Now we need arrest… A long journey into the end-of-flowering
Posted by Tom Bennett, on 29 June 2020
By Tom Bennett & Catriona Walker
25th May 2020: Publication
TB: The joy of seeing an article finally published is always slightly tempered by the long-drawn out process of peer review, re-writing, re-submission, re-review, proof-reading, required to get to that point. But the publication of our article ‘Auxin export from proximal fruit drives arrest in temporally competent inflorescences’ represented the end of a much longer journey. And to be honest, even a third round of peer review didn’t diminish the joy in fulfilling a long held personal ambition. Here, we look back at some of the key moments in the making of this paper.
2003: Inspiration
TB: I don’t know when exactly I first read ‘The fate of inflorescence meristem is controlled by developing fruit in Arabidopsis‘ by Linda Hensel and the late Tony Bleecker. Even 2003 is something of a deduction. What I do remember is that it was one of a bundle of articles photocopied out of the physical journal (those were the days!), and given to me early in my PhD by Jon Booker, one of the post-docs in Ottoline Leyser’s lab. And I remember being immediately struck by the paper upon first reading it. For the uninitiated, Hensel et al (1994) is an absolute masterpiece of experimental plant science, a combination of meticulous observation, meticulous experimentation and nuanced interpretation. It is a bona fide ‘classic’.
Hensel et al focuses on an important but somewhat overlooked developmental phase, namely ‘end-of-flowering’. Flowering itself is the reproductive phase in the most abundant group of land plants, the angiosperms (or ‘flowering plants’). Flowering occurs by the production of specialised shoots (inflorescences), which generate flowers. Each flower typically has male and female organs (staemen and carpels) that produce gametes (pollen and ovules respectively). Pollination of ovules leads to the formation of seeds containing embryos, and conversion of the carpels into a fruit that holds the seeds. Given that the majority of the world’s food supply is derived from the seeds and fruits produced by flowering, it’s a pretty important process. For the plants themselves, it is critical that flowering begins at the right time of year, but also that it ends at the correct time. While we know a lot about the developmental mechanisms that control the start of flowering, we know much less about those that control end-of-flowering.
In looking at end-of-flowering, Hensel et al chose to work on what was rapidly becoming the dominant plant model species, the small, short-lived weed Arabidopsis thaliana (or just ‘Arabidopsis’). Arabidopsis reproduction occurs in a single coherent burst, elaborated through a branching inflorescence system. What Hensel et al found was that Arabidopsis flowering seemed to end by the simultaneous developmental arrest of all inflorescences on the plant, in a ‘global proliferative arrest’ (GPA) event (Figure 1). They then showed that this arrest required the presence of fertile fruit, and that removal of fruit could delay arrest, or even re-start flowering if arrest had already happened. Further, they demonstrated that it was very specifically the seed inside the fruit that really seems to drive arrest. Ultimately, they proposed a model in which the cumulative build-up of a fruit-derived signal triggers the simultaneous arrest. I thought this was a stunningly elegant mechanism for coordinating development across the whole plant body, and I knew there and then that I wanted to work on this problem, although not right there and then…this was something I was going to save for the future.

12th March 2016: Inception
TB: As it turned out, the future was a long time in coming. Originally, I’d thought about this as something to work on towards the end of my PhD, but other tasks took precedence. And then I moved away to post-doc on somewhat different areas of plant development; I never forgot about GPA, but for a long while it was mostly at the back of my mind. Luckily for me, everyone else seemed to forget about GPA for quite a while. There was no follow up from Hensel et al, and no one else picked up the baton in the meantime. Thus, when I finally picked the idea back up in earnest in late 2015, we knew little more about GPA than we had done back in 2003. At the time, I was in the process of applying for academic positions, and GPA was clearly something that I was going to work on when I started my own group. In February 2016, I accepted a position at the University of Leeds to start later that year, and shortly thereafter, I set up my first GPA experiment (the imaginatively titled ‘GPA1’). When I started, I never really doubted that the phenomenon was already defined, and what I was looking for was just the mechanism. But GPA1 didn’t quite go as expected; yes, the plants arrested as expected, but I kept watering the plants just to see what would happen. To my surprise, the plants initiated new inflorescences and started flowering again (Figure 2). Clearly, this wasn’t going to be quite so straightforward as I’d thought…

20th January 2017: Proclamation
TB: As the ’new boy’ at Leeds, I was asked to give the ‘keynote’ talk at our annual Centre for Plant Sciences symposium, shortly after I started there. I’d never talked about my plans to work on GPA before, but I decided that’s what I was going to talk about, I think mainly because I was excited by the prospect of getting ‘stuck in’ to the project. I had very little actual data to present, but I pitched the talk on the importance of the phenomenon, and talked mostly about the theoretical background. I wasn’t sure how well it would go down, but as it turned out, it was something of a masterstroke…
CW: I had no idea who Tom was at the start of the symposium (other than the tall man who I occasionally passed in the corridors at work), but by the end of his talk, I had basically decided that I also wanted to contribute to understanding GPA. Tom had outlined some of what he planned to do, but I was particularly interested in the fact that he pointed out it could be relevant in crop species such as oilseed rape – I had an agricultural background, and wanted to do more work with crops. At this stage I had no interest in working with Arabidopsis, and figured I could provide some crop knowledge and avoid Arabidopsis. I wasn’t working for Tom at the time, so I figured any research I did contribute, I would just do in oilseed. Funnily enough, that’s not how things ended up working out.
5th June 2017: Recruitment
CW: Not only did I end up working for Tom, I also ended up spending the majority of my time working on Arabidopsis. With hindsight, my naivety about Arabidopsis was key to me completely buying into the end of flowering being a fascinating process. Very early on, I was carrying out an experiment where I was going to treat plants at floral arrest. During a routine check of the plants, Tom asked why I hadn’t carried out the treatments yet. I was confused by this – clearly the plants hadn’t finished flowering. Tom indicated that the primary inflorescences had all arrested – I countered that many of the lower inflorescences were still vigorously flowering. At this stage, we began really questioning GPA as a concept. If floral arrest was synchronous, how could these plants have some inflorescences that had arrested, and others which were still flowering several days later? The next step was to closely examine the flowering lifetimes of every individual inflorescence of a set of plants, where we clearly showed that floral arrest isn’t synchronous, it follows a basipetal wave, with the upmost inflorescences on the plant arresting first, followed by each inflorescence below it in turn. Each inflorescence class (primary, secondary, tertiary etc) also displayed a characteristic lifetime, where the timing of arrest appeared to be controlled specifically by the time since that inflorescence had initiated. I was hooked on the project from this point on; I needed to know how this was controlled!
1st November 2017: Nottingham
TB: Back in May 2016, a paper on GPA had been published in Plant Physiology, suddenly breaking the radio silence on the subject after 22 years (and giving me that terrible sinking feeling, until I realised that their approach was very different to what I was planning). So I was very much aware that interest in the area was starting to emerge again, but I had very little idea who else might be working on GPA. As it would turn out, at least two other research groups had also recently developed interests in GPA – Cristina Ferrandiz’s group in Valencia, and Zoe Wilson’s group in Nottingham.
It was therefore very serendipitous that my first seminar as a group leader on the ‘academic circuit’ was in Nottingham. I think it came as a surprise to Al Ware (the PhD student working on the project) and Zoe that there was someone else working on GPA, just as it came as surprise to me that my audience was really interested in what I had to say on that subject! Happily, we sat down to talk some more GPA after my seminar, and it rapidly became clear that our data and approaches were completely complementary. We agreed to collaborate there and then, and over the next few months, began to sketch out a plan for a manuscript that combined our data. Collaborating with Al and Zoe has been thoroughly enjoyable, and of massive benefit to both groups, so it really was a genuine stroke of luck to meet them so early in the project!
June 2018: Broken
CW: This was an ‘interesting’ period in the development of the GPA research. In each experiment we set up, we were convinced that – this time – we knew what the outcome would be, but each time the results were completely different to what we had expected (Figure 3) and led to multiple new hypotheses and experiments.
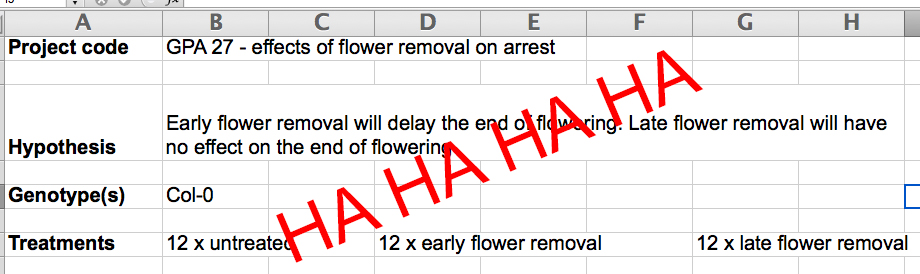
Ultimately, this resulted in GPA 27 (we’d come a long way from GPA 3 when I started a year ago) – simultaneously one of my favourite and most-hated experiments to date. The plan was to remove all open flowers from plants daily; I think at this stage we were expecting the plants to arrest at a ‘normal’ time – we were wrong. Not only was floral arrest delayed, but with no fruit production, the plants kept producing more branches, meaning flower production was becoming exponential. What had started out as half an hour each morning removing flowers became the majority of every day pulling flowers off increasingly more gnarled, brittle Arabidopsis (Figure 4). It wasn’t quite as fun as it sounds. I was doggedly insistent that I was going to see this experiment to a close, but was away at a conference for three days, so asked Tom if he’d look after the plants while I was gone – it was only a few days, what could possibly go wrong?

By the second day, Tom had emailed me to tell me that it was probably a good idea to halve the number of plants we were treating; we didn’t need as many as we had, and he’d leave the rest as a ‘recovery’ treatment (traitor). Funnily enough, my horror at this suggestion wasn’t enough to change Tom’s mind, and I returned to the lab to find that the ‘recovery’ plants had produced fruit and…arrested. The treated plants were still flowering on, but the recovery plants had happily arrested and stopped flowering, despite only producing a few fruits per inflorescence. I began to forgive Tom for the ‘recovery’ treatment, and we started questioning how many fruit were actually needed for arrest. Hensel et al. had suggested around 40% seed set was required for arrest, but hadn’t specified whether this was per fruit, per inflorescence… We’d removed hundreds of flowers from these plants, so clearly they hadn’t produced that much seed in the few days of the recovery treatment. We began to question whether it was the position of the fruit, rather than the number, that was crucial to arrest.
After GPA 27 finally finished, I ‘decided’ to take a few weeks of holiday to recover, and swore never to repeat another flower removal experiment. Obviously, when I came back, I’d forgotten the trauma of GPA 27, and carried out another flower removal experiment, only this time we focussed on individual inflorescences. Only a few inflorescences per plant had flowers removed, while the rest of the plant remained untreated. Pleasingly, I found that only the treated inflorescences had a delayed arrest – the rest of the plant arrested when we would expect. Not only had we shown that floral arrest wasn’t synchronous, now we’d also shown that it wasn’t global – it was local, and it was dependent on a small number of fruit.
7th February 2019: Submission
TB: By the end of 2018, we’d managed to collect all the data we needed for the planned manuscript, and we spent the first month of 2019 putting the manuscript together. We’d always planned to submit to Nature Plants; I’d been given some encouragement by one of the editors (Guillaume Tena) at a conference the previous summer, and I felt like our story was of sufficient general interest. Nevertheless, it’s always a tense wait to hear if the journal is going to send the manuscript for review. And often you don’t actually here anything from the journal at all until the reviews land in your inbox. Fortunately, Guillaume let me know fairly quickly that the manuscript was going out for review, making the whole process that bit less stressful!
Summer 2019: Redux
TB: The reviews — all four of them — came back at the start of April. Two of them were very positive, while the other two were positive, but had some serious reservations, particularly with respect to our divergences from the GPA model. But I have to say that, the comments were all very fair, and ultimately very helpful. Overall, this was by far the best experience I’ve had of peer review; I really feel like the reviewers helped us make a better and more coherent paper. We got straight to work on trying to provide new data to address the comments.
CW: Following the reviewers’ comments, we set up several key experiments to really focus in on the numeric and positional fruit requirement for arrest. My primary focus for the paper at this stage involved removing fruits at different timings prior to arrest, and allowing recovery periods to see under what conditions arrest occurred. Through this, we showed that only a small number of fruit are required to bring about arrest – and that the closer they are to the meristem, the faster the arrest; whereas a large number of fruit far from the meristem will not bring about arrest at all.
Interestingly, I accidentally further refined the number of fruit required for arrest while working on a totally different area of my PhD. I was looking at whether removing increasing numbers of flowers along an inflorescence (every other flower, 2 out of every 3, etc.), increased the final size of fruits. It didn’t. But nor did it really affect arrest either. If I removed 4 in every 5 flowers, the inflorescences arrested only a couple of days later than untreated inflorescences, despite only having 20% of the fruit. I particularly love that this experiment, which had a completely different purpose, ended up helping us to answer a different question we were working on. It’s so satisfying to see different areas and ideas link up in unexpected ways, but I hadn’t really experienced that in my own research before this time.
4th April 2020: Acceptance
TB: We finished re-writing the paper just in time for Christmas, and re-submitted on 17th December. I was pretty confident we’d addressed the reviewers comments, but after a second round of peer review (17th February), there were one or two new issues to address, though it was pretty clear publication was going to happen, as long as we addressed those issues. Fortunately, we already had the data, so we resubmitted within a week, and went back out for a final round of review. Then, finally, on 4th April, we received the good news…
CW: Looking back over the lifetime of this project, it’s incredibly satisfying to see how things have progressed so much – not just the research, which I’ve loved, but also generally how our approach and understanding developed as a whole. The first summer I was in the lab was very much a case of trying to get to grips with a new concept and species. The second summer was incredibly productive, but also very draining, both physically and mentally. We got great data out of that time, but it did come at a cost (although I did learn that 30+ consecutive days in the lab really is my limit). By the summer of 2019, we really knew what we were doing. Granted, we still found answers in accidental ways, but our whole approach was much smoother and more refined. Having the manuscript accepted really provides a satisfying point of perspective over how things have developed in the last few years. Now we’re on to the next exciting thing, and I can’t wait to find the answer. It won’t even involve any flower removal. Probably.
Auxin export from proximal fruits drives arrest in temporally competent inflorescences
Alexander Ware, Catriona H. Walker, Jan Šimura, Pablo González-Suárez, Karin Ljung, Anthony Bishopp, Zoe A. Wilson & Tom Bennett
Nature Plants 6, 699–707(2020)
 (1 votes)
(1 votes)Get involved
Create an account or log in to post your story on the Node.
Sign up for emails
Subscribe to our mailing lists.