POSTDOCTORAL POSITION is immediately available to follow up on our recent published work identifying that lymphatic-secreted signals are required for organ growth and repair using a variety of available mouse models (https://www.nature.com/articles/s41586-020-2998-x). Highly motivated individuals who recently obtained a PhD. or MD degree and have a strong background in mouse cardiovascular, molecular and developmental biology are encouraged to apply. Interested individuals should send their curriculum vitae, a brief description of their research interests, and the names of three references to:
Guillermo Oliver, Ph.D
Thomas D Spies Professor of Lymphatic Metabolism
Director Center for Vascular and Developmental Biology
Northwestern University Feinberg School of Medicine
Northwestern University is an Equal Opportunity, Affirmative Action Employer of all protected classes, including veterans and individuals with disabilities. Women, underrepresented racial and ethnic minorities, individuals with disabilities, and veterans are encouraged to apply. Hiring is contingent upon eligibility to work in the United States.
A position is available for a Post Doctoral Associate 1 in the Ameen Lab, within the Section of Digestive Diseases at Yale School of Medicine. Dr. Ameen is a pediatric gastroenterologist and physician scientist. She is Professor Pediatrics, Cellular and Molecular Physiology, and has been investigating CFTR in the intestine for over 25 years. Collaborative studies are ongoing with Dr. Kaelyn Sumigray in the Department of Genetics. This is a wonderful opportunity to learn complementary skills from the laboratory of a physician-scientist and basic stem cell geneticist.
A major goal of the Ameen laboratory is understanding the regulation of CFTR chloride channels in the native intestine and its relevance to intestinal diseases. Studies are predominantly conducted in animal models, including normal rat, transgenic mouse models of diseases where CFTR is implicated. We also conduct studies in human intestinal organoids and cultured human intestinal cells. Studies in the Sumigray lab focus on intestinal stem cell biology. Collaborative project with the Sumigray lab is using single cell RNA-sequencing and cutting edge techniques to examine and characterize a unique enterocyte subtype in the intestine and its relevance to Cystic Fibrosis. Yale is a very collaborative environment with many opportunities to interact with scientists and physician-scientists on campus.
Techniques used in the laboratory include: PCR, qPCR, tail clipping, animal husbandry, cell and organoid culture, Ussing Chamber electrophysiology in tissues and cells, immunofluorescence staining of cells and tissues, fluorescence and confocal microscopy imaging, immunoblot analysis of rodent tissue lysates, co-immunoprecipitation using tissue lysates, Ussing chamber electrophysiology.
Interested candidates please contact the PI: Nadia Ameen, Professor Pediatrics, Cellular and Molecular Physiology at Nadia.Ameen@yale.edu
In July 2021, a Workshop from The Company of Biologists will bring together experts in human cell fate engineering and experts in the manipulation and characterization of single cells, with the goal of understanding the molecular steps underlying the transformation of one cellular identity into another. The Workshop is organised by Marisa Karow, Samantha Morris and Barbara Treutlein and will be held in Buxted Park in East Sussex.
There are 10 funded places for early-career researchers to attend this Workshop along with the 20 speakers – a fantastic opportunity to interact with the leaders in the field (see the speaker list below.
The application deadline for this Workshop is 16 December 2020.
preLights is a preprint highlighting service that is centred around a community of early-career researchers. Launched in 2018, this initiative has gained significant attention from researchers as well as the publishing industry, being nominated for an ALPSP Award for Innovation in Publishing in 2019. As preLights nears its 1,000th post, we are looking for the right person to join us for the next phase of community building and the site’s growth and development.
Joining an experienced and successful publishing team, this is an exciting opportunity for an enthusiastic and motivated team player to take a step into publishing or for someone already working in publishing to extend their interest in online communities. Please see the full job description for further details.
To apply, or for more information, contact recruitment@biologists.com. Applications should be received by 31 December 2020, although late applications may be considered. Interviews will be virtual and are expected take place in January. Applicants should be eligible to work in the UK.
preLights, a preprint highlighting service that is centred around a team of early-career researchers, is looking for a new Community Manager.
Launched in 2018, preLights has gained significant attention from researchers as well as the publishing industry, being nominated for an ALPSP Award for Innovation in Publishing in 2019. As we near our 1,000th post, we are looking for the right person to join us for the next phase of community building and the site’s growth and development.
Joining an experienced and successful publishing team, this is an exciting opportunity for an enthusiastic and motivated team player to take a step into publishing or for someone already working in publishing to extend their interest in online communities. Please see the full job description for further details.
To apply, or for more information, contact recruitment@biologists.com. Applications should be received by 31 December 2020, although late applications may be considered. Interviews will be virtual and are expected to take place in January.
The Company of Biologists is also looking for a Features and Reviews Editor for the journal Disease Models & Mechanisms, see the job description here.
We at the Node and Development were greatly saddened to hear about the recent death of Kathryn Anderson, Professor & Chair of the Developmental Biology Program at the Memorial Sloan Kettering Cancer Center in New York,. Kathryn’s lab worked on the genetic pathways directing embryonic patterning and morphogenesis in the mouse embryo.
We also point readers towards this tribute from Tamara Caspary on Twitter – the dozens of replies show how highly the community thought of Kathryn.
I am heartbroken that Kathryn Anderson passed away this morning- at home and in her sleep. My thoughts are with her husband, her lab members and all my colleagues who loved her as I did. 1/n
— Tamara Caspary (she, her, hers) (@TamaraGenes) December 1, 2020
I was lucky enough to interview Kathryn for Development in 2016 – the year she won the SDB Edwin Conklin Medal. She was a generous interviewee and I can still remember sitting in Boston with her, bonding over our shared love of Radiohead!
Earlier in the year, we were similarly saddened to learn about the death of José Luis Gómez-Skarmeta, PI at the Centro Andaluz de Biología del Desarrollo in Seville who worked on vertebrate gene regulation, evolution and morphogenesis.
At the recent SEBD virtual meeting, a session on gene regulation was held to honour José Luis, and the society also announced the launch of the ‘José Luis Gómez-Skarmeta Awards for scientific excellence in Developmental Biology’. This year, the award were won by Manuel Irimia and Alvaro Rada-Iglesias.
Exciting PhD project supervised by Dr Sanchez-Soriano and Dr Alison Twelvetrees through the MRC DiMeN Doctoral Training Partnership on mechanisms of synapse loss during neurodegeneration and ageing.
The aim of this studentship is to understand the processes of ageing and neurodegeneration, through the study of mechanisms of synaptic loss. You will be part of a multidisciplinary collaboration between two experienced groups at the Institute of Systems, Molecular & Integrative Biology (ISMIB, University of Liverpool) and the Sheffield Institute for Translational Neuroscience (SITraN, University of Sheffield).
Nerve cells are organised into complex neuronal networks, wiring the body or brain regions over distances up to a meter away in humans. For this, neurons extend long and thin processes called axons. At the tip of these axons, neurons establish synapses, specialised neuronal cell junctions which contain complex machinery for rapid transmission of signals to partner cells. The maintenance of this synaptic machinery fails during ageing and in disease, and the resulting synaptic malfunction is an important cause for cognitive, sensory and motor decline. Maintaining synapses requires transport of synaptic proteins from the cell body to the distant synapses up to a meter away. The Jun-Kinase (JNK) signalling pathway is a key regulator of this process. Importantly, physiological changes such as oxidative stress typically occurring during ageing and neurodegeneration, alter JNK activation patterns. The goal of this project is to understand how the JNK pathway regulates the transport and precise delivery of synaptic components and how it links to synapse loss occurring during ageing and disease.
This studentship represents a unique opportunity to integrate in vivo models of ageing and neurodegeneration capitalising on the brain of the fruit fly Drosophila as a highly efficient model, together with mouse and rat neuronal models and in vitro reconstitutions assays. Using these systems, you will study the role of JNK during the regulation of intracellular transport and synaptic decay. You will receive training by the two supervisory groups in neuronal cell biology (fly neurons in culture and in vivo in the adult Drosophila brain, primary neuronal culture from mouse and rat), in genetic strategies, in quantitative live imaging of cultured neurons and whole tissue, in analytical methods, techniques required for in vitro reconstitution of transport assays with complementary quantitative analysis. Understanding the causes of synapse decay during ageing or disease is crucial to providing new avenues for therapeutic intervention.
Creative individuals with an eye for detail are encouraged to apply. The successful applicant will be based in the Institute of Systems, Molecular & Integrative Biology supervised by Dr Sánchez-Soriano (sanchezlab.wordpress.com/research), whilst working closely with the SITraN lab, Department of Neuroscience in Sheffield under the supervision of Dr Alison Twelvetrees (www.twelvetreeslab.co.uk). Applications from candidates, ideally with some background in cell biology, genetics, neuroscience and/or biomedical sciences are welcome. Interested applicants should contact Dr Sanchez-Soriano to discuss the project: n.sanchez-soriano@liverpool.ac.uk.
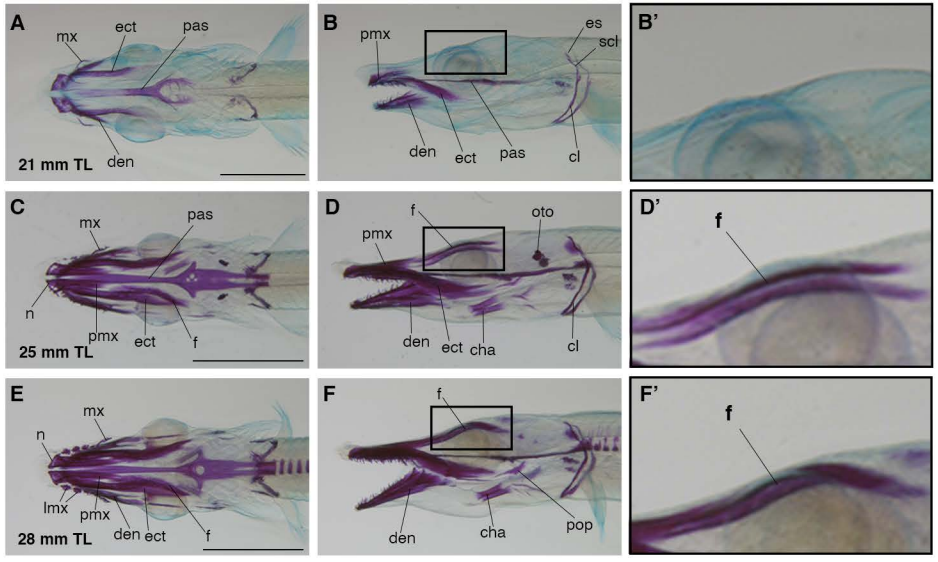
Hand2 delineates mesothelium progenitors and is reactivated in mesothelioma
Karin D. Prummel, Helena L. Crowell, Susan Nieuwenhuize, Eline C. Brombacher, Stephan Daetwyler, Charlotte Soneson, Jelena Kresoja-Rakic, Manuel Ronner, Agnese Kocere, Alexander Ernst, Zahra Labbaf, David E. Clouthier, Anthony B. Firulli, Héctor Sánchez-Iranzo, Rebecca O’Rourke, Erez Raz, Nadia Mercader, Alexa Burger, Emanuela Felley-Bosco, Jan Huisken, Mark D. Robinson, Christian Mosimann
CSF1R-dependent macrophages control postnatal somatic growth and organ maturation
Sahar Keshvari, Melanie Caruso, Lena Batoon, Anuj Sehgal, Ngari Teakle, Omkar L. Patkar, Cameron E. Snell, Chen Chen, Alex Stevenson, Felicity M. Davis, Stephen J. Bush, Clare Pridans, Kim M. Summers, Allison R. Pettit, Katharine M. Irvine, David A. Hume
A gene desert required for regulatory control of pleiotropic Shox2 expression and embryonic survival
Samuel Abassah-Oppong, Brandon J. Mannion, Virginie Tissières, Eddie Rodríguez-Carballo, Anja Ljubojevic, Fabrice Darbellay, Tabitha A. Festa, Carly S. Sullivan, Guy Kelman, Riana D. Hunter, Catherine S. Novak, Ingrid Plajzer-Frick, Stella Tran, Jennifer A. Akiyama, Iros Barozzi, Guillaume Andrey, Javier Lopez-Rios, Diane E. Dickel, Axel Visel, Len A. Pennacchio, John Cobb, Marco Osterwalder
Transcriptional Network Orchestrating Regional Patterning of Cortical Progenitors
Athéna R Ypsilanti, Kartik Pattabiraman, Rinaldo Catta-Preta, Olga Golonzhka, Susan Lindtner, Ke Tang, Ian Jones, Armen Abnousi, Ivan Juric, Ming Hu, Yin Shen, Diane E Dickel, Axel Visel, Len A Pennachio, Michael Hawrylycz, Carol Thompson, Hongkui Zeng, Iros Barozzi, Alex S Nord, John Rubenstein
Tracking H3K27me3 and H4K20me1 during XCI reveals similarities in enrichment dynamics
Sjoerd J. D. Tjalsma, Mayako Hori, Yuko Sato, Aurelie Bousard, Akito Ohi, Ana Cláudia Raposo, Julia Roensch, Agnes Le Saux, Jumpei Nogami, Kazumitsu Maehara, Tomoya Kujirai, Tetsuya Handa, Sandra Bagés-Arnal, Yasuyuki Ohkawa, Hitoshi Kurumizaka, Simão Teixeira da Rocha, Jan J. Żylicz, Hiroshi Kimura, Edith Heard
Aberrant gliogenesis and excitation in MEF2C autism patient hiPSC-neurons and cerebral organoids
Dorit Trudler, Swagata Ghatak, James Parker, Maria Talantova, Titas Grabauskas, Sarah Moore Noveral, Mayu Teranaka, Melissa Luevanos, Nima Dolatabadi, Clare Bakker, Kevin Lopez, Abdullah Sultan, Agnes Chan, Yongwook Choi, Riki Kawaguchi, Nicholas Schork, Pawel Stankiewicz, Ivan Garcia-Bassets, Piotr Kozbial, Michael G. Rosenfeld, Nobuki Nakanishi, Daniel H. Geschwind, Shing Fai Chan, Rajesh Ambasudhan, Stuart A. Lipton
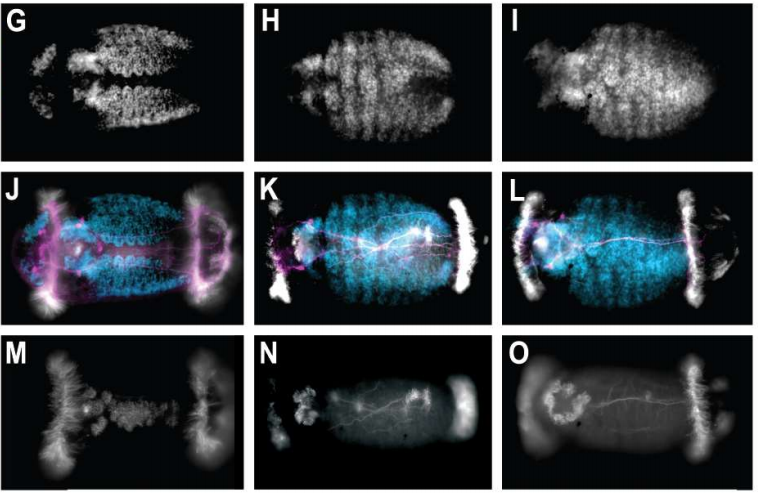
Highly multiplexed spatially resolved gene expression profiling of mouse organogenesis
T. Lohoff, S. Ghazanfar, A. Missarova, N. Koulena, N. Pierson, J.A. Griffiths, E.S. Bardot, C.-H.L. Eng, R.C.V. Tyser, R. Argelaguet, C. Guibentif, S. Srinivas, J. Briscoe, B.D. Simons, A.-K. Hadjantonakis, B. Göttgens, W. Reik, J. Nichols, L. Cai, J.C. Marioni
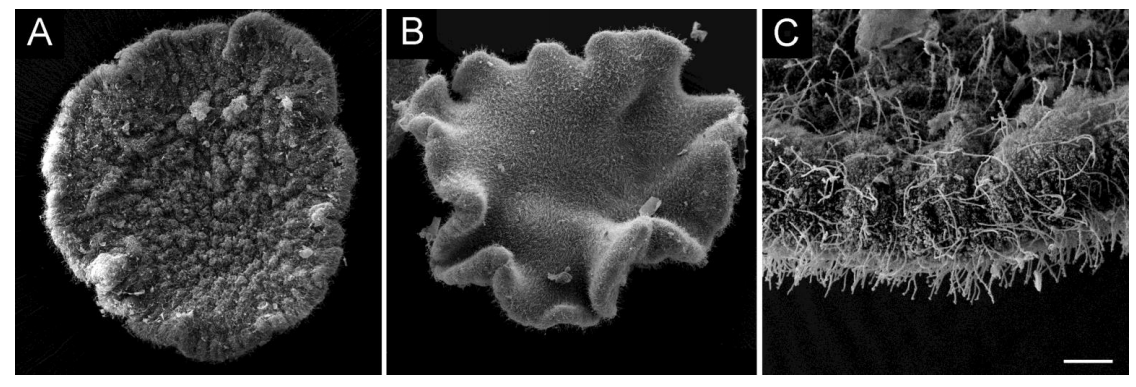
Isotropic 3D electron microscopy reference library of whole cells and tissues
C. Shan Xu, Song Pang, Gleb Shtengel, Andreas Müller, Alex T. Ritter, Huxley K. Hoffman, Shin-ya Takemura, Zhiyuan Lu, H. Amalia Pasolli, Nirmala Iyer, Jeeyun Chung, Davis Bennett, Aubrey V. Weigel, Tobias C. Walther, Robert V. Farese Jr., Schuyler B. van Engelenburg, Ira Mellman, Michele Solimena, Harald F. Hess
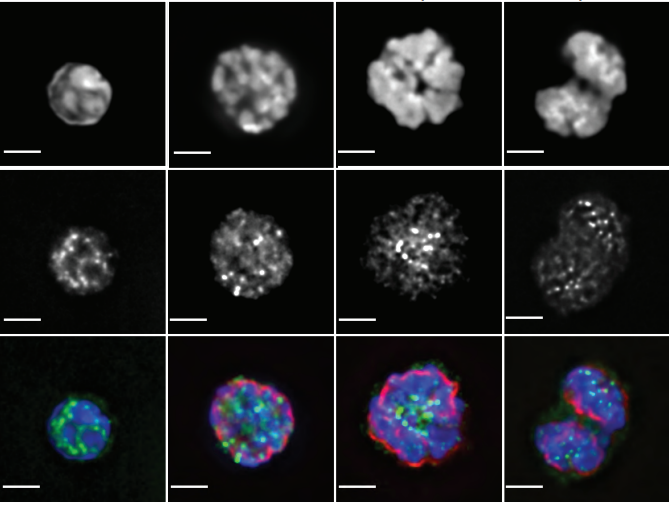
Far-red fluorescent genetically encoded calcium ion indicators
Rochelin Dalangin, Mikhail Drobizhev, Rosana S. Molina, Abhi Aggarwal, Ronak Patel, Ahmed S. Abdelfattah, Yufeng Zhao, Jiahui Wu, Kaspar Podgorski, Eric R. Schreiter, Thomas E. Hughes, Robert E. Campbell, Yi Shen
Yesterday we held the third webinar in our new series, this time focusing on plant development and chaired by Development Editor Yrjö Helariutta. Here you’ll find recordings of the talks and their live Q&A sessions moderated by Yrjö.
Marta Mendes (from Lucia Colombo’s lab at the University of Milan)
‘The RNA dependent DNA methylation pathway is required to restrict SPOROCYTELESS/NOZZLE expression to specify a single female germ cell precursor in Arabidopsis’
Marta’s paper is coming out soon in Development.
Krisztina Ötvös (from Eva Benková’s lab at IST Austria)
‘Modulation of root growth by nutrient-defined fine-tuning of polar auxin transport’
A postdoctoral position is available from January 2021 in the “Developmental Genetics” laboratory (http://gendev.ulb.ac.be/bellefroidlab/). Our lab studies cortical and sensory neurogenesis. To do this, we use mouse genetics and gain and loss-of-function experiments in Xenopus. We are interested in understanding how some transcription factors we have previously identified as key cell fate determinants in the developing cerebral cortex (Dmrt3 and Dmrt5) and in somatosensory ganglia (Prdm12) function in neuronal specification and to identify novel important regulators of neuronal diversification. The selected candidate will be in charge of testing the functional importance of one recently identified new putative regulator of neuronal diversification. To approach the function of this gene, he/she will use knock-out and conditional overexpressing transgenic mice as well as gain and loss of function experiments in the Xenopus model.
Location
The laboratory is located in the Biopark Charleroi Brussels South in the Institute of Biology and Molecular medicine (IBMM) of the University of Brussels. It is also part of the ULB Neuroscience Institute (UNI – http://uni.ulb.ac.be/groups/developmental-genetics/) that integrates and synergizes brain researchers of all neuroscience perspectives, from molecular and cellular neurobiology to systems / cognitive and clinical neurosciences.
Qualification and experience
We are looking for highly motivated candidates with a background in molecular and developmental neurobiology. Preference will be given to applicants with experience in mouse genetics and/or with the Xenopus model system. The position is opened for one year renewable up to 2 years, starting as January 2021. Fluency in English is mandatory. The ability to speak French is not required.
Application
Interested candidates should send a letter of motivation describing their past research experiences and full CV with the name and e-mail address of 2 references to Eric Bellefroid (ebellefr@ulb.ac.be)
Selected recent related publications:
1. Desiderio et al. (2019). Cell Reports 26, 3522-3536.
2. Desmaris et al. (2018). J. of Neurosci. 38, 9105-9121.
3. Young et al., (2017) PNAS, 114, E5599-E5607
4. De Clercq et al., (2016) Cerebral Cortex, 28, 493-509.
5. Thélie et al., (2015) Development 142, 3416-28.
 (No Ratings Yet)
(No Ratings Yet)


 (3 votes)
(3 votes)