The people behind the papers – Sa Geng & James Umen
Posted by the Node Interviews, on 19 April 2018
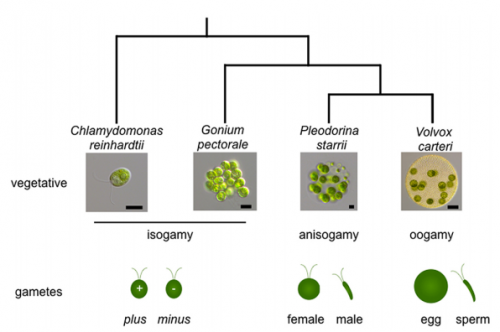
The transition to multicellularity in eukaryotes appears to be intimately linked to the transition from isogamy (gametes of the same size) to anisogamy (gametes of distinct sizes), and indeed to oogamy, a form of anisogamy with a large, immotile egg and a motile sperm. Volvocine algae provide a useful and fascinating model to study how this transition occurred, and a new paper in Development explores the genetics of the process in Volvox and some of its relatives. We caught up with first author Sa Geng and his PI James Umen, Joseph Varner Distinguished Investigator at the Donald Danforth Plant Science Center in St. Louis Missouri, to hear the story behind the work.

James, can you give us your scientific biography and the questions your lab is trying to answer?
JU I was a PhD student at University of California, San Francisco where I studied yeast pre-mRNA splicing in Dr. Christine Guthrie’s laboratory. I moved to Dr. Ursula Goodenough’s laboratory at Washington University as a postdoc to work on the mechanism of chloroplast DNA inheritance and its control by mating type in the green alga Chlamydomonas. During that time we made a somewhat fortuitous discovery of a mutation in the Chlamydomonas retinoblastoma tumor suppressor gene that initiated an interest in cell size control, a problem which we have continued to work on in my own laboratory. During my stay in the Goodenough laboratory we had joint meetings with Dr. David Kirk’s group who were doing research at the forefront of Volvox developmental biology. My exposure to Volvox and my attendance of a class on the emerging field of evolution and development catalyzed a long-term interest in pursuing an integrative program of cell, developmental and evolutionary biology in volvocine algae (i.e. Chlamydomonas, Volvox and other multicellular relatives) in order to gain a better understanding of how ancestral genetic and cell biological mechanisms are co-opted and how new ones evolve during the transition to multicellularity.
A major question associated with the transition to multicellularity is the evolution and origin of male and female sexes; but tackling this problem seemed like a distant goal when I began my first independent position at Salk Institute in San Diego in 2002. By the time I moved my laboratory to Danforth Plant Science Center in 2011 volvocine algae had entered the era of modern genomics which gave us new tools and information for investigating the transition to multicellularity and evolution of sexes. Besides our work on cell size control and evolution of multicellularity, we also became interested in the control of intracellular carbon partitioning in algae which has some important practical implications for sustainable production of lipid-based biofuels and other bioproducts.
And Sa, how did you come to be involved with this project?
SG When I joined the Umen lab, the function of Volvox MID had not been tested and some of the important methods we needed such as transformation of Volvox and sexual crosses were not yet firmly established. After optimizing those methods we figured out the spermatogenesis function of MID in Volvox and I became curious how MID evolved in algae and when it acquired the function to drive spermatogenesis. To answer these questions, I did cross-species MID transformation, putting GpMID and PsMID into female Volvox.
What makes volvocine algae useful models for evolutionary developmental biology?Are they easy models to work with on a day-to-day basis?
SG&JU From an evolutionary and taxonomic perspective volvocine algae are unique in being closely related, yet encompassing a graded series of multicellular innovations. They range from the well-studied model unicellular alga Chlamydomonas reinhardtii, to simple genera with small groups of coherently-organized undifferentiated cells, all the way to species like Volvox carteri that evolved a relatively complex pattern of morphogenesis, complete germ-soma division of labor, and oogamy. The haploid genomes of Chlamydomonas, Volvox, and other sequenced volvocine species have low genetic redundancy (no whole genome duplications) and are quite similar to each other in terms of gene content and gene number, prompting the question of how evolutionary innovations arose among organisms with such similar genetic toolkits. Both Chlamydomonas and Volvox (and other volvocine species) are relatively easy to culture and have short generation times. Besides classical genetics they have an expanding set of molecular genetic tools such as transformation, RNAi and CRISPR-based gene editing. Using these tools, it is now possible to test hypotheses about how gene/protein functions change (or not) in association with developmental innovations.
Can you give us the key results of the paper in a paragraph?
SG&JU Prior to our study we knew that a transcription factor called Mid (minus dominance) was a key mating type or sex determinant in volvocine algae. The Mid ortholog in unicellular Chlamydomonas (an isogamous species) specifies minus mating-type differentiation, while the Volvox Mid ortholog specifies spermatogenesis; but the Chlamydomonas MID gene was non-functional when expressed in Volvox. These results suggested to us that the spermatogenic function of Mid might have coevolved with sexual dimorphism. In our newly published study we expanded our cross-species complementation experiments and found that MID genes from two relatives of Volvox which are more closely related to it than Chlamydomonas—isogamous Gonium pectorale and anisogamous Pleodorina starrii—could both induce spermatogenesis in Volvox. The discovery that Mid from an isogamous species could function in Volvox spermatogenesis was unexpected and has forced us to rethink our model for the role of Mid and the Mid pathway in sex determination.
Since Mid proteins did not drive the transition to oogamy in volvocine algae, do you have any idea about which factors/genetic changes did?
SG&JU We are pretty sure that the number of Mid-controlled genes in Volvox has expanded substantially compared with Chlamydomonas and other isogamous genera, which makes sense because spermatogenesis involves the deployment of developmental programs that have no direct analogs in isogamous species. If Mid itself did not change substantially in the transition from isogamy to anisogamy/oogamy, then perhaps its hypothetical partner protein(s) changed to enable more target genes to be controlled. Alternatively, if a few key developmental TFs from Volvox gained Mid binding sites in their promoters, then perhaps those cis-regulatory changes were sufficient to indirectly expand the Mid network without requiring any major changes in Mid DNA binding specificity or changes its partner protein(s). We are now actively pursuing the identity of Mid target genes and Mid interacting proteins to get at this question.
What does your work suggest about evolutionary divergence of transcription factors following species divergence?
SG & JU This is an interesting topic that prompts some thought about the concept of homology and origins of phenotypic novelties. For Pax6/Eyeless the proteins from flies and vertebrates retained a high degree of sequence similarity and functional interactions, perhaps because they evolved as parts of a regulatory module for specifying eyes that was already highly constrained in their common ancestor and/or predisposed to evolve via downstream interactions, just as we think the Mid network did in volvocine algae. On the other hand, a small number of changes in the conserved plant TF Leafy was sufficient for neofunctionalization by altering its DNA binding site. In the case of Mid proteins in volvocine algae they are much faster evolving than Pax6/Eyeless or Leafy, so we were expecting that there might have been a major change in Mid function associated with the transition from isogamy to anisogamy/oogamy, but clearly Mid was not the major locus of change for the emergence and elaboration of this novel trait. Determining whether there are any patterns and rules for TF network evolution is still a major challenge.
When doing the research, did you have any particular result or eureka moment that has stuck with you?
SG Yes. I was intrigued to see that the induced female Volvox with a Gonium MID transgene produced sperm packets under microscope. Then I asked Dr. Umen to come over to check the culture on the slide. I noticed Dr. Umen was excited, but he was obviously not so sure with the result. Then we decided to do more experiments to confirm the finding. The lesson I learned here is to do more scientific analysis and experiments to make the data more convincing when unexpected results come up.
And what about the flipside: any moments of frustration or despair?
SG Frustration is always around when we are doing science, sometimes big, sometimes small. Transforming Volvox requires a lot of time and planning, and I felt frustrated when the transformation efficiency for Volvox was not ideal. When this occurred, I had to spend some time to figure out what’s the problem from the setting of the gene gun to the materials and reagents I used in the lab. I’m happy everything went well finally.
What next for you Sa?
SG My interest is still on the regulation mechanism of sex determination in volvocine algae. I will focus on the sex determination function of the Mid protein in Volvox and on the Mid network.
Where will this work take the Umen lab?
JU Some important new directions have emerged from our study. We think the Mid network expanded during the transition to oogamy in Volvox, but we don’t know how. We want to determine whether Mid binds DNA on its own or with other proteins, what are its target genes in different volvocine species, and how did its new targets get recruited during the transition to anisogamy and oogamy. On a more general level we are asking the question of how TF networks expand and integrate new inputs and developmental processes as they evolve more complex functions.
Finally, let’s move outside the lab – what do you like to do in your spare time?
JU My favorite hobbies outside of the lab are food (cooking and eating), tennis and fly fishing, plus anything else that gets me into the outdoors/nature.
SG I usually take care of my vegetable garden when I don’t need to go to lab to see my algae. I play table tennis with my wife almost every day, and I also go to fitness centre quite often recently.
Evolutionary divergence of the sex-determining gene MID uncoupled from the transition to anisogamy in volvocine algae. Sa Geng, Ayano Miyagi, James G. Umen
Development 2018 145: dev162537 doi: 10.1242/dev.162537
This is #40 in our interview series. Browse the archive here.
 (1 votes)
(1 votes)Get involved
Create an account or log in to post your story on the Node.
Sign up for emails
Subscribe to our mailing lists.