise to animals with true tissues was the extracellular matrix, very conserved from sponges to humans [1]. Before I started my PhD in the lab of Jose C. Pastor-Pareja at Tsinghua University in Beijing, he had described how the matrix protein Collagen IV is secreted and deposited into the matrix basement membranes (BMs) of the Drosophila fruit fly larva for shaping tissues and organs [2]. This research attracted my attention since I felt I’d like to understand the process and functional significance of extracellular matrices in building tissues. In my mind, extracellular matrices such as the BM were like the Great Wall (Figure 1), shaping, delimiting and defining tissues while keeping them from invasive damage.

Basement membranes are specialized planar matrices underlying epithelial and endothelial tissues, and surrounding nerves, muscles and adipocytes [3,4]. Collagen IV, thought to be an exclusive BM component, is a heterotrimer made of three alpha chains. These trimers can interact with each other to form a higher order scaffold. Despite a lot of research into extracellular matrices and a great deal of biochemical characterization, there are still many mysteries for developmental and cell biologists to solve. For instance, the hierarchy of assembly of BM components in vivo, why basement membranes are continuous planar layers rather than more amorphous structures or how matrices maintain the diverse organization of tissues and impact their physiology. In both flies and humans, adipose cells, called fat body cells in Drosophila,n IV, tissue morphog are cells of mesodermal origin lacking epithelial structure and yet they form true compact tissues. Our recent paper published in Current Biology shows how Collagen IV-containing structures different from BMs hold adipocytes together.
How it all started
One of the most wonderful tools we have in flies is protein traps, allowing visualization of endogenous proteins tagged with GFP. With the help of a Collagen IV functional GFP trap, we are able to visualize BMs very easily at great resolution. Collagen IV is widely regarded as an exclusive component of the planar BMs, not present in other more disordered types of matrices. However, images of the fat body, a favorite subject of study in our lab, always showed intercellular accumulations of Collagen IV that did not look as thin or continuous as BMs (Figure 2). We decided to call these novel structures CIVICs, which is short for Collagen IV Intercellular Concentrations. We then confirmed through electron microscopy that CIVICs were discontinuous, irregularly shaped and much thicker than BMs, for instance the one surrounding the fat body itself. Jose is a big fan of electron microscopy and always encourages us to include it in our projects. This is despite the fact that electron microscopy is a challenging and time-consuming technique, especially with fat body cells, which are difficult to handle because they are full of lipid droplets.
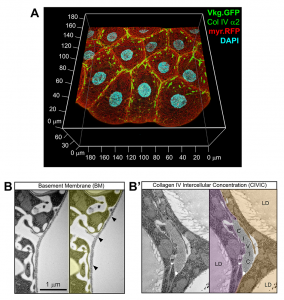
Adhesion and signaling by CIVICs
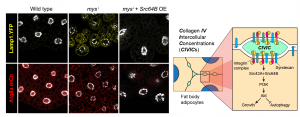
Finding these novel Collagen IV structures that looked so different from a BM in wild type flies was very exciting. The important question, though, was whether they were an accident, like fibrotic aggregates our lab had previously found [5], or they had an actual role. Knock down of Collagen IV created gaps between adipocytes in the normally continuous tissue layer, indicating that CIVICs were gluing cells together (Figure 3). At that point, we realized we had something truly interesting in our hands. Soon afterwards, we had the data showing that integrins and syndecan, another extracellular matrix receptor, were involved in the formation of CIVICs and in holding adipocytes together. This we highly anticipated, at least for integrins. However, it was a bit surprising to see that other BM components localized to CIVICs but did not have the same kind of effects as Collagen IV, especially for Laminin, which most people would place first in the hierarchy of assembly of the BM polymer. This is definitely an issue for further research for us.
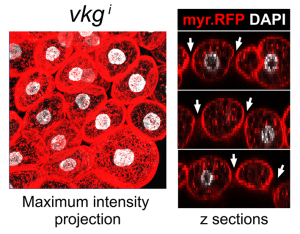
So far, we had the evidence showing CIVICs contributed to intercellular adhesion and therefore maintenance of tissue organization. The final question to address clearly was the functional consequences of this organization. Something we had noticed in our experiments knocking down Collagen IV, integrins or Syndecan was that the fat body cells did not become as large as the wild type. They did not seem to be undergoing anoikis or other kind of apoptosis, but they did not look entirely happy to us. Because others in the lab were working on vesicle trafficking in these cells, we had the tools for checking lysosome and autophagosome formation. Indeed, we saw that eliminating CIVICs triggered autophagy in these cells, which is something they normally will do if unhappy, for instance under conditions of starvation. Further experiments led us to connect CIVICs to the known PI3K/Akt anti-autophagy pathway through Src, explaining how CIVICs prevented these cells from undergoing autophagy (Figure 4). In summary, these peculiar Collagen IV intercellular structures were important for both maintenance of adipose tissue structure and physiology.
Reviewer concerns:
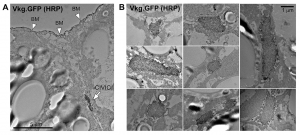
We appreciated that reviewers gave us many useful suggestions about quantifying our results and conducting appropriate statistical analysis. It was a lot of work, but we have to agree the paper is now more rigorous. Among other referees, concerns, the most difficult item to do was immune staining for electron microscopy, to prove that the structures we were characterizing actually contained Collagen IV. Since fat body cells are full of lipid droplets, they are very easy to break during the multiple fixation, permeabilization, staining and dehydration steps. I failed many times with immuno-gold staining. Others in the lab have been using APEX fusions for EM localization with amazingly good results, but there wasn’t enough time for us to create a Collagen IV-APEX transgenic line. Just at the moment I almost want to give up, I got the images with clear staining using HRP-coupled secondary antibodies and DAB reaction (Figure 5).
Final thoughts
We think our work opens new doors in the extracellular matrix and BM field, showing that the Collagen IV=BM equivalence may not be as strict as previously thought. It actually adds to work by others showing that Collagen IV-containing structures may not be all flat and homogeneously thin, blurring distinctions between BMs and more amorphous matrices [6,7]. We would definitely love to know whether structures similar to CIVICs can be seen in other Drosophila tissues or in mammals. Another distinction that is blurred by our work is that between adhesion proteins and matrix, as one could argue that Collagen IV at CIVICs is working as an intercellular adhesion molecule [8].
Finally, it is worth pointing out that the Great Wall and other defensive walls in history were, unlike The Wall in Game of Thrones, discontinuous systems of smaller walls. Actually, and despite their intended purpose, they were quite porous and ended up facilitating contacts between the peoples on both sides, enhancing commerce and cultural exchanges across them. In that sense, the Great Wall metaphor for how a system of CIVICs maintains adipocytes connected may still work.
This post is a comment on the paper: Dai, J., Ma, Mengqi., Feng, Zhi, and Pastor-Pareja, José (2017). Inter-adipocyte Adhesion and Signaling by Collagen IV Intercellular Concentrations in Drosophila. Current Biology. 18, 2729-2740.
References
- Hynes, R.O., and Naba, A. (2012). Overview of the matrisome-an inventory of extracellular matrix constituents and functions. Cold Spring Harb. Perspect. Biol. 4, a004903.
- Pastor-Pareja, J.C., and Xu, T. (2011). Shaping cells and organs in Drosophila by opposing roles of fat body-secreted Collagen IV and Perlecan. Dev. Cell 21, 245–256.
- Yurchenco, P.D. (2011). Basement membranes: cell scaffoldings and signaling platforms. Cold Spring Harb. Perspect. Biol. 3, a004911.
- Jayadev, R, and Sherwood, D.R. (2017) Morphogenesis: Shaping Tissues through Extracellular Force Gradients. Curr. Biol. 17, 850-852.
- Zang, Y., Wan, M., Liu, M., Ke, H., Ma, S., Liu, L.P., Ni, J.Q., Pastor-Pareja, J.C. (2015). Plasma membrane overgrowth causes fibrotic collagen accumulation and immune activation in Drosophila adipocytes. eLife 10.7554/eLife.07187.
- Medioni, C., and Noselli, S. (2005). Dynamics of the basement membrane in invasive epithelial clusters in Drosophila. Development 132, 3069-3077
- Isabella, A.J., and Horne-Badovinac, S. (2016). Rab10-Mediated Secretion Synergizes with Tissue Movement to Build a Polarized Basement Membrane Architecture for Organ Morphogenesis. Dev. Cell 38, 47-60.
- Zajac, A.L., Horne-Badovinac, S. (2017). Tissue Structure: A CIVICs Lesson for Adipocytes. Curr Biol. 27(18)1013-1015.
 (No Ratings Yet)
(No Ratings Yet)Get involved
Create an account or log in to post your story on the Node.
Sign up for emails
Subscribe to our mailing lists.