Applications are invited from highly motivated individuals who are interested in fundamental mechanisms of neurodevelopment and neurogenetic disorders. The focus of the project is to understand neural developmental and neurological phenotypes in mouse models ofTuberous Sclerosis Complex.The fellowship is in the laboratories of Sara Wilson at the Department of Integrative Medical Biology (IMB), and Leif Carlsson at Umeå Center for Molecular Medicine (UCMM) both laboratories at Umeå University, Sweden. The facilities provide an interactive modern environmentwith easy access to good core facilities. The fellowship is administratively placedat IMB, which is an interdisciplinary department focusing on questions in basic and medical sciences. The fellowshipis funded for two years and is available immediately. The working ‘day to day’ language in the laboratories is English.
Requirements:Individuals with a background in mouse genetics and handling, developmental biology, neuroscience, molecular and/or cell biology with a keen interest in developmental neuroscienceare encouraged to apply. Experience with rodent motor circuits or rodent motor or seizure behavior analysis is a big advantage.Technical experience with imaging, molecular biology, immunohistochemistry,in situhybridization, vertebrate embryonic model systemsare required. Technical experience with mouse embryo electroporation and /or neurite outgrowth/migration assayswill also be positively evaluated. The successful candidate will have or about to receive a Ph.D.in a relevant discipline, have good communication skills and be proficient in written and spoken English. The most successful candidatewill have ahigh level of motivation, be organized, rigorous and have the ability to work both independently and within a team.
Application:Please submit your application (reference 2019SW7) by 31stMay 2019 to sara.wilson@umu.seby sending the following documents as a single pdf file:
A short cover letter (not more than 1 page) to include a description of your research experience and suitability for the position.
Curriculum Vitaeincluding: publication list, technical expertise, names and contact information for three referees.
Informal enquiries may be directed to Dr. Sara Wilson (sara.wilson@umu.se) or Prof. Leif Carlsson (leif.carlsson@umu.se).
How can a genome’s cis-regulatory potential drive robust gene expression throughout embryonic development? Do topologically associating domains (TADs) truly contribute to gene expression by spatially constraining the regulatory actions of cis-sequences? And might phase separation by multi-valent proteins also be involved in the regulation of genes and genomic regions? The recent Company of Biologists Workshop on “Chromatin-based regulation of development”, organized by Benoit Bruneau (Gladstone Institutes, UCSF) and Joanna Wysocka (Stanford University), was the perfect playground to discuss important questions like these, and to stimulate interactions between early career and established scientists. Why was this Workshop so great to me (currently a postdoc in the lab of François Spitz, Institut Pasteur, France)? And why should you – a young investigator – apply to attend a future one? (more…)
Have you got a beautiful image of a developing organism you’d like to share with the world? Or a striking computer generated visualisation, or even something you’ve drawn or painted or stitched or crafted? We’re planning a Node calendar for the academic year 2019-20, and want to fill it with your images.
Entering is easy: simply send your image, along with a description of what’s being shown, tothenode@biologists.com, with the subject line ‘Calendar competition’.
Content-wise, the only limitation is that it should have a developmental biology angle to it. We’d love to have as much diversity in organisms and systems as possible. The image dimensions on the calendar are 24 cm L x 19 cm H, and some cropping may occur. One entry per person but individual labs can enter multiple images.
The deadline for submissions is 13.00 (GMT+1), Friday 10 May
We’ll collate all the entries and then run a public vote the following week. Good luck!
The idea for the calendar didn’t come from nowhere: Development used to print calendars each year and distribute them to researchers until it became economically unfeasible. But people would often ask us about them at conferences (or rather, complain about not having them any more!), and some of us in the Development team remember how nice the calendars looked hanging up in the lab before we left those labs for publishing. We hope our Node calendar will be equally beautiful and useful!
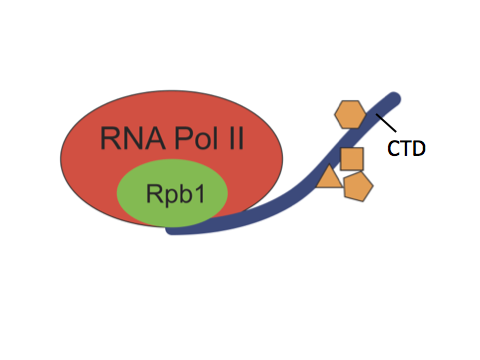
RNA polymerase II (Pol II) is the enzyme responsible for transcribing most genes in eukaryotes. The C-terminal domain (CTD) is a highly repetitive, unstructured domain on the largest Pol II subunit, Rpb1. It consists of numerous repeats of seven amino acids and serves as a binding platform for numerous factors involved in gene regulation, including chromatin modifying enzymes, transcription factors and RNA processing enzymes. Both length and sequence complexity of the CTD greatly vary among species of different lineages. For example, the CTD of S. cerevisiae has 26 repeats, the majority of which are consensus repeats of YSPTSPS. In contrast, mammalian CTDs have 52 repeats, the distal repeats are mostly divergent repeats whose sequence differ from the consensus at one or more positions.
The CTD (blue ribbon) is a repetitive, disordered tail on the Rpb1 subunit of RNA Pol II that interacts with a myriad of essential factors (orange).
The length and sequence complexity of the CTD were thought to regulate gene expression and be essential for the development of complex organisms. In the past decade, this proposition has been reinforced by several studies in mammalian cells. These studies showed that post-translational modifications occur on certain residues of the divergent repeats, some of which are recognized by factors only present in higher eukaryotes, and that mutations which prevent such modifications lead to misexpression of genes. Yet the significance of the divergent residues has never been systematically examined in the context of development.
Considering the above findings in mammalian cells, one would anticipate that a mouse would die if all of its CTD repeats were replaced with the consensus repeats that predominate the yeast CTD. However, generating transgenic mouse lines is very time and cost consuming. Instead, we decided to perform systematic mutagenesis of the CTD in Drosophila, an organism that is also highly genetically malleable but faster and cheaper to mutate compared to mice. In addition, 40 of the 42 CTD repeats in the fly diverge from the consensus, so it is an ideal model to interrogate the significance of divergent repeats.
The project was started at a time when CRISPR/Cas9 was not yet widely adopted by the fly research community, so initially we relied on RNAi knockdown to deplete the endogenous Rpb1 subunit. We also tested if the co-expression of an RNAi-resistant Rpb1 harboring mutations in the CTD would rescue the RNAi phenotype, making approximately 20 transgenic fly lines expressing various mutant forms of Rpb1 in an attempt to identify essential regions of the fly CTD. To our surprise, despite the high conservation in the amino acid sequence of the CTD within the fruit fly genus, most of our mutant CTD flies were viable, even when internal deletions removed up to 30% of the entire CTD. The only essential region we identified was an 8-repeat region encompassing the only two consensus repeats in the fly CTD (Gibbs et al., 2017). This suggests that the majority of the divergent repeats in the fly CTD are redundant.
Our initial systematic mutagenesis of the fly CTD emboldened us to test the idea that all divergent repeats could be replaced with consensus repeats. However, most previous findings in mammalian cells argue that the divergent CTD repeats are essential, so we still expected that flies would die with an all-consensus CTD. The most straightforward way to test this would be to simply replace the fly CTD with 42 consensus repeats. However, since many of the CTD repeats appear redundant, consensus repeats might be able to do what divergent repeats do, but with more or fewer repeats to achieve the same function. Therefore, we tested a series of consensus CTDs ranging from 10 to 52 repeats.
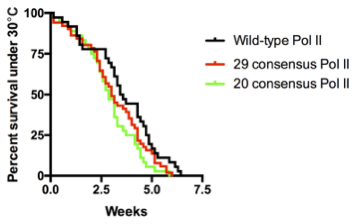
Surprisingly, we obtained normal looking flies with solely consensus repeats. This was achieved with only 20 to 29 repeats, which approximates the length of the yeast CTD. In contrast, flies with 42 consensus repeats, which matches the length of the wild-type fly CTD, barely survived to adulthood. In hindsight, we were fortunate to have tested varying numbers of consensus repeats because had we only tested the 42 consensus repeat CTD, we would have concluded that the divergent motifs are indispensable, which would have driven the project in a different direction. Additionally, flies with 52 consensus repeats died, whereas flies with the human CTD, which is also composed of 52 repeats but contains a mixture of consensus and divergent repeats, were able to survive. Therefore, it seems as though consensus repeats alone are sufficient, yet having too many of them is bad for fly development.
We were shocked by the results of the RNAi rescue experiment but at the same time concerned that residual endogenous Rpb1 might contribute to survival. Therefore, we decided to use CRISPR/Cas9 to mutate the endogenous Rpb1 gene. Remarkably, the CRISPR results agreed with our RNAi experiment. In particular, our flies with 20 to 29 consensus repeats did just as well as flies with the wild-type CTD even under temperature stress.
Death plots of fly lines with wild-type versus consensus Pol II when raised under heat stress (30°C). Pol II with 20 or 29 consensus repeats show comparable to wild-type survival rates.
As flies can survive with solely consensus repeats, consensus and divergent repeats should both be able to interact with the same group of factors. Our data also suggests that interactions provided by the consensus repeats are likely stronger, since in the case of an all-consensus CTD, fewer than half the number of wild-type repeats are needed for such interactions to occur properly. This fits nicely with the phase separation mechanism proposed by Weber and Brangwynne where protein interactions could be mediated by weak, multivalent interactions (Weber and Brangwynne, 2012). In reviews published when our project was being developed, Hnisz et al and Harlen et al proposed that such forces could also drive CTD:factor interactions (Harlen and Churchman, 2017; Hnisz et al., 2017). Also consistent with the phase separation theory was that the deleterious effects of having too many consensus repeats could be reverted either by shortening the CTD (as was the case with our truncated consensus CTDs), or by replacing some of the stronger repeats with weaker ones (as was the case with 52 consensus CTD versus the human CTD), both of which could reduce the overall valency of the CTD.
Coincidentally, our co-author Bede Portz had a chat with Stirling Churchman at the Cold Spring Harbor Mechanisms of Eukaryotic Transcription Meeting about our CTD mutagenesis in flies. Stirling was interested in our findings and was curious to know if the CTD by itself could target transcription sites. Bede thought that the fruit fly salivary glands would be an ideal system to test this. The salivary gland cells undergo rounds of replication without cell division, giving rise to polytene chromosomes where numerous copies of sister chromatids are fused together, which allows for visualization of each transcription site as a distinct band (or ‘puff’). Before us, John Lis’ group had shown that many transcription components such as Pol II are compartmentalized at heat shock puffs in salivary glands (Yao et al., 2006; Yao et al., 2007; Zobeck et al., 2010). Transcription compartments have also been documented recently using live super-resolution microscopy or in other systems where numerous copies of the same DNA sequences were introduced into a genomic location (Cho et al., 2018; Chong et al., 2018). However, the polytene chromosomes provide the power to visually determine a factor’s spatial location with respect to euchromatin, heterochromatin and nucleoplasm without having to rely on super-resolution microscopy or amplifying the number of copies of a candidate gene.
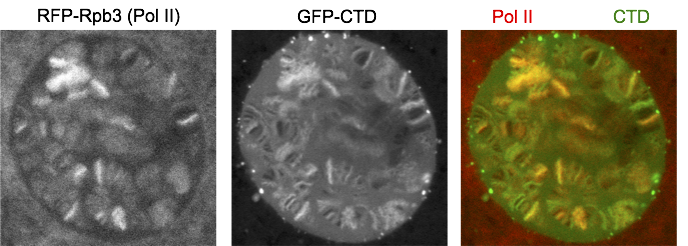
We fused just the CTD to GFP and expressed it in salivary glands. Interestingly, GFP-CTD colocalizes with Pol II on fixed polytene chromosome spreads. One possibility is that the CTD alone dynamically associates with the transcription sites. The alternative is that the GFP-CTD is stably bound by CTD binding partners that are abundant at transcription sites. To distinguish the two possibilities, we turned to live imaging and showed that GFP-CTD on puffs can rapidly recover upon photobleaching, suggesting that the CTD is dynamically recruited to transcription compartments. In addition, phase separation is likely the driving force for the behavior of the CTD, as 1,6-hexanediol, which has been shown to disrupt weak hydrophobic interactions, can disrupt the association of GFP-CTD to chromosomes.
Single Z confocal stack of a live nucleus from prepupal salivary glands with Pol II Rpb3 subunit labeled in red and the CTD alone labeled in green.
The partitioning characteristic of a molecule is influenced by its valency. This led us to investigate how the number of consensus repeats impacts the behavior of the CTD. To address this, we fused our series of consensus CTDs to GFP. Phase separation models predict that changes in valency could lead to pathological aggregates (Weber and Brangwynne, 2012). Indeed, our 42 and 52 repeats of consensus CTDs, which do not support normal function in the fly, showed less GFP on puffs and correspondingly more extrachromosomal foci when fused to GFP. Much of the extrachromosomal foci were neither recovered upon photobleaching nor dispersed by 1,6-hexanediol, suggesting that they were stable aggregates. Interestingly, none of our consensus CTDs partition into transcription compartments as readily as the wild-type CTD. These results suggest that the divergent repeats play a role in either preventing aggregation, or facilitating partitioning into transcription compartments, both of which could be forces that constrain the sequence conservation of the CTD in evolution.
At the start of this project, we had anticipated that the divergent repeats would be essential for fly viability since the sequences of the CTD in 12 species of Drosophila are highly conserved. Moreover, the ratio of synonymous to nonsynonymous mutations among the 12 species is high, thus indicating that the fly CTD is under significant purifying selection. Our discovery that the entire fly CTD can be replaced by consensus repeats poses a conundrum since it calls into question the basis for the purifying selection. One possibility is that intra and intermolecular interactions intrinsic to the CTD constrain the sequence. Our SAXs analysis showed that the CTD adopts a compact random coil structure and the compaction implies transient interactions within the CTD (Gibbs et al., 2017). Also, recent results show that CTD molecules self-associate to form droplets (Boehning et al., 2018). Both types of interactions might need to be finely tuned to prevent aggregation and allow for factor binding to the CTD. Despite the sequence differences, the 29 consensus and wild-type Drosophila CTDs appear to meet these constraints.
The theory of constructive neutral evolution provides another answer to the conundrum. This theory was formulated to explain the gap between the simplicity and complexity of molecular machines that serve the same function in different organisms (Gray et al., 2010). In our case, the 29 consensus CTD represents the simple end of the spectrum while the Drosophila CTD represents the complex end. For both to be functionally equivalent, we posit that the essential functions of the CTD are mediated by a limited number of proteins that interact with the consensus repeats. It is notable that while the Drosophila CTD only contains two repeats that exactly match the consensus, these are embedded in a region encompassing seven other near-consensus motifs. Deletion of this region completely eliminated the capacity of Rpb1 to support fly viability. Constructive neutral evolution posits that chance mutations in the consensus sequences are tolerated by the fortuitous binding of other proteins which compensate for the loss of the consensus sequences. This fortuitous interaction might offset the tendency of the mutation to cause the CTD to aggregate or become mistargeted. Alternatively, the loss of consensus repeat could diminish the affinity of an essential protein for the CTD but this could be offset by the fortuitous binding of another protein that simultaneously associates with the mutant repeat and the protein that normally binds directly to the consensus repeat. Occurrence of the fortuitous binding caused by one mutation sets the stage for mutating additional consensus repeats. The evolutionary trajectory is then dictated by chance and could explain why the consensus CTD, the human CTD, and the Drosophila CTD all support Rpb1 function in the fly. If the consensus CTD truly represents the simplest CTD, then there should exist a class of mutations in genes outside of the Rpb1 gene that are deleterious to wild-type flies but not to those carrying the 29 consensus CTD.
Considering the number of CTD mutants that we had to generate to come to our final conclusion, we feel extremely lucky to have started this project in flies: the fruit fly is indeed an amazing model system to study the CTD. In addition to this work, the divergent nature of the fly CTD has allowed our collaborators Dr. Scott Showalter at PSU, Drs. Yan “Jessie” Zhang and Jennifer Brodbelt to map structural changes and post-translational modifications to each individual repeat without having to introduce additional mutations to the CTD (Gibbs et al., 2017; Mayfield et al., 2016). Furthermore, the fly salivary gland present a unique system to characterize the spatial distribution of transcription factors with just a standard confocal microscope. We envision that the currently available gene-editing and optogenetic tools will allow more exciting discoveries of transcription to be made in the salivary glands.
One of the things I learned during my stay in another lab is that you can’t succeed completely by yourself. You can try, sure, but a relevant unanswered question in biology is a hard nut to crack. It usually takes a multidisciplinary approach and great ideas coming from more than one brain. Collaborations are key to overcome this issue.
Most of the times, what keeps you from collaboratingwith peer scientists is money. Money to travel and money to pay for your accommodation. Renting a room in the US can cost as much as your entire monthly wage in Spain. Time is another big factor. Traveling takes time. As a Ph.D. student though, my time is fully dedicated to research anyway, so that factor is negligible for me.
Collaborate or perish
When Kate McDole and colleagues published their paper (McDole et al., 2018) describing their innovative live imaging microscope, I thought, ‘That is a method I could use for my project’. However, in my research center, I don’t have the facilities nor the expertise to carry it out.
In Miguel Torres Lab, at the Spanish Center for Cardiovascular Research, I am using live imaging to study how the heart establishes its heterogeneous cell types during gastrulation and how mouse embryos manage to form a primitive tube that later on starts beating. However, our methodology does not support imaging live specimens for longer than 20h. Keller’s Lab, at Janelia Reseach Institute, managed to build a fast light-sheet microscope that adapts to the mouse embryo growth and optical properties, allowing its imaging throughout 48h at an unprecedented resolution.
A travel grant to make it happen
The Company of Biologists provided with the funds to cover my expenses, and so I was able to join Keller’s lab for 5 weeks. At Janelia, I found a welcoming and collaborative environment surrounded by cutting-edge facilities. Besides getting some precious datasets from their microscope, I learned how to analyze the vast amount of data generated by time-lapse acquisitions (they run for 48h and take 300 stacks every 4 minutes, do the math…). Learning to program in Python wasn’t in my plans but it ended up being the most valuable thing I learned during the stay. Programming was completely unknown to me before I did the stay, and now I don’t know what I would do without it. To be able to quantify the cell dynamics I am observing in the videos and extract meaningful information becomes way more efficient when one uses programming.
It’s true that a Skype call can help you work together with other scientists, but sometimes you need to be there to learn other techniques, bring up together a project or receive inputs from clear-eyed researchers. That, I think, makes a project more solid and likely to succeed in answering the proposed question. I strongly believe collaborations blossom science, and the role of these traveling fellowships is key to make them happen. At the end of the day, to get to your goal you need a team, and some members of that dream team might be in another part of the world.
If you want to know a bit more about my Ph.D. project, here’s a 4min video explaining it:
The Craft lab at Boston Children’s Hospital and Harvard Medical School is looking for an outstanding, highly motivated postdoctoral fellow to join our developmental biology and pluripotent stem cell-based team.
We established directed differentiation protocols to generate distinct articular and growth plate-like cartilage lineages from human and mouse pluripotent stem cells (Craft et al., Development 2013; Craft et al., Nature Biotech 2015). NIH and foundation funded projects in our lab include investigating mechanisms of articular chondrocyte lineage commitment and stability of this fate through deep sequencing and functional assays, specification and characterization of joint progenitor cells using newly generated fluorescent reporter lines, translational/preclinical experiments of cartilage repair in large animals, and developing iPSC models of congenital cartilage disease. Collaborative projects with Harvard faculty include studies of how modifications in regulatory elements of GDF5 impact gene expression and differentiation of joint lineages (with Dr. Terence Capellini, Harvard University), and evaluating the cartilage and tendon-inducing functions of small molecules and their associated signaling pathways (identified by Dr. Jenna Galloway, Massachusetts General Hospital, through zebrafish screens) in embryonic stem cell (ESC) differentiation cultures.
Requirements: The successful candidate will have received a PhD or MD/PhD within the past 2 years, a minimum of 3 years laboratory experience including tissue culture, at least one first author publication, and excellent communication skills in English. Individuals with experience in one or more of the following are encouraged to apply: limb/joint/cartilage or early embryonic development, signal transduction pathways, ESC/iPSCs, single cell RNAseq/programming.
Apply: Interested applicants should email their CV, a cover letter describing their research background/interests, and contact information for three references to april.craft at childrens.harvard.edu
Press release from Development. You can also read the Research Highlight for this article.
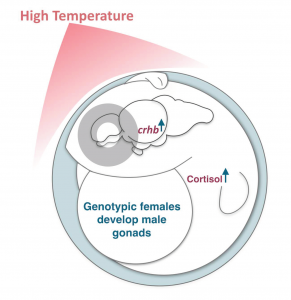
Researchers have identified the hormone that causes sex reversal of medaka fish growing in high temperatures. This study from the Instituto Tecnologico de Chascomus (INTECH) in Argentina is the first to report that the brain is involved in the masculinization of females, which has implications for fish populations as temperatures rise. The research has just been published in the scientific journal, Development.
Dr Juan Fernandino led the team investigating sex reversal, a phenomenon seen in some species whereby environmental conditions, such as high temperatures, cause genetically female fish to develop testes rather than ovaries. Now, researchers show for the first time that the brain can influence this process. “In the past, sex determination studies were specially focused on the gonad, probably because in genetic sex-determining genes are initially active in the gonad, driving the development of the testis,” explains Fernandino, “I was surprised at how long it took us to change our focus that sexual determination begins exclusively in the gonad.”
A schematic representation of the proposed mechanism for sex reversal in medaka fish. Credit. Diana Castañeda
After identifying the hormone produced in fish growing in high temperatures, the researchers used CRISPR/Cas9 genome-editing technology to block the ability of cells to detect it. Importantly, this stopped genetic females developing into males, “I was happily surprised that using gene editing we obtained a complete suppression of masculinization induced by temperature” says Fernandino, “our results highlighted for the first time the participation of the brain as a transducer of environmental stressors, directing development of the testis in genotypic females.”
Identifying how sex reversal occurs has important implications for how some species of fish reproduce. Increasing temperatures and warmer waters might result in more male fish and fewer females. Not only would this affect the ability of fish to reproduce, but also the repercussions on fish populations might have knock-on effects for the ecosystem and fishing industry. “Understanding the molecular mechanisms behind heat-induced masculinization is of great importance for controlling sex ratios in aquaculture and to predict the potential effects of climate change in an important group of animals,” adds Fernandino.
The Maehr Laboratory is seeking a highly motivated postdoctoral research fellow to investigate the molecular basis of immune syndromes and immune cell development.
The applicant will be taking on experimental and/or computational projects in a collaborative cross-disciplinary group. Envisioned projects integrate pluripotent stem cell differentiation approaches with applied developmental immunology approaches and functional genomics, to decipher the molecular underpinnings of human immune syndromes such as autoimmunity and immunodeficiency. In addition, the collaborative project will apply disease models, single-cell omics, and computational analysis strategies based on data-integration and machine learning.
The dynamic and collaborative Maehr lab is embedded in the cutting edge research environment of the Program in Molecular Medicine and Diabetes Center Of Excellence at the University of Massachusetts Medical School. Please visit our webpage for full list of affiliations and more information about us: http://maehrlab.net/ (web) and @MaehrLab (twitter)
Candidates should possess a Ph.D. and have a strong background in immunology, developmental and/or computational biology. Experience with stem cell differentiation, bioengineering and/or computational approaches is desirable. Excellent communication, writing, and collaboration skills are essential.
Interested candidates should email a cover letter and CV to Dr. René Maehr (rene.maehr@umassmed.edu).
In development and during regeneration in adults, muscle fibres develop from muscle progenitor cells, and the proliferation, differentiation and fusion of these progenitors needs to be tightly controlled and co-ordinated. A new paper in Development studies the role of the mTOR protein in this process using genetic deletions that target either of the two protein complexes mTOR functions in. We caught up with lead author Nathalie Rion and her supervisor Markus Rüegg, Professor of Neurobiology at Biozentrum, University of Basel, to find out more about the work.
Markus, can you give us your scientific biography and the questions your lab is trying to answer?
MR Even as a young teenager I was fascinated to learn about discoveries of the molecular mechanisms underlying the formation and function of the nervous system. During my PhD studies at the University of Zurich, I characterized cell adhesion molecules that are important for axonal pathfinding, and during my postdoctoral work at Stanford University I cloned and functionally characterized splice isoforms of the protein agrin, the key inducer of neuromuscular synapses. For more than 25 years since then I have been a group leader at the Biozentrum, University of Basel.
My laboratory has also evolved during these 25 years. From initially studying mechanisms underlying neuromuscular synapse formation, we are now interested in how skeletal muscle can adapt to changes in the environment. It is well known that skeletal muscle, which makes up to 50% of the entire body weight, adapts rapidly to changes. For example, if muscle is not properly innervated, it loses muscle mass. Similarly, at old age, muscle mass is lost in a syndrome called sarcopenia. However, the molecular mechanisms driving these adaptive processes are not well understood, and we are trying to better understand them by using mice and tissue culture experiments. In particular, we discovered that changing signalling of the protein mammalian target of rapamycin (mTOR) in skeletal muscle affects the development of sarcopenia in mice. Another interest in the laboratory is to try to develop treatment options for a very severe, rare congenital muscular dystrophy, called MDC1A or LAMA2 MD. In this disease, the muscle fibres cannot withstand the mechanical load that occurs during contraction and, as a result, they degenerate. Over the years, we have developed small linker proteins that are capable of stabilizing the muscle fibres and thereby counteracting this severe muscular dystrophy in mice.
In summary, the common theme of the laboratory is the attempt to understand pathological states of skeletal muscle and its innervation by motor neurons, and to develop approaches to counteract them.
Nathalie: how did you come to join the Rüegg lab, and what drives your research?
NR As a young student, I did not have a defined plan for my future and rather followed my curiosity and my fascination with nature and what is still to be discovered. I was an undergraduate at Biozentrum, and like all students there, I was introduced to all the research groups of the institute. The course of Markus and his group instinctively drew my attention and subsequently formed my future career. He is extremely motivated and engaged in supporting and educating students. I was very fortunate to have the possibility to learn, work and develop in his lab, not only professionally but also on a personal level. My research was also greatly driven by my mentor and supervisor, Dr Perrine Castets. She is an extremely talented, hard-working and dedicated researcher and teacher who inspired and motivated me during the past 8 years. We worked very closely together in Markus’s lab and supported each other in our projects. Therefore, I would like to emphasize her great contribution to this work and thank her for all her support.
Prior to your work, what was known about the role of mTOR-and the relative roles of its mTORC1 and mTORC2 complexes in muscle development?
NR & MR Published work mainly investigated the role of mTORC1 and mTORC2 with shRNA-mediated knockdown experiments in cultured myoblasts from a mouse cell line (C2C12). These results suggested that mTORC1 had a role in muscle differentiation, but there were others who provided evidence that this function was largely mTORC1-independent and mTORC2-dependent. Another important piece of work showed that activation of mTORC1 was involved in the ‘priming’ of quiescent satellite cells for activation. This work, however, did not study its role in regeneration or in myogenesis. In addition, previous work from our laboratory using Cre-drivers specific for skeletal muscle fibres showed that depletion of raptor (an essential component of the mTORC1 complex) but not of rictor (an essential component of the mTORC2 complex) during muscle growth led to myopathy. It was thus important to understand the role of raptor and rictor during myogenesis and during regeneration in the adult muscle.
Can you give us the key results of the paper in a paragraph?
NR & MR By depleting raptor or rictor in embryonic muscle progenitors, we show that mTORC1, but not mTORC2, deficiency in developing muscle impairs embryonic myogenesis without completely abolishing it. Inactivation of mTORC1 in muscle stem cells impairs injury-induced regeneration of the adult tissue due to a delay of activation and commitment into the myogenic lineage, as well as proliferation deficits. In vitro, raptor depletion in myoblasts slows down proliferation, differentiation and fusion. Nevertheless, muscle progenitors deficient of mTORC1 signalling contribute to the formation of skeletal muscle.
Control (L) and raptor-Myf5-knockout (R) E13.5 mouse embryos, with embMHC in red, laminin in green and DAPI in blue.
Do you think mTORC1 is doing the same thing in embryonic and regenerating adult muscle?
NR & MR This is a difficult but very interesting question. From our work, one can conclude that the cell-intrinsic function of raptor in the embryonic and adult stages is similar. Proliferation of raptor-depleted cells, isolated from embryonic or adult skeletal muscle, is strongly impaired, and of course proliferation is an important process in embryonic myogenesis and adult muscle regeneration. However, the extracellular environment is quite different between embryos and adult muscle. Hence, differences in extrinsic factors could also affect the dependence of satellite cells on mTORC1 signalling.
How do you think muscle progenitors that lack mTORC1 are able to form myofibres?
NR & MR Removal of raptor does not completely abolish protein synthesis. Hence, a low, basal level of protein synthesis may suffice to allow some regeneration and myofibre formation in the absence of mTORC1 signalling. Thus, non-mTORC1-dependent pathways, such as the Mnk1/Mink2 kinases, could become active. It would be interesting to investigate whether there is such a compensatory mechanism and to investigate the possible reasons for this.
When doing the research, did you have any particular result or eureka moment that has stuck with you?
NR Thinking back, I can still remember the first time I performed FACS sorting from embryonic muscle tissue as one of my biggest eureka moments. It was so fascinating and exciting for me to see that it is possible to isolate, analyze and culture living cells from such a tiny amount of tissue, and that the cells survive such a long and harsh procedure.
And what about the flipside: any moments of frustration or despair?
NR Our embryonic work required us to set up timed-matings before the actual experiment could be initiated. Therefore, the most challenging part of this project was to deal with the long timelines and associated limitations, including the small amount of harvested muscle tissue arising from our Rptor knockout mouse model. My biggest despair was when, after these long planning phases, the females were not gestant, no Rptor knockout embryos were among the offspring, or an experiment just failed. However, I learnt from such unfortunate events, and still had a great time working with mouse models: they allowed me to analyze the role of mTOR in myogenesis at both an embryonic and an adult stage.
So what next for you after this paper?
NR I highly enjoyed doing laboratory research for the past 8 years, and I was very fortunate to work on such a fascinating project. After completing my PhD studies and this project, I wanted to change my point of view in science and learn about the journey of medicines from research and development in pre-clinical and clinical studies towards marketing authorization. Today, I am proud to be an employee at SFL Regulatory Affairs & Scientific Communication GmbH, here in Basel, in the field of regulatory affairs. I am very happy to implement my knowledge and experience from research, scientific writing and project management in the support we provide to clients, ranging from regulatory strategy and advice on the development of healthcare products to hands-on operational support during product registration and postmarketing activities.
Where will this work take the Rüegg lab?
MR The work now published in Development was the first time we looked at myogenesis per se and examined the function of satellite cells (the adult muscle stem cells). Satellite cells are also the cells that allow muscle fibres to fully regenerate after an injury, and we found that mTORC1 signalling is important for the regenerative process. Now, we would like to study the molecular processes that are involved in muscle regeneration in general, with the aim of understanding why regeneration is often hampered in the context of muscular dystrophies. Although muscular dystrophies are primarily muscle-degenerative diseases, successful regeneration of muscle is of fundamental importance to overcome some of these degenerative processes. Recent evidence indicates that in many muscular dystrophies, including MDC1A/LAMA2 MD, muscle regeneration is severely impaired.
For me, cooking and baking are quite similar to performing an experiment
Finally, let’s move outside the lab – what do you like to do in your spare time in Basel?
NR I play the piccolo (a special flute) at the Carnival of Basel, which in 2017 was added to UNESCO’s list of Intangible Cultural Heritage. Between February and March each year, Basel transforms into a large, jovial festival with costumed carnivalists, concerts and parades of fife and drum cliques, lanterns and wagons for 72 h.
Besides playing music and enjoying sports, I am a passionate baker and cook, especially in the company of my family and friends. For me, cooking and baking are quite similar to performing an experiment. Both procedures are based on the following of a ‘recipe’ with described components and processes. However, the understanding and feeling for the individual steps and ingredients determine the success of the outcome.
MR As I mainly sit at my desk during work, I need to get some physical challenges. I therefore enjoy all kinds of sports activities, my favourites being jogging, bicycling, hiking in the summer and skiing in the winter.
Department of Biology at the Faculty of Science, University of Copenhagen is offering a Postdoc position in Physiology and Genetics commencing 1 August 2019 or as soon as possible thereafter.
Description of the scientific environment
The Andersen-Colombani group at the Department of Biology, University of Copenhagen, is recruiting a Postdoc to work on a 3-year project aiming at understanding the mechanisms controlling intestinal stem cell proliferation and gut homeostasis and pathologies using Drosophila Melanogaster as a model organism. The position is available to start on 1 August 2019 or as soon as possible thereafter. The group is housed in the section of Cell and Neurobiology at the Department of Biology (https://www.biocenter.ku.dk/) and affiliated with the Novo Nordisk Center for Stem Cell Biology, DanStem (https://danstem.ku.dk/).
Background
The intestine, which represents one of the largest interfaces with the external environment, plays a key role in relaying environmental inputs to other organs to produce systemic responses. In turn, the gut is subject to multiple regulatory inputs from the brain, muscles, liver and adipose tissues. At steady-state turnover rates, the human intestine undergoes complete self-renewal every 4-5 days, a process which is highly accelerated in response to damage of the gut epithelium. This capacity for self-renewal relies on the proliferative activity of the intestinal stem cells (ISCs), which is tightly controlled by multiple local and systemic signals released from neighboring cell populations (the ISC niche) and non-gastrointestinal organs. Despite the physiological divergence between insects and mammals, studies have shown that flies represent a model that is well suited for studying stem cell physiology during ageing, stress, and infection. Our team is interested in identifying the intra- and inter-organ couplings contributing to gut homeostasis and disease
Project Description
The project advertised here aims at identifying local signals controlling intestinal stem cell proliferation and gut homeostasis. For this purpose, RNAis will be used to known down all genes encoding secreted peptides specifically in the stem cell niche. Sensitivity to oral infection with the gram-negative bacteria, Ecc15, will be used as readout to identify niche-derived signals required for ISC-driven intestinal regenerative growth. The potential of the identified signals to control ISC proliferation during homeostasis and disease will be studied. Identifying paracrine stress signals required for ISC-dependent tissue self-renewal is of importance, since the same signals tend to initiate colorectal cancers in predisposed individuals. Since large-scale functional approaches are not feasible in vertebrate, this project could reveal novel couplings contributing to mammalian gastrointestinal homeostasis and disease. The postdoc’s duties will include working on the project described here as well as teaching.
We are looking for highly motivated individuals with a PhD degree in Biology and with prior experience in fly physiology and genetics.
Qualifications/Selection criteria
Applicants should hold a PhD degree in Biology
Having prior experience with the Drosophila model and Genetics is essential
Having a solid background in Physiology is an advantage
Strong motivation and very good scientific skills are essential
Good communication skills, oral and written
Terms of employment
The position is covered by the Memorandum on Job Structure for Academic Staff.
Terms of appointment and payment accord to the agreement between the Ministry of Finance and The Danish Confederation of Professional Associations on Academics in the State.
The starting salary is currently up to DKK 430.570 including annual supplement (+ pension up to DKK 73.627). Negotiation for salary supplement is possible.
The application, in English, must be submitted electronically by clicking APPLY NOW below.
Please include
Curriculum vita
Diplomas (Master and PhD degree or equivalent)
Research plan – description of current and future research plans
Complete publication list
Separate reprints of 3 particularly relevant papers
The deadline for applications is Tuesday 23 April 2019, 23:59 GMT +2.
After the expiry of the deadline for applications, the authorized recruitment manager selects applicants for assessment on the advice of the Interview Committee.
The University wishes our staff to reflect the diversity of society and thus welcomes applications from all qualified candidates regardless of personal background.
 (No Ratings Yet)
(No Ratings Yet) (15 votes)
(15 votes)
 (2 votes)
(2 votes)