The BSDB had its joint Spring Meeting with the BSCB earlier this year in April (10th -13th). It was my first time at the Meeting, which had been enthusiastically sold to me as “a good one, a fun one – they have a pub quiz!” and although I had been ‘sent to Coventry’, I have to confess, I had a great time. It was a busy 3 days, brimming over with interesting talks and stellar plenary and medal lectures. The concurrent sessions were on topics of interest for both cell and developmental biologists and so I ended up flitting between the talks of the concurrent sessions each day. Unfortunately, due to the impossibility of being in two places at once, there were excellent talks that I did miss and I apologise to these speakers for my non-attendance. I also want to say a big thank you to the speakers who have very kindly given me permission to write about the unpublished work they presented.
The meeting started, for me, with a Career Workshop on the Sunday afternoon. It was organised as a series of roundtable discussions with lecturers, group leaders, and people in industry, science publishing and communication. I got to speak to Paul Conduit, a Henry Dale Fellow at the University of Cambridge, Anne Wiblin from Abcam who provided the perspective of science life in industry, and Catarina Vicente from the Node, with whom I discussed the power of Twitter (and cakes) for communicating science.
Sunday evening’s plenary lectures were given by stalwarts – Mark Kirschner gave the BSDB Plenary Lecture and Ruth Lehmann the BSCB Plenary Lecture.
Mark Kirschner’s lecture sought to give quantitative answers to a fundamental question – “what is the economy of RNA and protein in embryonic development?’ with impressive technology and precision. By examining the dynamics of RNA and proteins on a single cell level, the Kirschner group has been able to show that while the correlation between the mRNA and protein levels is 0, the protein synthesis rate correlates with the mRNA synthesis and decay rates (Peshkin et al. 2015).
Jordan Raff, while introducing Ruth Lehmann, mentioned that he remembered her work, where injecting nanos mRNA into the anterior of the embryo caused the formation of an embryo with “essentially 2 bums”. Co-incidentally Ruth Lehmann came back to the “2 bums” embryo in her talk in the context of mitochondrial segregation in primordial germ cells. A second research story she presented was of mitochondrial maturation during germline cyst formation – developmentally regulated folding of the inner mitochondrial membrane into cristae is dependent on ATP-synthase dimer formation (Teixeira et al. 2015).
In a break with tradition, the student and post-doc social did not have a pub quiz, but instead we were put into random pairs and teams to build spaghetti towers and play science Pictionary. It was an excellent way to meet fellow attendees and bond over our total lack of talent at either drawing or building design (or complete mastery of, in a few cases).
Monday was a day packed with scientific goodness – 4 sessions of talks, 2 medal lectures and 2 poster sessions! The sessions of the morning were Cell and Tissue Architecture and Evolution. In the Cell and Tissue Architecture session I listened to Val Wilson talk about neuromesodermal progenitors (NMPs) – bi-fated cells at the caudal end of the mouse embryo who contribute either to the spinal cord or paraxial mesoderm, depending on their position in the progenitor region. She talked about NMP behaviour during the formation of the mid-trunk (Wymeersch et al. 2016). Olivier Hamant went on to talk about the role mechanical forces play in providing growth cues – microtubule dynamics in response to stress give the characteristic shape of sepals in Arabidopsis (Hervieux et al. 2016). Yara Sanchez Corrales showed how tube morphogenesis in the embryonic Drosophila salivary gland occurs. Cell division does not occur during the process and tube formation is driven entirely by cell shape changes and rearrangements. Their live cell imaging approach allows for the cell shape dynamics to be analysed in 3D.
In the Evolution session, I took in the talks of Marie-Ann Felix, Andrew Gillis and Marty Cohn. Marie-Ann Felix presented work aimed at understanding the effect of random mutation at the phenotypic level. They assayed the sensitivity of the vulval precursor cells of C. elegans to fate changes after the accumulation of random mutation and find that the P3.p vulval cell fate is most affected and is sensitive to mutations in many loci. This means that mutational effects can produce altered responsiveness to signalling pathways and, in this view, the P3.p cell fate is evolving the fastest. Andrew Gillis showed that Sonic hedgehog is required for gill arch anteroposterior polarity and for the branchial ray development in the gills of sharks, skates and rays (Chondrichthyans). This finding parallels paired fin development, giving traction to the hypothesis that there is serial homology between paired fins (and tetrapod limbs) and Chondrichthyan gill arches. Marty Cohn talked of the origin of cartilage – while true cartilage is considered to be unique to vertebrates, invertebrates like cuttlefish show cartilage-like tissue that develops via a deeply conserved gene regulatory network (Tarazona et al. 2016).
The afternoon sessions were Hijacking Cell and Developmental Processes and Polarity. In the Hijacking session, Shuchen Zhang presented her work trying to understand the genetic interactions of Sox2 that underpin its dual functions as a regulator of pluripotency in human embryonic stem cells and as a key factor in neural differentiation. Steve Jackson presented work on how cell-based screens in his lab have identified novel drug targets for cancer therapies, focussing on a PARP inhibitor that is now used in chemotherapy to treat hereditary ovarian cancers. In the Polarity session, Takashi Hiiragi showed that the apical domain of cells is instructive for the initial symmetry breaking in early mouse embryonic development by some fantastic live imaging. Nate Goehring showed us how through the use of novel small molecules, his lab has manipulated the activity and localisation of kinase PKC to understand the PAR polarity network in the C. elegans embryo. Ray Keller talked of the “mechanome” and the game plans – convergent extension, epithelial to mesenchymal transition and convergent thickening (driven by changes in cell affinity) – that tissues use to generate the forces required to drive morphogenesis in amphibians (Pfister et al. 2016).
The evening was host to BSCB’s Hooke Medal and BSDB’s Waddington Medal lectures. Tom Surrey was the recipient of this year’s Hooke Medal. He presented 3 facets of his lab’s research into understanding microtubule dynamics. Through impressive time-lapse fluorescence microscopy movies, we were shown the dynamics of microtubule growth, catastrophe (Duellberg et al. 2016) and nucleation (Roostalu et al. 2015) in in vitro reconstitutions. Tom Surrey’s medal lecture (and all other medal lectures of the meeting) can be watched here:
The recipient of the Waddington Medal was completely shrouded in secrecy until just minutes before the lecture, Ottoline Leyser (the BSDB president) even engaging the audience in a little guessing game to introduce him. It was Enrico Coen! He started his talk with a series of sketches of bulls by Picasso and posed the question – what is harder, describing something with every detail or capturing its essence? In a very cinematic presentation with beautiful images and movies, videos of collaborations with potter and glass-blower friends set to a lush soundtrack and a live demo of growth conflicts with melting plastic, Enrico Coen gave us the essence of his obsession – the snapdragon flower. The talk strongly resonated with the conference. References were made to it in almost every subsequent session and he might just have convinced everyone to drop everything they are doing and start studying snapdragons and bladderworts. I have it on good authority that he believes a good talk is like the movie High Noon. I know what you’ll be doing at your next lab meeting…
On Tuesday morning I attended the Growth and Cell division session. I heard Anja Geitmann talk about the mechanisms the pollen tube uses to grow towards the ovaries in plants and it genuinely seems a bit of a wrecking ball. Using a microfluidic device her group has been able to measure the pressure the pollen tube can exert – it is about 150kPa. That’s the pressure in a car tyre! Shane Herbert presented data showing that asymmetric cell divisions and Notch-Delta mediated lateral inhibition establishes the hierarchy of motility in the endothelial tip cells that allows for the leader-follower mode of cell migration in angiogenesis and Silvia Santos brought the session to a close by shedding some light on the temporal control of the cell cycle. Mitosis time is typically short and constant and insulated from the timing of early cell cycle phases. This seems to be regulated by the positive feedback of Cdk phosphorylation. Breaking the feedback leads to longer, more variable mitosis that is coupled to the interphase time.
The Graduate Student Symposium was held on Tuesday afternoon. Despite being fraught with technical difficulties, it was one of the most enjoyable and engaging sessions of the meeting. Kudos to the speakers for their quick thinking and improvisation in face of the ‘your-presentation-won’t-play’ challenge! We heard about (and saw, when the computers co-operated) Mycobacterium infection, cell divisions in motile cells and stretched tissues, centrosome clustering in cancers, neuroretinal self-organising aggregates, neural tube lumen formation, neuromesodermal progenitors, segmentation in spiders and flies, zebrafish cilia formation and calcium signalling in angiogenesis.
Uri Alon provided a brief and unexpected (i.e. not in the schedule) interlude titled the ‘The Life Scientific’ before the Woman in Science Medal talk. With a flip chart, a guitar, witty and tuneful songs that required back up singing by the audience, Uri Alon highlighted the need to acknowledge and discuss the emotional and subjective nature of the scientific process. Feeling lost, stuck and being in “the cloud” is all part of doing innovative science. Please, please watch his TED talk on YouTube, whether you are lost in “the cloud” or not. It is Game of Thrones-level essential viewing.
The Woman in Science Medal was awarded to Lidia Vasilieva for her work in understanding mechanisms of gene expression. Lidia began by highlighting the progress made in science to help women achieve their goals and then focussed on her work in understanding the regulation of gene expression. Her lab has discovered that exosome mediated RNA degradation, in co-operation with the splicing machinery, can regulate levels of mRNA (Kilchert 2015).
Abigail Tucker was the recipient of the first Cheryll Tickle medal. The Cheryll Tickle medal is being awarded by the BSDB to a mid-career female scientist for outstanding contributions to her field. And Cheryll Tickle herself was present to award the medal to Abigail Tucker. She quipped that she was rather glad that the BSDB went with her full name for the medal as “the Tickle medal” might suggest an award for something else (but the medal does feature a feather (!)). Abigail Tucker’s talk was a simultaneous career and life retrospective. She presented a career timeline, talking briefly about her PhD and postdoc work before taking us through the current activities of her lab studying the development of opossums, pit vipers and cobras. I imagine that this is the kind of work Indiana Jones would do, if he were a scientist. She also highlighted personal events of great significance on her career timeline. It is both inspiring and heartening to know that it is possible to have a thriving scientific career alongside a family.
Wednesday morning was a bit of tough start thanks to the late-night/early-morning revelry and dance floor antics that followed the conference dinner. But, I did manage to make it for Wendy Bickmore’s opener for the information processing session at 9.30am (!) about enhancer-promoter interactions studied by chromatin conformation capture and single molecule FISH. She was followed by Stefano De Renzis who showed how he can reconstitute invagination in embryonic Drosophila tissues that normally wouldn’t by optogenetically modulating the local actomyosin contractility.
I then caught the last 2 talks of the ageing and regeneration session. Yves Barral showed us how the bud lineage in budding yeast stays immortal – a diffusion barrier made of phytoceramides separates the mother and daughter preventing exchanging of membrane proteins and allows age to accumulate in the mother cell. Allison Bardin talked about mechanisms causing instability of the genome in adult stem Drosophila intestinal stem cells and how this affects homeostasis. Using an X-linked Gal80 construct she showed that mitotic recombination promotes loss of heterozygosity. She also showed that genomic rearrangements in these cells could lead to the spontaneous development of neoplasias in male flies (Siudeja et al. 2015).
Elena Scarpa, recipient of the Beddington medal for the best PhD thesis, presented her work in Roberto Mayor’s lab before Uri Alon brought the conference to a close. Elena showed the role of cadherins and the interplay of intracellular and external forces in contact inhibition of locomotion in migrating neural crest cells (Scarpa et al. 2015). Uri Alon spoke about how quantitative thinking could be brought into discussions of morphology and showed work from his lab where phenotypes had been studied using Pareto optimality. The approach is based on the logic that no phenotype can be good at all tasks and there is a trade-off with respect to the tasks to ensure maximal fitness, leading to optimal phenotypes. These phenotypes fall into simple shapes such as lines and triangles (Pareto fronts), the vertices representing an archetype – phenotypes that are specialised at a single task (Hart et al. 2015).
To sum up my experience at Warwick – I had spent 3 days being inspired by great talks, fascinated by all the new science I had heard, making new friends, having interesting discussions about my project and science in general. I discovered I am as hopeless at art as I am at origami and have no future at all as an architect. I was also strangely buoyed by the knowledge that most of my experiments are destined to fail. I returned home and fell into the dreamless slumber of a happy and exhausted child and woke up refreshed and ready to get lost in “the cloud”.
The laboratory of Developmental Genetics is looking for a PhD student to study the molecular mechanisms of neurogenesis in the developing mouse nervous system. The student will have to apply for a FNRS FRIA fellowship (http://www.fnrs.be/index.php/news-fnrs/517-fria-fresh-2016). The deadline for the application is the 30th August 2016.
The unit is located in the Biopark Charleroi Brussels South (http://ibmm.biopark-it.be/bced/), about 25 miles south of Brussels, in the Institute of Molecular Biology and Medecine (IBMM), a leading multidiciplinarity Institute from the faculty of Science and Medecine of the ULB (http://www.ulb.ac.be/rech/inventaire/unites/ULB578.html). The unit is part of the ULB Neuroscience Institute (http://uni.ulb.ac.be/groups/developmental-genetics/). The group is studying the molecular mechanisms that control the transition from neural stem cell to neurons in the developing vertebrate nervous system. The focus is on the role of some transcription factors in the molecular mechanisms that control neural progenitor maintenance, differentiation, and the generation of neuronal diversity. Major ongoing researches focus on the role of Dmrt transcription factors in cerebral cortex development and of Prdm transcription factors in pain perception, in health and diseases.
The laboratory uses in vivo genetic approaches in the mouse as well as gain- and loss-of-function experiments in the frog to approach gene function in the developing embryo.
The candidate will be involved in one of the two following projects:
Project A:
Pain perception has evolved as a warning mechanism to alert organisms to tissue damage and dangerous environments, and is therefore essential for survival. In human, erroneous activation of the pain-sensing system, as in chronic and neuropathic pain, represents a major health burden with insufficient treatment option. New therapeutic options have recently been developed from studies of a small number of individuals with Congenital Insensitivity to Pain (CIP). The majority of these have Mendelelian disorders of painlessness, where disruptive mutations in a single gene are responsible for their inability to sense pain.
Prdm12 has recently been identified as mutated in individuals with CIP (Chen et al., Nature Genet 47, 803-808, 2015). In our laboratory, we have obtained evidence that Prdm12 is crucial for the generation of the nociceptors, the type of neurons that sense noxious stimuli and transfer nociceptive information to the CNS (Nagy et al., Cell Cycle 14, 1799-1808, 2015). Prdm12 belongs to a family of evolutionarily conserved epigenetic regulators that control neuronal specification (Thélie et al., Development 142, 3416-3428, 2015). It is highly and selectively expressed in differentiating nociceptors and remains expressed in these cells post-natally, suggesting that modulating it may be a new route for pain control.
In this project, our aim is to elucidate Prdm12 mechanism of action in nociceptor differentiation during mammalian embryonic development and determine whether its activity also influences nociceptive function in the adulthood. These objectives will be approached through the detailed characterization of Prdm12 null knock-out and transgenic mice and cell lines available or under construction and using state-of-the art genomics, epigenetic, proteomic and electrophysiological approaches.
Projet B:
Understanding the mechanisms that control the generation of distinct types of neurons from multipotent progenitors constitute a major challenge in developmental neurosciences. Transcription factors are at the core of the programs that control cortical development. Two members of the Dmrt family of zinc finger transcription factors, Dmrt3-5, are expressed by cortical progenitors in a similar high caudomedial to low rostrolateral gradient. Our laboratory has shown that Dmrt5, whose mutation in human has been recently associated with microcephaly (Urquhart et al., Clinical Genetics, 2016) is essential for the development of the caudomedial part of the cerebral cortex including the hippocampus and that it plays a direct role in neocortical progenitors in the control of their specification (Saulnier et al., Cereb. Cortex 23, 2552-2567, 2013 ; De Clercq et al., submitted). Recent results of the laboratory indicate that Dmrt3 also contribute to cortical patterning. Despite their importance, the mode of action of Dmrt5 and Dmrt3 in the specification of cortical progenitor identity and in the control of cortical growth remains largely unknown.
In this project, our objective is to understand these mechanisms through the characterization of the phenotype of Dmrt5-/-;Dmrt3-/- double mutant mice and the identification of their in vivo genomic binding sites (using chromatin immunoprecipitation). These studies should provide important insights into the transcriptional mechanisms controling early cortical development.
PhD candidates should be highly motivated and have previous experience in mouse handling, neurobiology or molecular biology. Interested candidates should send their CV including a motivation letter and contact information of at least two previous supervisors able to recommend their research ability to ebellefr@ulb.ac.be
For more information, see our recent work:
Saulnier et al. (2013). The Doublesex Homolog Dmrt5 is Required for the Development of the Caudomedial Cerebral Cortex in Mammals. Cerebral Cortex, 23, 2552-2567.
Nagy V, Cole T, Van Campenhout C, Khoung TM, Leung C, Vermeiren S, Novatchkova M, Wenzel D, Cikes D, Polyansky AA, Kozieradzki I, Meixner A, Bellefroid EJ, Neely GG, Penninger JM. The evolutionarily conserved transcription factor (2015). PRDM12 controls sensory neuron development and pain perception. Cell Cycle. 14, 1799-808.
Thélie et al. (2015). Prdm12 specifies V1 interneurons through cross-repressive interactions with Dbx1 and Nkx6 genes in Xenopus. Development 142, 3416-3428.
Mark Hintze and Diana Gradinaru introduce their collaborative animation about the wonders of developmental biology.
How are you built? How do you become the shape and form that you are? How are your arms the same length? These questions and many others lead me to study for a PhD in developmental biology. Yet, during my PhD I was often explaining the fundamental ideas about developmental biology to my parents and friends. Having a huge passion for the wonder of the developing embryo, I thought it would be great if this science was more accessible to everyone.
YouTube videos from channels including minutephysics, veritasium and smarter every day do an amazing job of explaining complex phenomenon in an easy and fun format. I thought a similar format would be a great way to showcase developmental biology and how it attempts to answer the question of how are you formed.
Kings College London runs a scheme providing a grant for a cultural collaboration with an artist to fuse science and art. I applied to this scheme and won a small grant to collaborate with an artist to create an animated video. This scheme put me in collaboration with the brilliant artist and animator Diana Gradinaru. Her amazing creativity helped push the idea from my head into the wonderful animation that you see in the video. Below the video, she tells us how she approached and applied the creative process to animating Life’s Symphony. MH
I remember being a pupil at school loving biology. We had the most wonderful collection of hand-painted large-size illustrations of plants and animals (all handsomely dissected on the page). This was in 90s post-communist Romania when illustration was a craft honed by many working-class men and women trying to do their bit for schools. There was so much clarity in the way these artists had represented the simplified world of science for the benefit of our young minds, but they equally reminded me of Victorian studies I had stumbled upon in art history books.
There was a certain effectiveness to the interactivity between the teacher coming to the front of the class explaining these wonders to us; using these studies, the microscope and the ability to render a perfect depiction of the digestive system on the black board.
I was thrilled when Mark proposed a collaboration revolving around his practice. Animation lends itself effectively to science. It is a time-based medium which can implement a range of intriguing drawing styles and can represent abstract concepts with humour and panache. It can draw the viewer in by alternating the delivery of information with moments of ease and laughter. As I advanced through the ranks at school, the science books we had become more and more rigid and bare. If only science was rendered with more wit and charm, then people would feel less daunted by it and understand that science, far from being an enclosed subject with a clear-cut trajectory, is actually a bewildering realm, constantly evolving and permeating all aspects of our life.
The animation illustrates a journey through developmental biology. Showcasing how species have evolved to use many of the same genes in different ways and at different times to create the shapes and patterns we see in the world before us. We hope that it displays how the formation of the embryo and adult form is much like a symphony. Where many parts must play together at the right moments to create a concordance and harmony allowing development to occur. DG
Fin to limb transition was a crucial evolutionary adaptation necessary for the vertebrate land invasion in the late Devonian around 400 MY ago. Without this key event, I would not be writing this blog, we as a species would definitely not exist, and neither would any other tetrapod. While the gross morphological changes for this process are fairly well understood, the cellular and molecular mechanism controlling these changes remained a total mystery. In our recent publication (Masselink et al, 2016) we identified a unique and novel lineage of cells within the pectoral fins that have been gradually lost throughout tetrapod evolution and regulate the fin to limb transition by modulating FGF8 and FGF24 expression in the Apical Ectodermal Ridge (AER) and the underlying fin bud mesenchyme.
Comparative embryological studies by Thorogood and others suggested that a gradual delay and eventual complete disappearance of the AER to Apical Fold (AF) transition was a driving force of evolutionary fin to limb transition (figure 1, Thorogood, 1991). More recent work by Yano experimentally confirms Thorogood’s hypothesis (Yano, 2012). The Apical Ectodermal Ridge, an ectoderm derived structure that covers the underlying mesenchyme of both the fin and limb bud, secretes pro-proliferative factors such as FGF8 to provide cues for distal outgrowth of the underlying mesenchyme. While in tetrapods the AER perdures and eventually undergoes apoptosis, in fish the AER transitions into the AF, which eventually gives rise to the fin paddle and are populated by actinotrichia, non-ossifying collagen ridge fibrils which provide rigidity to the embryonic fin paddle. Thus a gradual delay and eventual complete disappearance of AER to AF transition would not only allow for the disappearance of the fin paddle but the prolonged presence of the AER would also allow for increased distal outgrowth of the underlying mesenchyme.
Thorogood’s clock model suggest that the gradual delay and eventual complete disappearance of AER to AF transition during embryonic development is an important step in fin to limb transition. (Modified from Thorogood, 1991)
We identified a novel lineage of fish-specific AER infiltrating cells that control AER to AF transition that also secrete the collagens required for actinotrichia formation (figure 2). We have thus termed these cells Apical Fold Inducing Cells (AFICs). Using 3 different methods (confocal time lapse microscopy, photoconversions, and somite transplantation) we conclusively show these cells have a somitic origin, a lineage which despite decades of lineage tracing experiments was never identified in tetrapods, suggesting that this is indeed a process unique to fish. Metronidazole and laser mediated ablation were used to selectively ablate AFICs, resulting in fins to form with certain limb-like characteristics. Specifically we observed phenotypes consisting of an expanded mesenchyme, a disruption in AF formation, and a loss of actinotrichia, we also observed an increased expression of FGF8 in the AER and a delay of FGF24 translocation from the mesenchyme to the AER. Inhibition of FGF signalling by treatment with the FGF inhibitor SU5402 completely blocks the increased proliferative response in the mesenchyme, and thus provides a molecular basis for the fin to limb transition.
AFIC infiltration into the AER controls fin fold induction. AFICs have a shared somitic origin with the muscle progenitor pool (green) and migrate into the AER (yellow). Upon infiltration into the AER, the AER folds onto itself and forms the AF (red). This infiltration event is absent in tetrapods such as mouse and chicken, which have an AER until much later in development until it apoptosis away. The continued exposure to AER derived pro-proliferative signals, allow for the expansion of endoskeletal and myogenic tissue.
The induction of the fin fold and actinotrichia formation are two distinct processes, both controlled by AFICs. We have shown AFICs to consistently localize to the distal edge of the actinotrichia, and secrete actinotrichia-associated collagens. The loss of actinotrichia -by knockdown of Actinoidin 1 and 2- results in a fin fold defect but not an expansion of the underlying mesenchyme (Zhang, 2010). Thus while AFICs are essential for actinotrichia formation they play a second role in blocking the expansion of the fin bud mesenchyme, likely through fin fold induction and the subsequent modulation of FGF8 and FGF24.
To interrogate the function of AFICs in key phylogenetic representatives of the jawed vertebrate (gnathostome) lineage, we took advantage of the Collagen1a1a antibody that detects zebrafish AFICs. The extant gnathostome lineage is broadly made up of sharks, ray-finned fish and lobe-finned fish. Species within each group were selected based on availability. An active breeding facility of the small benthic Epaulette shark was established providing access to embryonic tissue, while frozen tissue sections of embryos from both the American paddlefish (basal ray-finned species) and the Australian lungfish (lobe-finned species) were fortunately available. We found that both the timing of residency and number of AFICs are gradually reduced throughout evolutionary representatives. While the epaulette shark contains a large number of AFICs in the AER at an early stage of embryonic development, this is already delayed in timing and reduced in number in the American paddlefish. Most strikingly the Australian lungfish, a species that abuts the tetrapod lineage contains very few AFICs, which infiltrate at a late stage, and are lost all together at later stages of development. The total number of AFICs and their timing of residence is in line with the phylogenetic sampling these species present and support Thorogood’s clock model.
While our results reinforce the validity of Thorogood’s clock model, and provide a cellular framework for the changes that had to occur during fin to limb transition, they open many new questions. What controls AFIC infiltration, and what controls its timing? At this point we can only speculate. AFICs migrate in a pool of muscle progenitors into the embryonic fin bud. Here they migrate through the basement membrane into the AER. While active migration through the basement membrane is very likely to be important, this does not in and of itself explain the temporal differences observed between species. Muscle progenitors are spread throughout the zebrafish fin bud mesenchyme, coming into direct contact with the basement membrane that separates the AER from the underlying mesenchyme. In contrast, in both chicken and mice the muscle progenitors in early limb buds are restricted to the proximal region of the mesenchyme and do not come into direct contact with the basement membrane. As such, it is possible to imagine that the time between fin/limb field establishment, and muscle delamination and subsequent migration might be essential.
References:
Masselink, W. et al. A somitic contribution to the apical ectodermal ridge is essential for fin formation. Nature doi:10.1038/nature18953(2016)
Thorogood, P. in Developmental Patterning of the Vertebrate Limb (eds Hinchliffe, J. R. et al.) 347-354 (Springer US, 1991).
Yano, T., Abe, G., Yokoyama, H., Kawakami, K. & Tamura, K. Mechanism of pectoral fin outgrowth in zebrafish development. Development139, 2916-2925 (2012).
Zhang, J. et al. Loss of fish actinotrichia proteins and the fin-to-limb transition. Nature466, 234-237 (2010).
Here we highlight some developmental biology related content from other journals published by The Company of Biologists.
Aging zebrafish an ideal model
Mice have been the traditional model of choice
for investigating telomere shortening in aging, but zebrafish provide an upcoming complementary system with well-conserved physiology. Ferreira and colleagues discuss how fishes have helped our understanding of how telomere attrition contributes to cellular senescence, organ dysfunction and disease. [OA]
Diverse phenotypes in neurofibromatosis models
Neurofibromatosis type 1 is characterised by nerve sheath tumours but also has significant clinical heterogeneity. Robert Kesterson and colleagues show that two mouse models carrying NF1 patient-specific mutations have distinct effects on embryonic development, neurofibromin function and plexiform neurofibroma formation, with implications for therapeutic studies. [OA]
Maternal effects of chromosomal abnormalities
Early emerging problems during oogenesis, such as DNA double-strand breaks, can affect chromosome duplication and segregation in embryogenesis in Drosophila, Andreas Kegel and
colleagues report. Moreover, environmental cues including temperature are important for proper oogenesis. [OA]
Antibodies light up membrane traffic
Sean Munro and
colleagues report the generation and
characterisation of a toolkit of
monoclonal and polyclonal antibodies for labelling the major compartments of the secretory and endocytic pathways in flies, and demonstrate their applicability in multiple cell types. [OA]
Sperm centrosomes defines spindle assembly
Isabelle Vernos and colleagues use a Xenopus egg extract system to analyse how the first embryonic spindle is formed, and find that the duplicated sperm centrosomes define the kinetics of spindle bipolarization, ensuring their own inheritance to the daughter cells.
Probing chromatin in living cells
Gladys Mirey and colleagues present Chromatibody, a
genetically encoded single-domain antibody that binds in a non-invasive manner to the heterodimer of H2A-H2B histones across eukaryotes. This tool allows real time chromatin imaging across eukaryotes, as well as a means to target proteins to chromatin. [OA]
An overview of Myosin-I motors
In their Cell Science at a Glance poster and article, Betsy McIntosh and Michael Ostap illustrate the proposed functions of metazoan myosin-I molecular motors, examining structural, biochemical, mechanical and cell biological evidence.
Smaller broods promote quicker endothermy
Jan-Åke Nilsson and colleagues showthat the ability of blue tit chicks to generate their own body heat is influenced by the size of the brood they grow up in, with chicks from smaller broods becoming endothermic earlier than those from larger broods.
From autumn 2016, a postdoctoral research position is available in the Panfilio lab to investigate morphogenesis of the insect extraembryonic (EE) membranes. These simple epithelia are highly dynamic in their role as transient, protective covers for the embryo. The aim is to understand how EE morphogenesis works at multiple levels of biological organization, from cellular ultrastructure to coordination between tissue sheets. The primary research species for the project is the red flour beetle, Tribolium castaneum, an excellent model for studying interactions between developing epithelia via fluorescent live imaging and sophisticated genetic approaches (see the lab’s recent publications in eLife5:e13834, and Developmentdoi:10.1242/dev.133280).
extraembryonic cell shape changes during tissue remodeling
The lab is in the Institute for Developmental Biology, University of Cologne, Germany (lab web page: http://tinyurl.com/c24gpte), and has research and collaboration links with other evolutionary, developmental, and insect labs across the Biology Department of the university. With one million inhabitants, Cologne is an international, vibrant city that is well connected within western Europe.
Successful applicants will have a strong interest in developmental genetics and imaging, demonstrated by holding a Ph.D. degree and at least one first author publication in a relevant field. Required skills include standard developmental genetics techniques associated with RNAi, tissue staining, and conventional light microscopy. Experience with electron microscopy, quantitative analysis of multidimensional imaging data, transgenesis, and/or transcriptomics is desired. Depending on expertise and interest, there are possibilities for the applicant to develop his/her own research program among the topics of dynamic tissue adhesion, remodeling of the extracellular environment, and comparative transcriptional profiling. The working language of the lab is English, and strong oral and written communication skills are expected.
The position is for one year in the first instance with the possibility for annual extensions. The University of Cologne is an equal opportunity employer in compliance with German disability laws. Women and persons with disabilities are strongly encouraged to apply.
To apply send a research statement, CV, starting date availability, and contact details (including e-mail address and phone number) for two references as a single PDF file to Kristen.Panfilio@alum.swarthmore.edu. Informal enquiries to further discuss the position are welcome. All applications will be considered until 20 August 2016.
It seemed like an ordinary morning, with the lecture on Drosophila genetics due to start at 8 o’clock. But when the professor walked in something remarkable happened: instead of starting the lecture right away, he posed an unusual question: “One of the students of this class has a publication on this topic. George Streisinger, where are you?” But the student he was looking for was not there – he slept in. Lotte Sielman, sitting in the audience, was just as puzzled as her classmates about the curious incident. She could not have guessed that George Streisinger would soon become her husband and would contribute a number of exciting and important chapters to the history of modern molecular genetics.
Not to forget, the other author of the above mentioned Drosophila paper was one of the giants of the 20th century biology, Theodosius Dobzhansky. And if we look at the later works of Streisinger, this is not even unusual: being one of the most talented geneticists of his era, he built strong friendships and professional relations with almost all of the celebrated biologists of the 1950s.
From Budapest to Manhattan
Streisinger was born in Budapest on 27th December 1927. His parents were merchants: his father, Andor Streisinger dealt with leather goods and his mother, Margit Freund was selling ties. Little Gyuri (as he was called at that time) started his life in the city: his family first lived in the centre in one of the later yellow-star houses, then in the Castle district, followed by a few years in the center city again, where his father kept pigeons on the roof. (Andor Streisinger probably had a serious injury during WWI, as he was entitled to an injury allowance.) Gyuri started school at the Lutheran Primary School next door, and later followed brother Ervin (7 years his senior) to the legendary Fasori Lutheran Secondary School. (The fame of the Fasori School is due to the fact that it was also the Alma Mater for a number of other Hungarian-born luminaries of the era: Nobel-prize winners Eugene Wiegner and John Harsanyi, the mathematician John von Neumann, and the physicist Edward Teller.)
The Streisingers lived a typical upper-middle-class life. A family cook and a maid helped with the housework, but also took care of Gyuri – perhaps this was the origin of Streisinger’s later fondness for culinary delights. He was an omnivorous gourmand, who was ready to prepare or taste any kind of food. Well, almost any: kohlrabi was the exception. He hated it as a child, but if he did not eat it for lunch or dinner, it was back on his plate next morning.
The little boy was diagnosed with a heart murmur at an early age, which prohibited him from taking part in the PE classes or playing soccer with his friends. Instead, together with Ervin, they went out to catch butterflies in the mountains of Buda. These early voyages as a budding amateur naturalist became the inspiration for his later career.
His attraction to natural sciences would have been supported by the teachers in the Fasori School, however the turbulent history of the 20th century did not let this happen. After a few months in the famed institution, the increasingly severe discrimination against Jews forced the family to pack up and leave everything behind for a better life on the other side of the Atlantic.
Margit Freund and her sons George (on the left) and Ervin (on the right) a few years after their immigration. (Photo courtesy of Cory Streisinger).
They were not jumping in the great unknown, though. Margit’s two siblings, Fred and Viola have fled Hungary several years before, and they had already established a new life in New York. Fred worked for the General Oil Burner company, while Viola married a dry-cleaner, Anthony Burachio. Margit visited them in 1936, although probably she did not really expect to join them as an immigrant in a few years time.
The first to set foot in the New World was Ervin, freshly out of high school. He arrived on 8th October 1938, followed by his father on 25th November. Margit and George were held back for several months, most likely due to the difficulty to secure the necessary visas. (Being born in Losonc, at that time already part of Czechoslovakia, Andor Streisinger was under the jurisdiction of the Czechoslovakian travel quotas, whereas his Budapest-born wife and younger son had to wait for their turn within the Hungarian quota queues. Andor, with the help of Fred Freund, tried to lobby the American authorities several times, to issue the visa for Margit and Gyuri faster, but for little avail.)
Finally, on March 1939, the USS Manhattan arrived from Le Havre to Ellis Island, with the missing members of the family on board. The reunion, sadly, was not the happy meeting they had all hoped for. By this time Andor was quite sick, with metastasized gut cancer that spread to his spine. He was hospitalized the next day and in less than two months he died in the Long Island College Hospital.
The freshly widowed Margit Freund found herself with her two children in the middle of a huge and unknown metropolis. Erwin was able to look after himself, first as an employee in a tailor shop, but after a few years of college he enrolled into the Army. Margit and his younger son stayed for a few months with their relatives, and later moved to the 87th street. The mother, with the help of her sister found jobs as a dry cleaner and tailor. The housework was all left to young George.
The allurement of science
George mastered English extraordinarily fast, and just in a few years time, after taking one of the most competitive entrance exams in the city, he got enrolled into The Bronx High School of Science. Here, perhaps due to the earlier inspiration of the trips to the Buda mountains, he joined the Herpetology Club, which organized collecting trips to the Pine Barrens of New Jersey. His fondness for natural history was so obvious that he was soon hired by one of the fish experts of the New York Zoological Society, Dr. Myron Gordon. (The fish experience will become an important point of reference in Streisinger’s later life.) He relished working with Gordon, and due to his enthusiasm, after finishing high school, on the summer of ‘44 he got the opportunity of a lifetime: to work with Dobzhansky in the latter’s Columbia laboratory. The joint work resulted in the aforementioned paper, which was published in the Proceedings of the National Academy of Sciences.
Dobzhansky was fascinated about the possible role of sexual selection on the evolution of species, and the work of Streisinger was key to understand this. Young George has spent the summer with pairing males and females from different populations of a South-American Drosophila species to find out whether they attracted each other or not, by dissecting the females. As far as we can tell the renowned naturalist soon became more than a simple mentor for Streisinger, and he acted more like a father, taking the boy to his family holidays to California. But the most important influence on George’s life was his extended professional network: Dobzhansky helped Streisinger to gain access to Cold Springs Harbor, the cradle of modern molecular genetics, which became the defining moment in the young scientist’s life.
En early CV of George Streisinger. (Courtesy of Cory Streisinger.)
In the fall of 1944, aged only 16, Streisinger enrolled to Cornell University’s School of Agriculture. This might sound as an odd choice, but unlike other schools at Cornell the School of Agriculture was free to attend, with the small caveat that students had to spend some of their time with farmwork. And in the financially precarious situation faced by the Streisingers this was the only way George could actually enroll to the prestigious institution. Even this way he had to find other sources of income to get by: he worked regularly in the local coffee shops, and later tested the bacterial content of fresh milk in the Dairy Department. Ironically, the small caveat about the farmwork almost wrecked Streisinger’s career: while his classmates accomplished this work during the summer break, he was too busy toiling away in the laboratories of Cold Springs Harbor. And while this summer work helped him enormously in his later career, at Cornell he was forced to postpone his graduation and worked at a farm, shoveling manure during the coldest winter of the decade (something that made him proud even years later). Being a staunch pacifist, he also refused taking part in ROTC sessions, a decision that also contributed for his delayed graduation, but finally in the summer of 1950 he received his degree.
The Sielmans arrived to the US in 1938 also escaping the deportations of nazi Germany. Lotte and George found out soon that they had a lot of common, and after the first date they became an inseparable couple. They went together for bird watching in the morning, and in the spring their night rendez-vous often ended in the nearby marshes, where they observed the mating rituals of salamanders and frogs. They married in 1949, the day before Lotte’s graduation. A good friend was playing their favourite tunes on the bells of the chapel, and the wedding party was hold in a teacher’s garden, where an accordionist provided the music.
In the shadow of the bomb
The aftermath of the two atomic bombs, dropped after Germany’s capitulation (when most considered the war to be effectively over) caused serious soul-searching for many physicists working for the Manhattan project. Many considered the bombing unnecessary, a cynical exercise of power that was intended to send a message to the Soviet Union sacrificing tens of thousands. Many scientists turned away from physics for good and searched for other research topics. Influenced by Erwin Schrödinger’s essay, ‘What is Life?,’ for many of them biology would provide the next frontier. In his highly influential little book, Schrödinger tried to explain how simple physical and chemical reactions could explain the behavior of biological systems. One of the key problems, identified in the essay, was the nature of genetic information. No one understood at the time how this information was encoded on a molecular level (Schrödinger posited the existence of aperiodic crystals), and the book clearly catalyzed efforts to reveal this great unknown.
Answering the question required a new biological model, and under the influence of John von Neumann many opted to study the simplest model available, phages. (In a letter to Norbert Wienernek, the father of cybernetics, von Neumann pointed out that if scientists want to understand the information flow in living organisms, they need a simple model, such as a phage.)
Bacterial and phage genetics dominated the coming decades and delivered results (often followed by Nobel prizes) at an astonishing pace. Phages played a central role in demonstrating that DNA is the molecule of inheritance. Alfred Hershey’s and Martha Chase’s experiment complemented beautifully the bacterial work started by Frederick Griffith and finished by Oswald Avery. And while after Avery’s results there were still some who were adamant about the role of proteins in inheritance (DNA was considered just “too boring” to play such a central role in biology), the Hershey-Chase experiment silenced all the skeptics.
Hershey, together with two other colleagues, Salvador Luria and Max Delbrück (themselves immigrants, just like Streisinger), also founded the legendary “Phage group”. During the 1940s they organized summer courses at Cold Springs Harbor, and the list of attendees now reads as who-is-who for the molecular genetics of the 20th century. Seymour Benzer, Sydney Brenner and James Watson were amongst the first to come to CSH, as was Leo Szilard who also became interested in biology at the time. And amongst all these luminaries there was also George Streisinger.
Aaron Novick and Leo Szilárd (in the middle) during the 1953 Phage course. On the left, Max Delbrück is reading a newspaper, whereas on the right a young James Watson is sitting on the ground. (Photo courtesy of Cold Spring Harbor Library.)
Luria and Delbrück escaped the fascist and nazi regimes by emigrating to the US, where they soon became central figures of molecular biology. (Together with Hershey they received the Nobel prize in 1969 for their pioneering work). After a short detour at Columbia, in 1943 Luria started to work at the University of Indiana, where his first PhD student was Watson himself. The same year, in a series of inspired experiments, Luria and Delbrück demonstrated that mutations arise spontaneously in bacteria (something that was far from trivial in an era when many even questioned the existence of genes in prokaryotes).
The summer courses at CSH had a profound influence on the young Streisinger, and after finishing at Cornell, he started a PhD in Luria’s group. Probably equally importantly his friendships from the Phage group provided him with a fantastic social network for the rest of his life.
Phages
Streisinger followed Luria to the University of Illinois, where finally he could reunite with his wife. According to Lotte’s memoirs, this was also the time when George became involved in progressive political activism. He pursued this activism with a characteristic passionate rationalism till the end of his life. This was the time when he became deeply involved in the fight against desegregation – with motivated activist friends they provided evidence that in several neighbourhoods landlords choose renters based on skin color.
After receiving his PhD, George moved to California for a postdoc in Max Delbrück’s group. It wasn’t so much the research topic that attracted him to the West Coast as the realisation that everybody who counted in molecular genetics those days had gone through the Delbrück and Pauling labs at Caltech. During these three years (1953-1956) the family also expanded: soon after their arrival Streisinger’s first daughter, Lisa was born at the Pasadena Clinic, soon to be followed by Cory.
Streisinger’s research project at this time was to establish an experimental system for plant viruses, akin to the one that was already used for phages. The work progressed slowly, and in spite of the many experimental failures he established a fruitful barter system with his office roommate, Harry Rubin. Working with Rous’ Sarcoma Virus Rubin used chicken as his model, which complemented (in the kitchen, that is) very nicely the coconuts of Streisinger (used for their milk), thus for several weeks the Rubin and Streisinger families ate copious amounts of coconut chicken. This wasn’t the only extravagant gastronomical experiment, in which George took part at the time: during New Year’s Eve parties he cooked suckling pig in the departmental autoclave.
Finally, by the time everyone was fed up with chicken and coconuts, George, frustrated by the experimental failures, turned back to work again with phages. This led him to his first appointment, back at his alma mater, Cold Springs Harbor.
Thanks to the fame of the Phage group, by this time CSH has left behind the stigma of eugenics for good and became the Mecca of molecular biology. After the slow and quiet winter months the summer always brought in new people from all over the world, eager to collaborate with the local scientists, or to attend the Phage course. By this time, instead of the original Delbrück-Luria-Hershey trio, the course was taught by the resident researchers, including Streisinger himself.
The closing ceremony for one of the later Phage courses. Frank Stahl can be seen on the left and Geroge Streisinger on the right.
The three idyllic years at CSH were interrupted by a sabbatical year that the family spent in England. George was invited by Francis Crick to spend some time at the famed Cavendish Laboratories, together with Benzer and Brenner. Cambridge had a bubbling social scene at the time, with an endless number of programs and parties, often organized at the Crick’s home, “The Golden Helix”.
During their stay Benzer, Brenner and Streisinger came to agree that the English kitchen in the 1950s was awful. After his arrival Benzer suggested Crick that they should try out every day another restaurant, however soon they realised that in the absence of quality places the exercise was futile. Therefore, when a month later the freshly arrived Streisinger enthusiastically suggested the same thing, he was just met sarcastic laughs. But George did not give up the hope to eat quality food, so soon together with the Benzers and Renato Dulbecco they started a gourmet dinner-club. They went to great lengths to get the best local venisons – George once traveled to Yarmouth to by fresh clams for a dish of moules marinière. In the end the members of the dinner club were spending almost as much time with preparing meals as doing actual research.
Back from England Streisinger started looking for a new job, and the first offer came from Brandeis. It offered everything George needed, yet neither him, nor Lotte felt enthusiastic about the poshy Boston suburbs that were hard to square with the progressive worldview of the Streisingers.
During their sabbatical in Cambridge, the Streisingers toured the British countryside on a motorcycle with sidecar. (Photo courtesy of Cory Streisinger.)
However, while considering Brandeis an irrefutable offer came from Aaron Novick, the freshly appointed director of the Institute of Molecular Biology (IMB), in Oregon. Novick asked Streisinger to join him, to become one of the “Founding Fathers” of the new research center.
Novick graduated as a chemist in Chicago, where he was promptly recruited to the Manhattan project in 1943. This is where he met Leo Szilárd, who became soon a mentor for the young scientist. As for many fellow scientists, the detonation of the atomic bombs over Nagasaki and Hiroshima caused immense consternation and soul-searching for Novick. He wanted to get out of nuclear research as soon as possible and also became an ardent supporter of nuclear disarmament till the end of his life (for many years Novik edited the Bulletin of Atomic Scientists promoting a pacifist worldview).
The change in career was facilitated by Szilárd’s invitation to join him in attending the phage course at CSH, or in other words to join him for “an adventure in biology”. The opportunity excited Novick, who saw in it a chance “to exchange death for life”.
After the war Novick returned to the University of Chicago, where in 1959 he got the offer from University of Oregon to become the director of the freshly minted IMB (as the story goes, this was the first institution with “molecular biology” in its name). He also got a free hand in selecting his coworkers, and this is how he met Streisinger, who was suggested by a former colleague, Frank Stahl, himself a new IMB fellow.
Eugene, the home of IMB, and its surroundings enchanted the Streisingers who accepted the offer almost immediately. Soon they were settling into a small loghouse, close to the city. The new house and the nearby barn became the home to a number of animals as well, and George took great pleasure in his Nubian goat herd. This actually brought him fame, and soon he spent several weekends every year acting as a goat judge at different country fairs.
The idyllic life had a positive impact on George’s research as well. At IMB he continued his phage research, and his new results were getting the acclaim of his peers. This is demonstrated by a recommendation letter that Watson wrote for him when George was applying for a promotion: “George Streisinger is now one of the most productive geneticists in the United States. The work of his group at Eugene on the circular nature of the T4 chromosome is first rate. I wholeheartedly concur in your proposal to promote him to the rank of full professor.” The same esteem can be found in Streisinger’s correspondence with Francis Crick, where they discuss some of George’s more recent results that contributed to the deciphering of the genetic code, but also recognized that mutations at repetitive DNA sequencing are due to the stuttering of the polymerase, a phenomenon still recognized as ‘Streisinger slippage’ in the literature.
Watson’s recommendation letter for Streisinger. (Courtesy of Cold Spring Harbor Library.)
The more collaborative and much less competitive spirit of the era is also demonstrated by the fact that while Streisinger happily shared his data at conferences and private correspondence, he was much slower in writing them up into formal papers. This was the reason why at an IMB jamboree he was presented with “Doctor of Delayed Publications” honor. However, the greatest gap in his publication record in the 1970s was not due to his tendency to procrastinate, but because of his desire to “do something big”. And this was nothing less then establishing a new vertebrate model organism.
Fish
By the mid-1960s the phage as a genetic system showed signs of exhaustion. Most of the work instrumental in understanding the basics of gene function and regulation have been either already accomplished, or were close being so. Obviously there were still many questions to be answered, but many grandees of the phage field decided that they needed other models to properly examine these questions. As Sydney Brenner put it in a letter to Max Perutz, it was only a matter of time before “all the chemical details of replication and transcription will be elucidated. Because of this, I have long felt that the future of molecular biology lies in the extension of research to other fields of biology, notably development and the nervous system.“
Accordingly, some of the biggest names decided to leave the crowded phage-field and started pioneering work to find other model systems. Seymour Benzer started to study the nervous system of the fruitfly, whereas Brenner elevated a simple nematode, Caenorhabditis elegans, to the pantheon of genetic model organisms. Streisinger was also enthusiastic to change, but unlike his friends, he firmly believed that due their individual characteristics non-model organisms will be only of limited use in understanding and studying the complex nervous system and behaviour of vertebrates. To understand vertebrates, one needs a vertebrate model, and that is how zebrafish came into the picture.
Truth to be told, it wasn’t Streisinger who first used the zebrafish as an experimental organism. This honor most likely goes to a researcher at Wayne State University, Charles W. Creaser. Back in 1934 Creaser already argued that this little fish from Southeast Asia, a favourite of aquarists all over the world would be a suitable model to study embryology. Its small size, external fertilization and fast development make it an ideal model species for vertebrate developmental biologists. During the next couple of decades zebrafish research progressed at a slow space, with an occasional paper here and there. But at this time, it was only one of the many bony fish species that were used in research (and its relatives, goldfish and carp were far more popular). Nothing suggested that soon it would have a prominent role in biology.
Streisinger learned a lot about fish as an assistant of Myron Gordon, and due to that experience he became a hobby aquarist himself. Thus his choice of zebrafish might not seem too surprising. However, despite the fact the zebrafish was already used in lab experiments, the success was far from guaranteed. Due to the high risk of failure, at the beginning it was really hard for Streisinger to find colleagues who would join him in his new enterprise. Considering that it took many failures, and almost nine full years to get the system going, the critics might have had a point. (It is worth pointing out that probably the IMB lead by Novick was the only place at that time that would have had the patience and confidence to sustain Streisinger’s work for such a long time. But Novick not only personally supported George, but also convinced other IMB groups to help him out as they could.)
The university provided a World War II quonset hut to facilitate the new project. This is where Streisinger and his assistant, Charlene Walker worked out the standard conditions for maintaining and breeding zebrafish. As the hut became overheated during the summer, and too cold during the winter, the biggest challenge was to provide constant temperature. Against the heat they sprinkled water on the surface of the hut, whereas during winter dozens of electric heaters were installed (and often got short-circuited in the humid environment). Yet the problem of the temperature was easy compared with the headache that parasites caused to the experimental duo. These wiped out fish tanks again and again. In order to fight them, George and Charlene started to add more and more chemicals to the water. These restrained the parasites, but also interfered with fish development. Finally, after many trials and errors, Charlene was able to standardize the conditions, and George could start with the real experiments.
As he put it in a 1974 NSF application, his goal from the beginning was to “study features of the organization and embryological development of the vertebrate nervous system through the use of mutant strains.[He was] particularly interested in the mechanisms leading to the formation of specific synaptic connections and in the nature of the signals that guide specific axons to particular target sites.” He wanted to create mutants with nervous system abnormalities, but in order to create homozygous animals for rare mutations he had to overcome their diploidity.
A huge advantage of the worm, C. elegans, is that most of the individuals are hermaphrodites, capable of self-fertilization. This makes them well suited to study recessive mutations. Although hermaphroditism is almost nonexistent in vertebrates, there are quite a few examples for parthenogenesis, which also makes the analysis of mutations much easier. However, zebrafish is not one of the parthenogenic species. Streisinger, obviously, was aware of this, but he was convinced that he could find a way to jump start the development of unfertilized zebrafish eggs, thus in effect making them parthenogenic. After a few abandoned trials George and Charlene figure out the right conditions for the procedure, and they started producing mutants with vigour. The first results were published on the frontpage of Nature in 1981, and a new science was born: zebrafish genetics.
The first zebrafish clone aroused the interest of the general press, exemplified by the cartoon of the Chicago Tribune.
The popular press also enthused, but for different reasons: as the new fish became practically the clones of their mothers, soon Streisinger was inundated with questions about the ethics and politics of cloning. (The most unexpected admirers of the work were activist from the radical feminist groups of the 1970s. In their interpretation Streisinger’s work proved that the “male part of the conception process is irrelevant.)
In one of the many interviews he said: “We’ve developed a vertebrate system for doing interesting genetics. It just hadn’t occurred to us by any stretch of imagination that our work would generate any of this… There is a general worry among scientists, that overuse of cloning would lead to a loss of variety in food crops, but almost anything could be misused so it is very difficult to know at the time whether the good you get out of something will outweigh the bad.” He considered human cloning at the time unrealistic, but he also said that he had much greater, immediate worries. “I see the danger of global nuclear war imminent. The use of poison warfare, the widespread use of chemicals that may be hazardous, the lack of any serious attempt to deal with population growth, the lack of any real concern about just incredibly unequal distribution of wealth.” These were all things that should have been more important for newspapers than zebrafish cloning.
After the University of Illinois years Streisinger never stopped to be civil rights activist, and the atmosphere at IMB was very supportive for this sort of activities. Novick’s campaign against nuclear proliferation was only the most prominent of the campaigns originating within the walls of the institute. IMB researchers became important allies of EPIC (Eugene Peace Information Center), and none of them was as vocal as Streisinger himself. He became the local representative of the national Turn Toward Peace lobby group, but his major contribution came in the fight against toxic herbicides.
In 1962 Rachel Carson’s “Silent Spring” successfully focused the attention of the public to the potential health hazards of agricultural chemicals. The book that ushered the creation of EPA, was drawing attention to the hazard of DDT, but it was obvious from the start that there are many other potentially dangerous pesticides. In the Oregon woods the local Forest Service used dioxin-based chemicals against weeds, and this unsettled the locals. Many of them were afraid that these chemicals (all related to the infamous Agent Orange) could accumulate in the plants and get into the groundwater. EPA originally concluded that there was no reason to worry, but many were unconvinced. They asked IMB researcher to help to prove that dioxin is toxic and dangerous. Streisinger became the star witness of the trial, and argued successfully that in the case of dioxin there is no such thing as “safe dosage”, as even at very low concentrations that chemical causes chromosome breakage, thus it must be mutagenic. The judge accepted this, and ordered the EPA to perform new, systematic studies. These, in turn, lead to the banning of dioxin-based herbicides in a few years time.
Streisinger’s opinion carried weight, because as a geneticist he studied often the mutagenic potential of different molecules. These observations were important not merely as toxicological observations (although, undoubtedly, zebrafish is still an important model in toxicology). George wanted to understand how mutagens work, so he could use them in his project to create mutant fish lines.
One of the last photos of George and Erwin Streisinger was taken in 1981 on the porch of George’s loghouse. (Photo courtesy of Cory Streisinger.)
Besides research and civic duties, George took teaching extremely seriously as well. Usually he started his lectures at 8 am, so he could do experiments the rest of the day. But even after several decades of experience he still worried how the students will receive the material, thus in order to get well prepared, he woke up at 4. His methods of teaching were way ahead of his time: one could easily think the Dance your PhD competition is something original, a creative 21st century idea, yet four decades ago Streisinger collaborated with the local Dance Department to illustrate his lecture about protein synthesis.
By the early 1980s as the founder of a zebrafish research, Streisinger was becoming one of the star researchers of IMB. His efforts were not only recognized by his peers (by this time many other IMB groups started to work with fish), but also by the wider public of Oregon. However, as the IMB was preparing to celebrate it’s 25th birthday a tragedy happened: on 11th August 1984, a few weeks before the ceremony Streisinger died during a scuba diving exam near the Oregonian coast (that old heart murmur never disappeared completely). The IMB birthday promptly became a Streisinger memorial. It was attended by the very best of the 20th century molecular biology. From Hershey to Watson they all wanted to pay their tributes.
Without its father, the future of zebrafish research became in peril. It was up to the IMB colleagues to carry the torch, and they promptly did so. Thanks to their work as well, today it is obvious that zebrafish fulfilled the expectations. It became one of the most widely used genetic model organisms, used as much to study simple developmental processes as to model complex human disease. But the thousand or so zebrafish labs around the world today exist only because of that one quonset hut in Eugene, where George and Charlene heroically worked out the basics of zebrafish research.
Epilogue
Although Streisinger emigrated at a very young age, he never completely forgot where he came from. The recipes of Hungarian pastries were always held in high esteem in the family cookbook, and with Margit and Erwin George they always spoke Hungarian (after many decades he still spoke English with a characteristic Hungarian accent). Yet, we know of a single occasion when he visited Hungary after 1939. In the 1960s he arrived to Budapest for a lecture, together with Frank Stahl. The visit was a good opportunity to revisit his city of birth and meet some relatives. As Frank Stahl remembers, they walked a lot on the banks of the Danube, ate in small restaurants, and Streisinger was fond of recalling his memories of Gellért bath. At the lecture George was adamant to speak in Hungarian, but probably his mixed accent and the topic that was completely novel to his audience (biology in the Eastern bloc was still under the influence of lysenkoism) just confused the people in the room. So in a few minutes time he was asked to continue in English.
This is an edited version of a Hungarian language text that appeared last year. We are republishing it because George Streisinger is such an important person for the zebrafish community – and next year we want to celebrate his 90th anniversary at the 10th European Zebrafish Meeting. (If you are interested in zebrafish genetics, please come!)
To write this post I relied heavily on Lotte Streisinger’s book “From the sidelines”, Jim Endersby’s “A Guinea Pig’s History of Biology” and Errol Friedberg’s “Sydney Brenner – A biography”. Some facts were also taken from George Streisinger’s autobiographical essay, “Terminal Redundancy: All’s Well That Ends Well”. I am extremely grateful to Cory Streisinger for sharing the countless family stories and photos. I also want to thank Frank Stahl, Ferenc Müller and Julianna Vig for helping me with this story. Photos are used with the permission of the Streisinger family and Cold Spring Harbor Library.
The Paridaen lab at the European Research Institute for the Biology of Ageing (ERIBA) in Groningen, The Netherlands, is looking for a PhD student on Stem cell biology.
Stem cells act in developing and adult organisms to produce the proper number of specialized cells in the body. It is important that stem cells select an appropriate division mode to ensure proper development, maintenance and repair of tissues. In the Paridaen lab, we aim to understand the mechanisms underlying stem cell decisions in developing and ageing tissues. We employ single cell analyses techniques to study stem cell behaviour in the developing brain, using zebrafish as our main model system. In particular, we are interested in the role of fate-determining factors such as signalling pathways and subcellular structures in symmetric and asymmetric stem cell divisions.
ERIBA is a newly established institute at the University Medical Centre Groningen (UMCG). The mission of ERIBA is to better understand the molecular mechanisms that result in age-related decline and age-related diseases. We aim to develop novel strategies to prevent or combat age-related diseases and to provide evidence-based recommendations for healthy ageing. The research teams are working with different model systems and technology platforms and share their knowledge and expertise to accelerate discoveries. The ERIBA is housed in a new building and is part of the central UMCG campus in the vibrant University City Groningen in the north of the Netherlands.
Fertilization marks the start of life. This is followed by highly coordinated epigenetic reprogramming that allows protamine-histone exchange, zygotic genome activation, and the generation of a totipotent embryo. However, the true state of chromatin at the level of DNA during this crucial period is a long-standing mystery.
Our lab is dedicated to understanding epigenetic reprogramming in early development. But for a long time, we were struggling to find the right tools. When I first heard about ATAC-seq developed by the Greenleaf and Chang groups in Stanford, I was very excited as this is exactly the approach that we were looking for. I brought this up to my PI and we quickly decided to give it a try. We started by testing it in various numbers of mESCs, using either crosslinked or native chromatin from either frozen or fresh samples as well as different kinds of detergent in lysis buffer. In our hands, it appeared that ATAC-seq worked best on native chromatin of fresh samples from 100 to 1,000 cells with 0.15% to 0.5% NP-40 in lysis buffer.
We then began to apply ATAC-seq to mouse early embryos. Working together with another graduate student Bo Huang who has extensive experience in mouse embryos, we collected the early 2-cell, 2-cell, 4-cell, and 8-cell embryos, as well as ICMs isolated from blastocysts. However, we were surprised to see that the sequencing libraries were strongly contaminated by mitochondrial DNA, which occupied up to 99% of the reads. This is mainly due to the large quantities of mitochondria inherited from oocytes. My colleagues and I then spent almost a year to test several approaches to reduce mitochondrial contaminations. Most methods we tried did not meet our expectation, until we developed CARM, the Cas9-Assisted Removal of mitochondrial DNA. The 114 sgRNA evenly targeting 16kbp mitochondrion genome is designed to introduce the cutting event on mitochondrion DNA fragment in ATAC-seq library, so that these reads could not be sequenced due to the lack of sequencing adaptor on the both end of the fragments. We were very happy to see that CARM significantly reduced the about 70% of mitochondrial DNA at that time. Developing a new method is never easy, but the support from my mentor enabled me to proceed without any hesitation. He always encourages us to take the challenge to solve the problem that matters.
This story is now published on Nature as an article in June 30 (Wu & Huang et al., Nature 534 (652-657)). There are several interesting discoveries that we have made in this study.
Schematic of ATAC-seq and CARM for probing accessible chromatin in mouse preimplantation embryos
Firstly, we found that unlike asymmetric DNA methylomes between the two alleles throughout preimplantation, accessible chromatin landscapes already become comparable between the two parental alleles after the 2-cell stage. Secondly, the open chromatin in early embryos occurs not only at the transcription start sites (TSSs) but also at the transcription end sites (TESs) of active genes, indicating a unique and unknown regulation mechanism in early development. Thirdly, we identified putative enhancers that are active in early development, through which we derived candidate transcription factors that may regulate early development. We validated the roles of two such regulators, Gata4 and Nr5a2, in lineage-specific transcription programs in preimplantation development. Finally, in early 2-cell embryos where major ZGA has not started, we found many large open chromatin domains (up to 110kb) over regions showing repeat-driven promiscuous transcription. These data support an unusually permissive chromatin state at this stage.
Taken together, our findings unveiled highly dynamic spatiotemporal configuration of chromatin states in early mammalian development. However, we believe this is just the beginning. Future investigations are warranted to fully unlock the mechanisms and the functions of these unique regulatory modes. As a graduate student, this was truly an exciting journey for me. The happiest thing is to make discoveries in a world full of unknowns.
Here are the highlights from the current issue of Development:
Gestational stress: at the heart of birth defects
Congenital heart disease (CHD) is the most common form of human birth defect, yet the genetic and environmental factors that contribute to CHD remain poorly understood. Here, Sally Dunwoodie and colleagues investigate how gestational hypoxia affects heart development in mouse embryos (p. 2561). They reveal that the exposure of developing mouse embryos to short-term hypoxia in uteroresults in heart defects, notably perturbations to the outflow tract (OFT). These changes are mediated by altered cell proliferation and FGF signalling in the secondary heart field (SHF), which contains progenitor cells that contribute to the OFT. The authors further report that hypoxia leads to rapid induction of the unfolded protein response (UPR) in SHF cells. This, in turn, causes a global decrease in protein translation and may contribute to the reduced levels of FGFR1, and hence FGF signalling, observed in SHF cells following exposure to hypoxia. Together, these results suggest that hypoxia-mediated UPR induction during pregnancy can give rise to CHD. Given the key role of FGF signalling during embryogenesis, these findings also have important implications for understanding birth defects that affect other organs.
Shox2 goes out on a limb
Hox-TALE factors are involved in patterning the vertebrate limb but precisely how they regulate specific and regional gene expression patterns is unclear. Here, on p. 2548, YiPing Chen and co-workers uncover a limb patterning transcriptional programme that is coordinated by the transcription factor Shox2. The researchers demonstrate that, although Shox2 is expressed in mesenchymal progenitors of multiple cell types in the proximal limb, its deletion specifically in the osteogenic lineage causes limb defects and loss of the stylopod – the most proximal region of the limb. ChIP-Seq analyses indicate that Shox2 binds predominantly to limb-specific enhancers that are involved in skeletogenesis; these regions are also co-occupied by Hox-TALE factors. Finally, the authors show that Shox2 is expressed in a gradient that is complementary to that of TALE factors and that it represses the expression of TALE factors in the stylopod. Overall, these observations, together with other findings, highlight the existence of a Shox2-coordinated transcriptional programme that functions to pattern the vertebrate limb and provide insights into the ‘enhancer grammar’ that is used to mediate specific transcriptional outputs.
Pinning down spindle orientation
Spindle orientation is regulated by a conserved mechanism in which Pins/LGN anchors Mud/NuMA to the cell cortex. This pathway has been assumed to operate in nearly all animal cell types but here (p. 2573) Daniel St Johnston and colleagues reveal that Pins is not required for spindle orientation in the Drosophila wing disc. Using live imaging, they first discover that spindle angles in the wing disc vary widely; spindle angles are initially random but gradually align with the plane of the tissue as cells enter anaphase, highlighting that spindle angles are not accurate predictors of division orientation until this point. Importantly, the researchers reveal that spindle orientation does not require Pins, or aPKC, Dlg or Lgl. They further report that Mud is able to localize to the cell cortex in the absence of Pins, suggesting that a parallel but as yet unknown mechanism must act to localize Mud in wing disc cells. In summary, these surprising results indicate that a Pins-independent mechanism can orient the mitotic spindle in the Drosophila wing disc and lead the authors to propose that this system provides robustness to this rapidly developing epithelial tissue.
A sticky situation in plants
Cell-cell adhesion in plants is known to be regulated by pectins, and levels of homogalacturonan (HG; the main component of pectins) within the cell wall have generally been linked to cell adhesion. But how is cell-cell adhesion maintained and regulated in the face of the dynamic cell wall remodelling that takes place during cell growth and division? Here, Grégory Mouille and colleagues investigate this issue (p. 2536). Using a cell adhesion defect suppressor screen, they identify a putative O-fucosyltransferase – an enzyme that mediates the transfer of sugar residues onto substrates – that regulates cell adhesion in Arabidopsis thaliana. They further reveal that mutations in the gene encoding this enzyme or another putative O-fucosyltransferase perturb cell adhesion. Importantly, a comparison of mutant and suppressor lines suggests that cell adhesion does not rely on HG content per se. Based on their findings, the authors propose a model in which a pectin-related signalling pathway, rather than simply HG levels, contributes to the control and maintenance of cell adhesion during plant growth and development.
Plus:
An interview with Enrico Coen
Enrico Coen CBE FRS is a Project Leader at the John Innes Centre in Norwich, UK, who uses a variety of approaches to study patterning and morphogenesis in plants. We met with Enrico at the Spring Meeting of the British Society for Developmental Biology, where he was awarded the Waddington Medal, to ask him more about his career and his passion for art and book-writing. See the Spotlight article on p. 2479.
Exosomes in developmental signalling
Cells can signal by releasing exosomes – extracellular vesicles containing bioactive molecules such as RNA, DNA and enzymes. Recent work has suggested that exosomes can also carry signalling proteins, including ligands of the Notch receptor and secreted proteins of the Hedgehog and WNT families. Here, Ian McGough andJean-Paul Vincent describe various types of exosomes and their biogenesis and critically assess the role of exosomes in developmental signalling. See the Review on p. 2482.
Direct neuronal reprogramming: learning from and for development
The key signalling pathways and transcriptional programmes that instruct neuronal diversity during development have largely been identified. Here, Magdalena Götz and co-workers discuss how this knowledge has been used to successfully reprogramme various cell types into distinct types of functional neurons, and the extent to which direct neuronal reprogramming recapitulates embryonic development. See the Review on p. 2494.


 (1 votes)
(1 votes) (No Ratings Yet)
(No Ratings Yet)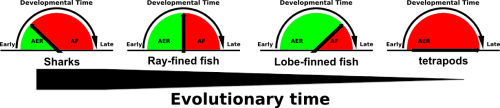
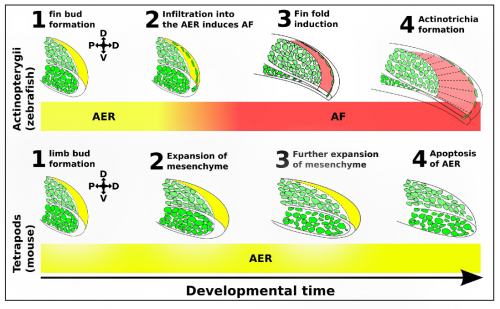

 Mice have been the traditional model of choice
Mice have been the traditional model of choice Neurofibromatosis type 1 is characterised by nerve sheath tumours but also has significant clinical heterogeneity. Robert Kesterson and colleagues
Neurofibromatosis type 1 is characterised by nerve sheath tumours but also has significant clinical heterogeneity. Robert Kesterson and colleagues 
 Maternal effects of chromosomal abnormalities
Maternal effects of chromosomal abnormalities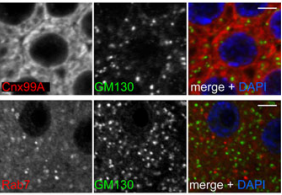 traffic
traffic
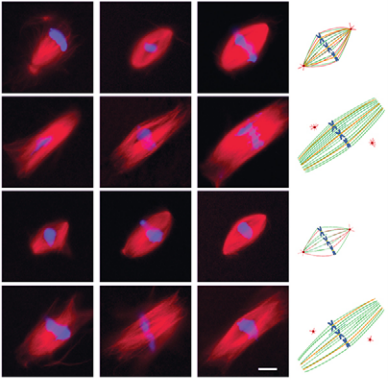 perm centrosomes defines spindle assembly
perm centrosomes defines spindle assembly

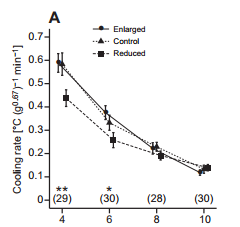









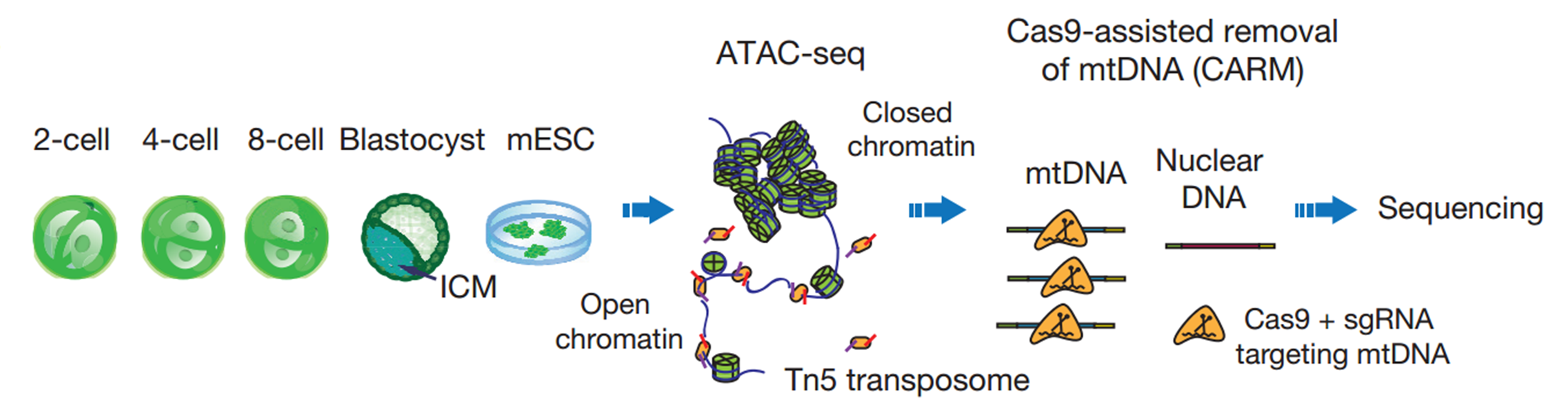




 Enrico Coen CBE FRS is a Project Leader at the John Innes Centre in Norwich, UK, who uses a variety of approaches to study patterning and morphogenesis in plants. We met with Enrico at the Spring Meeting of the British Society for Developmental Biology, where he was awarded the Waddington Medal, to ask him more about his career and his passion for art and book-writing. See the Spotlight article on p.
Enrico Coen CBE FRS is a Project Leader at the John Innes Centre in Norwich, UK, who uses a variety of approaches to study patterning and morphogenesis in plants. We met with Enrico at the Spring Meeting of the British Society for Developmental Biology, where he was awarded the Waddington Medal, to ask him more about his career and his passion for art and book-writing. See the Spotlight article on p.  Cells can signal by releasing exosomes – extracellular vesicles containing bioactive molecules such as RNA, DNA and enzymes. Recent work has suggested that exosomes can also carry signalling proteins, including ligands of the Notch receptor and secreted proteins of the Hedgehog and WNT families. Here,
Cells can signal by releasing exosomes – extracellular vesicles containing bioactive molecules such as RNA, DNA and enzymes. Recent work has suggested that exosomes can also carry signalling proteins, including ligands of the Notch receptor and secreted proteins of the Hedgehog and WNT families. Here,