November turned out to be a bumper month on the Node with posts on research (current and historical), meetings and new resources, as well as interviews and a meeting report. Plus some beautiful science-inspired art. Here are some of our highlights, as well as our pick of the best of the web this month.
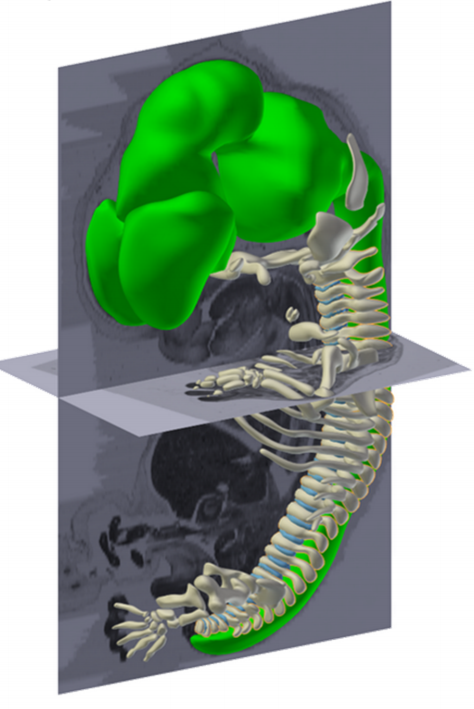
Happy Thanksgiving! Yes, we CT scanned our turkey, and like any good dinosaur biologist, prepared and accessioned the skeleton (OUVC 10789). pic.twitter.com/V2YbF6mF22
The first issue of @Nature was printed today in 1869. In opening article Huxley writes on obsolescence of theories but permanence of poetry. pic.twitter.com/rNXy5mbyAD
Staying underwater, Uwe Irion and colleagueslink heterotytpic, gap junction-mediated cell interactions with cell morphology during zebrafish skin patterning.
The JCS team featured Celeste Nelson as a Cell Scientist to Watch, whose lab is “focused on studying how groups of cells physically position or turn themselves into tissues.”
Nicholas Pilon and colleagues describe how a mouse line found in a screen for genes involved in neural crest development provides a model for Waardenburg syndrome type 4.
Colin Bingle and colleagues develop an in vitro model of the murine middle ear epithelium, recapitulating cell populations and protein production.
Our latest monthly trawl for developmental biology (and other cool) preprints. See June’s introductory post for background, and let us know if we missed anything
This month, we found preprints covering various aspects of plant growth and patterning, a lot of cell biology – including insights into microtubules organisation, RNA localisation and yeast size control – as well as a bunch of tools. One of the most talked about preprints of the month comes from our ‘Away from the bench‘ section: a guide for how to structure scientific papers. Happy reading!
MultiCellDS: a community-developed standard for curating microenvironment-dependent multicellular data. Samuel H. Friedman, Alexander R.A. Anderson, David M. Bortz, Alexander G. Fletcher, Hermann B. Frieboes, Ahmadreza Ghaffarizadeh, David Robert Grimes, Andrea Hawkins-Daarud, VStefan Hoehme, Edwin F. Juarez, Carl Kesselman, Roeland Merks, Shannon M. Mumenthaler, Paul K. Newton, Kerri-Ann Norton, Rishi Rawat, Russell C. Rockne, Daniel Ruderman, Jacob Scott, Suzanne S. Sindi, Jessica L. Sparks, Kristin Swanson, David B. Agus, Paul Macklin
The Monarch Initiative: Insights across species reveal human disease mechanisms. Christopher Mungall,Julie McMurry, Sebastian Koehler, James Balhoff, Charles Borromeo, Matthew Brush, Seth Carbon, TOM CONLIN, Nathan Dunn, Mark Engelstad, Erin Foster, Jean-Philippe Gourdine, Julius Jacobsen, Daniel Keith, Bryan Laraway, Suzanna Lewis, Jeremy Nguyen Xuan, eKent Shefchek, Nicole Vasilevsky, Zhou Yuan, Nicole Washington, Harry Hochheiser,Tudor Groza, Damian Smedley, Peter Robinson, Melissa Haendel
The Wellcome Trust – Medical Research Council Cambridge Stem Cell Institute is founded on the concept that deep understanding of stem cell biology will contribute to transforming future healthcare (http://www.stemcells.cam.ac.uk). In 2018 we will move into a new purpose built building adjacent to Addenbrooke’s Hospital and multiple research institutes – http://cambridge-biomedical.com/.
The Institute has openings for Group Leaders who will complement and synergise with our existing programmes. Areas of particular interest include:
i. The interface between physical, materials or engineering sciences and stem cell biology
ii. Cell and gene therapy
iii. Ageing of stem cells
Junior group leader candidates will have a minimum of 3 years post-doctoral experience, distinctive research achievements, and an original project proposal. Senior group leader candidates will be internationally recognised for independent high quality science and have an exceptional and well-founded research proposal.
The Institute offers a collegiate environment with excellent core facilities plus extensive opportunities to pursue basic and disease focussed studies. Successful candidates will be supported to obtain external personal fellowship and grant support within 1-2 years. Interim start-up packages may be available. Depending on experience, non-Clinicians can expect remuneration between £39,324 and £66,835.
To apply online for this vacancy and to view further information about the role, please visit: http://www.jobs.cam.ac.uk/job/12123. This will take you to the role on the University’s Job Opportunities pages. There you will need to click on the ‘Apply online’ button and register an account with the University’s Web Recruitment System (if you have not already) and log in before completing the online application form.
Applicants should upload a curriculum vitae (max 3 pages, to include date of PhD and details of any career gaps if applicable) with contact details of 3 referees, and a 1-2 page outline of your research proposal, by Sunday 29th January 2017.
Interviews will be held in April 2017. Please quote reference PS10734 on your application and in any correspondence about this vacancy.
The University values diversity and is committed to equality of opportunity. The University has a responsibility to ensure that all employees are eligible to live and work in the UK. Benefits include generous maternity/ paternity leave, flexible working and funds for returning carers and other family-friendly schemes.
Since the first reported results from Yamanaka et al. in 2006, pluripotent stem cell culture has become an advantageous approach for modeling human disorders and diseases. The directed differentiation of stem cells into particular cell types can also be the basis for powerful in vitro models of early developmental defects in humans. Our lab is interested in neural tube closure as well as neural crest cell development, and to investigate our questions we use the mouse as our model system. However, we also aim to translate our findings to human development. Thus, my mentor, Dr. Lee Niswander at the University of Colorado Denver School of Medicine, and I decided to implement an in vitro model using human stem cells and test our findings from mouse in human neural crest development.
My thesis project in the Niswander Lab aims to investigate the epigenetic regulation of neural crest cell development, a migratory cell population that can differentiate into disparate cell types. My interests in both neural crest development and the epigenetic mechanisms that regulate transcription, is what sparked the idea of a collaborative visit with Dr. Ruchi Bajpai and her lab at the University of Southern California, in Los Angeles, California. Dr. Bajpai’s research interests are very similar to my own, and we identified her as a great potential collaborator as she has pioneered the directed differentiation of stem cells into neural crest, the use of fluorescently tagged enhancers in stem cells, and also defined the epigenetic signatures of neural crest enhancers. Therefore, we sought to establish a collaboration with her lab so that I could learn the directed differentiation of iPSCs into neural crest fates and subsequently test our own hypotheses of the epigenetic regulation of human neural crest development.
With the gracious assistance from the Company of Biologists, the Traveling Fellowship that I was awarded enabled me to travel from Denver, Colorado to Dr. Bajpai’s lab in the City of Angels, also known as Los Angeles. This was a tremendously rewarding experience for me. It was wonderful being able visit a new university and with the help of Dr. Bajpai and her lab, I was not only able to learn how to successfully culture and differentiate iPSCs into neural crest, but I was also able to use this method to begin testing our own hypotheses. One important step was learning how to perform chromatin immunoprecipitation (ChIP) with our differentiated neural crest cells. I also learned the technique of lentiviral infection of stem cells, which we used in combination with fluorescently tagged enhancer sequences. Back in Denver I will continue with these methods of lentiviral based modification and the directed differentiation of iPSCs into neural crest during my thesis work investigating the epigenetic regulation of neural crest development. In the future, we hope to compare datasets that we will obtain from our own ChIP-seq experiments with those generated by Dr. Bajpai with other epigenetic regulators.
Overall the experience was fantastic. It highlights the importance of collaborative science and the impacts of a strong scientific community. I have not only improved myself as a scientist, but I have also expanded my scientific network, which will benefit me for the rest of my career. I would like to thank the Company of Biologists, and also Dr. Bajpai and her lab, for the opportunity and support.
This post originally appeared on Annotations, the DMDD blog.
New image and phenotype data for embryos and placentas from embryonic lethal knockout mouse lines has been made available on the DMDD website today. The knockout data includes the ciliary gene Rpgrip1l as well as Atg16l1, a gene encoding a protein that forms part of a larger complex needed for autophagy. In total we have added HREM image data for 10 new lines, embryo phenotypes for 11 lines and placenta image and phenotype data for 6 lines.
The new data was released at the same time as enhancements to our website, which were described on the DMDD blog. Keep reading to see some highlights from the phenotype data.
DETAILED EMBRYO PHENOTYPES REVEALED
The comprehensive and detailed nature of DMDD embryo phenotyping means that we are able to identify a wide range of abnormalities. In the data released today, a total of 423 phenotypes were scored across 78 embryos. These included gross morphological defects such as exencephaly and edema, but also abormalities on a much smaller scale such as an unusually small dorsal root ganglion, absent hypoglossal nerve and narrowing of the semicircular ear canal.
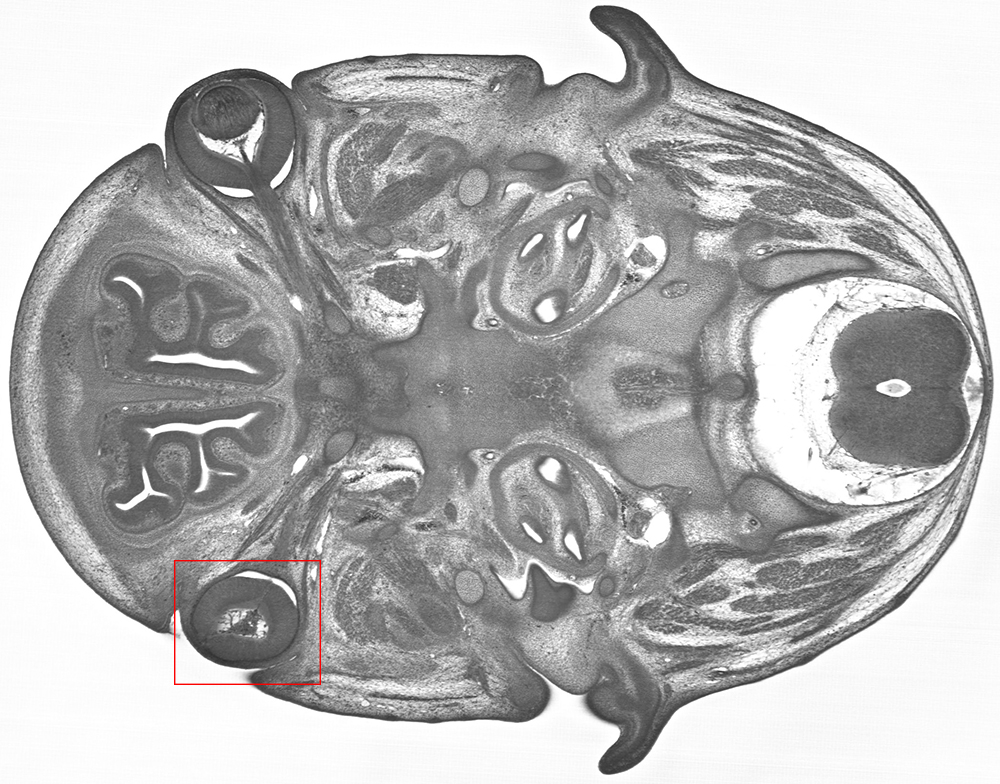
In the image below, a Trim45 embryo at E14.5, was found to have abnormal optic cup morphology and aphakia (a missing lens).
HREM imaging of a Trim45 knockout embryo reveals abnormal optic cup morphology and aphakia on the left side.
3D modelling of the exterior of an Rpgrip1l knockout embryo at E14.5 revealed a cleft upper lip, as well as polydactyly.
A 3D HREM model of an Rpgrip1l embryo shows a cleft upper lip.
All phenotypes are searchable on the DMDD website, highlighted on relevant images, and the full-resolution image data is available to explore online.
SYSTEMATIC PLACENTAL ANALYSIS
DMDD also carries out systematic phenotyping of the placentas from knockout lines. The image below shows a Cfap53 knockout placenta at E14.5, which was found to have an aberrant fibrotic lesion. The density of fetal blood vessels was also considerably reduced, the overall effect being to reduce the nutrient flow from mother to embryo.
Placental histology for the line Cfap53 shows a fibrotic lesion (large arrow) and several regions of reduced blood vessel density (small arrows).
GENE EXPRESSION PROFILES
Work is underway to measure the gene expression profiles for embryos from embryonic lethal knockout lines, a study that complements the morphological phenotype data we are gathering. One of our ultimate goals is to allow data users to explore correlations between gene, morphological phenotype and gene expression profile. The first part of this dataset was released recently – a temporal baseline gene expression profile for wild type embryos.
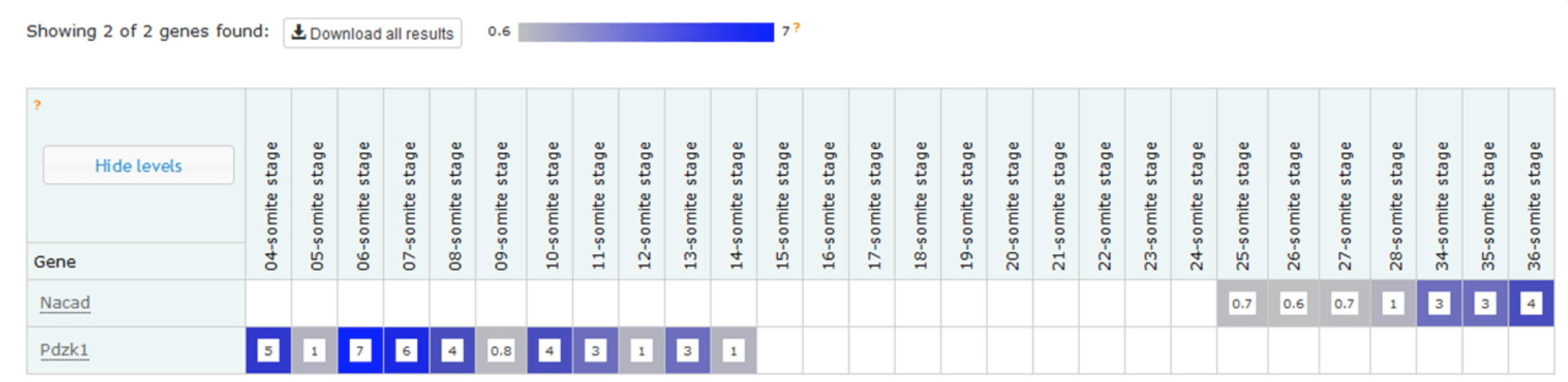
Example expression profiles of Nacad and Pdzk1 with increasing somite number. The data shows that, at this depth of sequencing, Nacad is switched on during somitogenesis and Pdzk1 is switched off.
The expression data is now accessible via a dedicated wild type gene expression profiling page on the DMDD website, which also gives background information about the analysis. Mutant expression data will follow in the new year.
LINKS BETWEEN DMDD GENES AND HUMAN DISEASE
Many of the genes studied by the DMDD programme are known to have links to human disease, including several new lines that have been made available in this release.
Cfap53: the human ortholog of this gene is known to be associated with visceral heterotaxy-6, in which organs have an abnormal placement and/or orientation.
Embryonic lethal lines with no known links to human disease may also be novel candidate genes for undiagnosed genetic disorders. Visit the DMDD website to explore the phenotype data.
Sarah, Kristian and Joy in the lab, plus some toys
So Kristian, how did you come to establish your lab, and what are the main questions you are interested in?
KF I originally started off as a vet. However, after a short time in practice I realized that all pets have owners and that I had to spend a significant amount of time with them rather than with the animals. I quickly figured out that science is much more fun and started my PhD in neuroscience. Soon after I moved towards biophysics, and started my lab 5 years ago in the Department of Physiology, Development and Neuroscience at the University of Cambridge, after finishing my postdoc at Cambridge’s Physics Department. Our lab works at the interface of life and physical sciences; we are mainly interested in how mechanical signals control cell function in the nervous system. Particularly, we want to understand how local tissue stiffness, cellular forces, and cellular mechanosensitivity contribute to CNS development, and how changes of cells’ mechanical environment impede, for example, neuronal regeneration after spinal cord injuries.
Do you think cell mechanics are adequately appreciated in current developmental biology?
KF Definitely much more than in many other branches of biology. Forces are an old concept in developmental biology. During development, cells move all the time, and there is no motion without forces. Pretty much every developmental biologist I know appreciates cell mechanics, which makes me happy.
“During development, cells move all the time, and there is no motion without forces. Pretty much every developmental biologist I know appreciates cell mechanics”
On to the current paper: I understand it has had a long gestation?
KF That’s correct. It started some 8 years ago, when I applied for a postdoctoral fellowship with Christine Holt. That’s when I came in contact with Xenopus for the first time. Since then, we have been working on the story, had to develop a number of new experimental techniques and approaches, and to learn a lot about the mechanobiology of developing retinal ganglion cells. It went through 2 generations of highly talented postdocs and graduate students in my lab, and now I’m very happy that we finally managed to publish the paper.
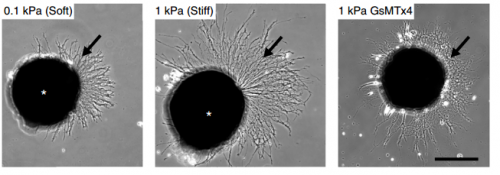
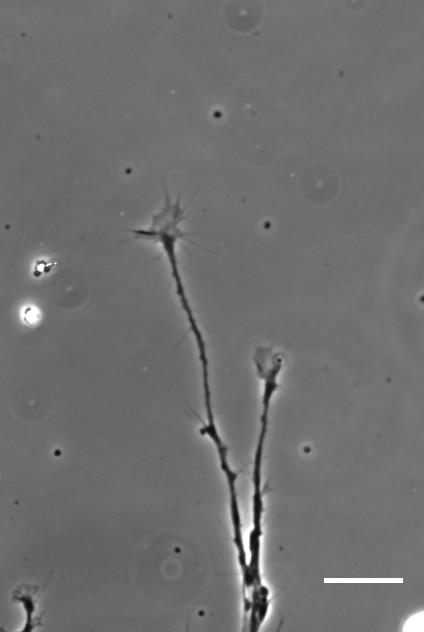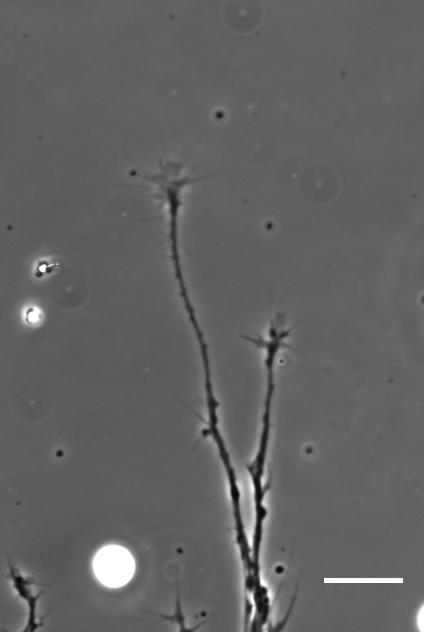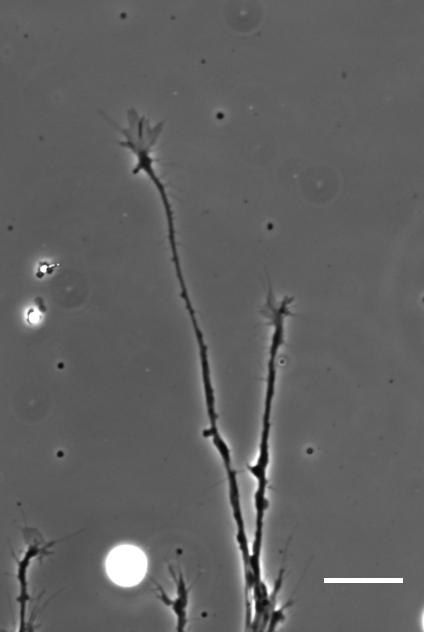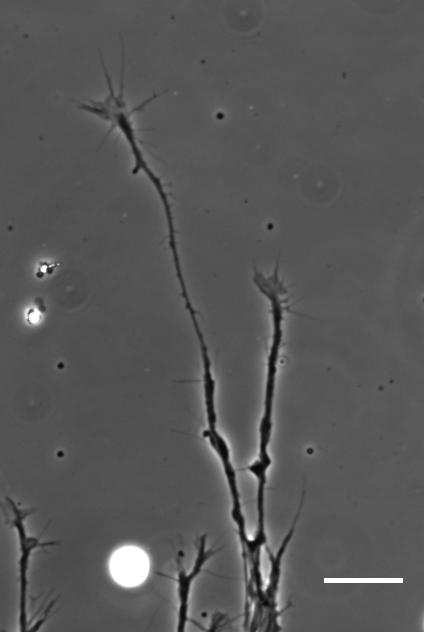
Cultures of Xenopus eye primordia on soft or stiff substrates, and with spider venom peptide added. From Figure 1, Koser, et al. 2016. Nature Neuroscience.
And Joy and Sarah: how did you come to be involved with this story? Were you working fairly separately, or was there a lot of collaboration?
AJT & SKF This story grew up over a long time! Joy started working on it before Sarah joined the lab, focusing on brain mechanics during development in vivo. Sarah started about a year later, working on the in vitro side. The experimental work was fairly independent, but there was a lot of scientific discussion behind the scenes among all the authors and other members of the lab. We have since started working on a collaborative project that uses Sarah’s expertise in in vitro cultures and Joy’s experience in atomic force microscopy.
Why choose the Xenopus retinal ganglion cell (RGC) as a model?
AJT & SKF The RGC model is great because it provides both a simple in vitro system and an elegant in vivo system. The structure of the optic tract is stereotypic and easily visualized, making it a brilliant system for seeing how different perturbations affect axon growth and guidance. To study in vivo brain mechanics, we needed an organism where the brain was both accessible in the embryo and developed at a rate that was practical for our measurement timeline. That’s quite a challenging set of requirements and Xenopus fit the bill nicely. Furthermore, and fortunately for us, the rate of Xenopus development can be sped up or slowed down by changing the temperature, which allows a lot of flexibility when planning experiments.
“The Xenopus retinal ganglion cell model model is great because it provides both a simple in vitro system and an elegant in vivo system”
KF Also, with Christine Holt we had a great collaborator who is a leading expert in Xenopus CNS development. This made the choice pretty easy.
AJT & SKF – On the downside, frogs are a less widely used model system and also have a complicated (allotetraploid) genome. They’re therefore less developed as a genetic model than other organisms like zebrafish, Drosophila, and even mice. Sometimes that can be quite frustrating—knockout frogs simply don’t exist, so other techniques, like morpholinos, drug treatments, or RNA/DNA injections must be used instead. Joy worked with Drosophila during her MPhil and undergraduate dissertation, so swapping to Xenopus embryos for the PhD felt a bit constraining at first! However, the Xenopus genome is known now, and CRISPR/Cas9 is showing great promise.
KF: In the end, the pros outweighed the cons by far.
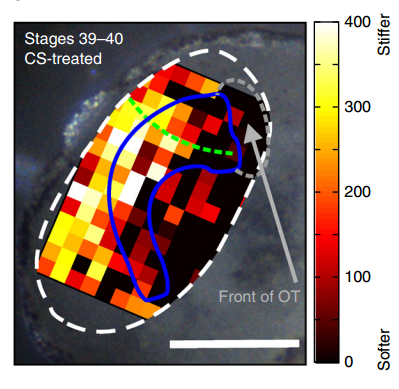
Xenopus brain overlaid with AFM-based stiffness map, from Figure 2, Koser, et al. 2016. Nature Neuroscience.
How did you go about testing the mechanics of the brain, and the responses of neurons to their mechanical environment?
AJT To measure in vivo brain mechanics, I borrowed a technique from materials science called atomic force microscopy (AFM). This is an extension of the sense of touch, just as the optical microscope is an extension of our sight. AFM uses a flexible probe (a leaf spring) to apply a set force to the brain surface; the amount by which the probe deflects (bends) is proportional to the stiffness of the tissue. It’s a bit like tapping a piece of concrete or prodding a piece of jelly. The method we used was developed in our lab over years to enable the measurement of local brain stiffness in the living embryo at cellular resolution. The AFM probe can also be used to apply a mechanical stimulus to the brain (e.g., to push on it for hours and hours) and track neuronal responses, which I did for the compression stiffening experiments.
SKF For the in vitro studies, we made compliant gels out of a polymer called polyacrylamide. Varying the ratio of two of the gel’s components allowed us precisely control the stiffness of these substrates, and we selected compositions that gave us stiffnesses near the upper and lower bounds of in vivo Xenopus brain stiffness. As brain tissue is extremely soft (comparable to cream cheese), finding the right composition was a challenge. Once we grew neuronal cultures on these substrates we were able to analyse their morphology and growth dynamics as a function of substrate stiffness using simple phase contrast microscopy.
“For the in vitro studies, we made compliant gels out of a polymer called polyacrylamide…As brain tissue is extremely soft (comparable to cream cheese), finding the right composition was a challenge.”
To further explore neuronal responses to their mechanical environment, we developed substrates containing a linear gradient in stiffness. These gradient substrates allowed us to mimic the stiffness gradients that the AFM measurements revealed in the developing brain in vivo; we could then isolate this aspect of the neuron’s environment to get a clearer idea of their mechanical response in the absence of other stimuli like chemical cues.
KF To test the importance of neuronal mechanosensitivity in vivo, we perturbed both brain stiffness and neuronal mechanotransduction in the developing embryo. While mechanically perturbing the local stiffness gradient found in the mid-diencephalon redirected axon growth, both chemically softening brain tissue and preventing neurons from detecting ‘stiff’ (either by pharmacologically blocking mechanosensitive ion channels including Piezo1 or by downregulating the expression of Piezo1) led to unusual neuronal growth patterns, and the axons never arrived at their target, the optic tectum.
Could you sum up the key results of your paper in a paragraph?
KF, AJT & SKF The key result of our paper is that neuronal growth is not only controlled by chemical signals, as it is often assumed, but also by mechanical signals. We have shown that neurons grow faster and straighter on stiffer substrates, while they slow down and grow in a more random fashion on softer ones. Piezo1, a mechanosensitive ion channel, is crucially involved in transducing the mechanical signal (here: tissue stiffness) into a response of the neuron. Developing brain tissue in vivo is mechanically highly heterogeneous and exhibits very reproducible stiffness gradients, which are likely established by changes in local cell densities in the tissue. These stiffness gradients guide neurons towards the softer side, making them turn towards their target – of course in combination with chemical guidance cues. Interfering with either tissue stiffness or neuronal mechanosensitivity was sufficient to mess up neuronal growth and pathfinding.
Time-lapse movie of cultured Xenopus retinal ganglion cell axons. Supplementary Video 2 from Koser, et al. 2016. Nature Neuroscience
So how does a cell integrate mechanical information with the classical chemical cues? Do mechanical signals carry more weight?
SKF That’s an area we’re increasingly interested in now, and in fact is the topic of my Ph.D! Growing neurons encounter a massively complex environment. They are barraged with myriad chemical and mechanical cues and must integrate all this information to arrive at a coherent response. The growth cone – a sensory-motor apparatus at the tip of the growing neuron – is amazingly complex and how it performs this ‘analysis’ is not yet clear.
We think that chemical and mechanical signals likely work together in development. We require multiple senses in order to understand our surrounding environment completely, and likewise neurons need both chemical and mechanical information to inform their growth and development.
Additionally, once mechanical cues get past the membrane they are likely carried through similar or overlapping signal transduction cascades as chemical cues. For example both chemical and mechanical guidance signals likely affect Rho/Rac signalling to control cytoskeletal dynamics in the growth cone, which eventually determines neuronal growth. That chemical and mechanical signals converge on common pathways and signalling cascades indicates that these two types of signals likely interact in many elegant and subtle ways.
“Growing neurons encounter a massively complex environment. They are barraged with myriad chemical and mechanical cues and must integrate all this information to arrive at a coherent response”
KF I’m not sure if mechanical signals carry more weight than chemical ones. It’s probably more likely that tissue stiffness is used to modify responses to chemical cues and make the responses more robust. Topological cues (such as scratches in the surface of a culture substrate), on the other hand, which I’d also classify as mechanical signals, have long been known to be able to override chemical signals and guide growing neurons or migrating cells along them. I think this is a difficult question and we are only beginning to understand the role of mechanical signals in controlling development.
Unlike in the durotaxis of cell collectives in vitro, the neurons prefer to grow towards softer tissue. Is this just an in vivo/in vitro distinction, or do you expect this preference to vary depending on developmental context?
AJT We see this in vitro as well as in vivo, at least with Xenopus retinal ganglion cells. When bundles of retinal ganglion cell axons grow over a culture substrate with a stiffness gradient, they still tend to turn towards the softer region. Since RGC axons grow faster on stiffer substrates than they do on soft, we think this ‘mollotaxis’ is at least partly a collective effect. When a bundle of axons is exposed to a stiffness gradient, the ones on the stiffer side will grow faster than the ones on the softer side. However, as the axons are tightly coupled to each other along their length by cell adhesion molecules, the faster growing ones will be held back by the slower ones and bend towards them, i.e. towards soft. It’s analogous to plants growing in the sun: the cells in the stem pointing away from the sun grow faster than the cells on the brighter side and thus push the whole plant towards the light.
Whether this is a general property of developing neurons is still an open question. Certainly previous in vitro work has shown that different neuronal populations vary in their response to soft versus stiff substrates, so there may be an element of cell type specificity. Different neuronal populations may have different preferences. Growing axons can also change their response to the same chemical cue over time – so why not to mechanical ones too?
“Growing axons can also change their response to the same chemical cue over time – so why not to mechanical ones too?”
When doing the research, was there a particularly exciting result or eureka moment that has stayed with you?
AJT The compression stiffening experiments really stood out to me. Compression stiffening is a phenomenon where brain tissue increases in stiffness when a force is applied to it. I took advantage of this to increase the local tissue stiffness in different areas of the embryo brain in vivo and see if the RGC axons changed their growth behaviour. It turned out that they grew away from the stiffened region regardless of where the force was applied. It was amazing to see such a dramatic neuronal response to a stimulus that was purely mechanical (while all chemical guidance cues were very likely present as normal)! This was an exciting experiment, but at the same time the principle was refreshingly straightforward: what would this do if I poked it?
SKF Once we got the in vitro GsMTx-4 experiments working – where we inhibited mechanosensitive ion channels and then examined neuron morphology- the phenotype of treated axons lined up really precisely with that of axons grown on soft substrates. There was a beautifully clear correspondence between those two conditions—much better than we could have hoped for!
And what about the flipside: any particular moments or frustration and despair?
AJT On my first try of the compression stiffening experiment – which took a whole weekend, and an additional two days and nights to prepare the brains for imaging – I saw to my horror that the control specimens looked very, very wrong. It turned out to be a problem with one of our stock solutions, and thus easily fixed, but it was still extremely frustrating to repeat such a time-consuming experiment for such a simple reason. (Not to mention losing my Sundays off!)
SKF Working with the GsMTx-4 – the tarantula-venom derived peptide that blocks mechanosensitive ion channels – was quite frustrating. Batches seemed to vary in strength quite significantly and also degrade and lose strength over time, so you never quite knew what was going to happen when you put GsMTx-4 on your neurons. I killed quite a few sets of cultures by overdosing them, before I figured out how to handle it properly.
Finally, Joy and Sarah: what next for you two after this project?
SKF I’m still at the beginning of my PhD, so I’ll spend the next few years trying to gain some insight into how cells integrate chemical and mechanical cues.
AJT I am working on a follow-up to the in vivo experiments in the paper, looking in more detail at the changes in brain mechanics at different stages in development. The overarching goal is to finish writing up my PhD, as I only have a year or so left now!
“If tissue stiffness changes during pathological processes, what are the consequences for cells in the tissue?”
And Kristian, where will this work take you?
KF That’s an excellent question. This work has raised many more questions than it actually answered. We are now in the great position to choose from many fascinating questions and to work on what excites us most. We will certainly continue to work on the importance of mechanical signals for neuronal and CNS development, but will also look at regenerative processes. If tissue stiffness changes during pathological processes, what are the consequences for cells in the tissue? Also, the question you asked earlier about the integration of mechanical and chemical signals is key to understanding what is really going on during development and pathology. I guess we have enough open questions to keep us busy for quite a while.
David E Koser, Amelia J Thompson, Sarah K Foster, Asha Dwivedy, Eva K Pillai, Graham K Sheridan, Hanno Svoboda, Matheus Viana, Luciano da F Costa, Jochen Guck, Christine E Holt & Kristian Franze. 2016. Mechanosensing is critical for axon growth in the developing brain. Nature Neuroscience 19, 1592–1598.
David McClay is the Arthur S. Pearse Professor of Biology at Trinity College of Arts and Sciences, Duke University, North Carolina. His lab works on the transcriptional control of morphogenesis in the sea urchin embryo. We caught up with David at the 2016 Society for Developmental Biology – International Society of Differentiation joint meeting in Boston, where he received the Lifetime Achievement Award.
You’ve been awarded the DB-SDB Lifetime Achievement Award. What does this mean to you?
Well, I think everyone likes to think that what they’ve done has mattered. In a way, the award is a reflection that somebody else has appreciated it and it wasn’t me operating in a vacuum! And the award isn’t just for the research output, but also for the development of students, the teaching, the whole thing. So that recognition was really gratifying.
Let’s go back to the beginning: did you always want to be a biologist?
My dad was a college professor, so the trade was always alluring. But then there was the question of what I would do. I switched majors several times in college – I think we all go through this process of trying to decide what’s important – and finally took a course in genetics that I found absolutely fascinating. The whole idea of how genes work together in building an organism has always fascinated me. So I decided to head off in that direction, although here I am working on an organism that’s not really a genetic model!
“The whole idea of how genes work together in building an organism has always fascinated me.”
Your early papers were on sponge aggregation, with the animals collected in Bermuda. How did that work come about?
At the time, there were two different courses you could take: the Woods Hole Embryology Course, or a separate but parallel course in Bermuda, so I applied there. Ray Keller was my lab partner that summer and it was delightful. This was between my first and second year of grad school and, though I’d picked a couple of things I’d like to head toward, I wasn’t really committed. At the end of the summer they would pick one person to bring back the following year and I decided I wanted to be that person.
So I got back to the library in Chapel Hill, and thought: what could I do that could only be done in Bermuda, and nowhere else? After reading some papers, I came up with this idea of looking at sponges and sponge aggregation. The idea was to go where the different species live cheek-by-jowl with one another and ask whether differences in cell adhesion kept them separate. So I went down to Bermuda, and that turned out to be the case: adhesive specificity is seen at the species level, rather than the tissue level as in vertebrates. That work became my PhD thesis. That was the last time I worked on sponges, but I ended up doing close to 20 summers in Bermuda. I then switched my summers to Woods Hole, partly because participation in the course there was just awesome, and partly because my kids were growing up and the Marine Biological Laboratory manages kids fantastically well, so it worked out very well when the family was young.
So once you’d transitioned to sea urchins, your initial papers were on questions of cell adhesion, aggregation and affinity. Why that particular focus?
I was interested in – and I still am interested in – the question of how the embryo ‘works’. Cell adhesion seemed to be a way in to studying this process. When I started, I was mostly trying to figure out the molecular basis of adhesion; at that time, we didn’t have cadherins or integrins, or any of that. So in my post-doc I was working on purification of adhesion molecules, and then Masatoshi Takeichi broke the field open with the cadherins, and Erkki Ruoslahti, Richard Hynes and Clayton Buck continued with the integrins. I was working on all those same molecules and got a few publications in the same mix. But then I started to think: I’m not just going to work on adhesion, there are a lot of people doing that – I want to look at the whole cell, the whole system.
The sea urchin work started out as a very simple summer project but, because of its simplicity and the variety of experiments I could do, I began to focus on it more and more. And even though at the time I was working with chick, mouse and other things, every grad student who entered the lab wanted to work on the urchin! So, by around 1990, my lab became a sea urchin lab sort of by default.
The sea urchin is one of the longest-running model organisms in developmental biology, and one you’ve worked on for much of your career. What explains its longevity?
I’d say three things. The first reason for its longevity is very human: you have to go to marine labs to get your organism! In the past, people just gravitated to these marine labs in the summer. The second reason is its simplicity; it’s very easy to work on. And the third is the number of technologies you can throw at it. I’d been affected by all three of these: I love going to marine labs, I love the organism’s simplicity and I love the technologies. It’s amazing what you can do nowadays, particularly with imaging.
“I love going to marine labs, I love the organism’s simplicity and I love the technologies.”
It is a small field. Each of the fields that are represented here at the SDB has a sort of carrying capacity. We thought Drosophila was virtually limitless for a while, but it’s turning out not to be. There are only so many questions and so many groups that can work on a given question. The sea urchin carrying capacity is smaller than others, but I still think it makes a valuable contribution to the field.
Throughout your career, you seem to have straddled the mechanics and the signalling sides of developmental biology. Was this a conscious choice?
I don’t really care what aspect I’m looking at, I just want to know how the system works. So, for instance, with the epithelial-to-mesenchymal transition (EMT) I don’t want to be restricted to one aspect, I want to know everything! The mechanics, gene regulation…whatever it is that tells me how that system works, I want to know.
Ten years ago the sea urchin genome was published and you were involved in that project. How did the completed genome complement your work?
The story really starts years before the genome publication, in the late 1990s. We’d been looking at these different morphogenetic activities and wanted to know how they’re controlled. That quest to understand their control naturally led me to transcription factors, and I got a call one day from Eric Davidson asking whether I was interested in collaborating in building the gene regulatory network (GRN) for sea urchin development. What we needed then was all the genes, and that meant the genome. And so Eric in particular worked very hard, starting in about 2002, to sequence and annotate the genome.
I’d travel around the world to annotation parties, and we’d pull a whole bunch of people together in a room and teach them how to annotate – how to go in to an open reading frame and know what to look for. People started to have great fun when their names were associated with what they had done. We ended up annotating 12,000 genes – not particularly well, but in a very short time.
There were teams of us: Eric was the overall project leader and I was in charge of one of the subteams for the cell biology side. It was a wonderful collaboration, I think there were 120-130 people altogether, and working together was really fun.
And the impact? Now when we do RNA-seq, we have a reference standard, which very quickly and computationally gives us a list of genes present at a given time or after a perturbation. We just couldn’t have done that without the genome; it’s been great.
How and why did GRNs come to feature so prominently in your work?
Eric was at the head of the sea urchin GRN consortium and I was in the middle. We shared an interest in building the GRNs but with different purposes: Eric wanted to learn the entire cis regulatory code that drove gene expression in the network, whereas my interest was and is in using GRNs as a tool for understanding morphogenesis. Once we had the GRN I could use it as a template to understand different events in morphogenesis. GRNs allow you to eliminate the mystique behind these really complex problems.
“My interest is in using GRNs as a tool for understanding morphogenesis…they allow you to eliminate the mystique behind these really complex problems.”
Our studies on EMT illustrate this point. Cells undergoing EMT go through de-adhesion, become motile, change shape, change polarity, invade through the basement membrane, and extend filopodia. Each of those could be separated as a distinct cell biological activity and, lo and behold, each activity is controlled by a different transcriptional subcircuit. So there are all these little subcircuits in the GRN that regulate this event in a coordinated manner.
But it’s not quite so simple. We knew, for instance, that de-adhesion is primarily controlled by Twist and Snail. But in a nine-hour period of sea urchin embryogenesis, five different cell types go through an EMT. My hypothesis was that all five would be essentially similarly controlled. Wrong! Twist and Snail participate in two out of the five, but the other three have different controls for de-adhesion. It’s a little bit disconcerting but that’s the way nature operates.
Your lifetime achievement award also celebrates mentorship. Do you have any advice for young researchers today?
I think all of us have somewhat mixed feelings: I love what I do, but at the same time I recognise how hard it is. You’re in two minds – trying to get across your love of science and how rewarding that can be, but at the same time recognising there’s a huge gamble that someone takes on when they’re going further into science.
I cite the statistic of a few years ago when there were 6000 PhD graduates in biomedical science and 600 opportunities in academia. That meant 90% of the people had to find a related occupation, many of which are great, but perhaps different from the initial vision that the student might have had.
It’s unlike other occupations in that the very occupation restricts the number of people who can enter it, so as well as the love and excitement, you want to paint a realistic picture of the opportunities.
Once a student does take that gamble, then it shifts: you want to optimise what they’re going to get out of it. You can either give them everything or you can set up an environment where they can build their own career. I try to provide both, but lean toward them building their own career. And I’ve been really proud of them: so many of my students have gone on into successful careers.
And a final question: what might people be surprised to find out about you?
Well, I’ve been driving the same car to work for 42 years: a Fiat Spider convertible, a beautiful old car.
“I don’t think I’m ever really going to grow up! I started going to Bermuda in the summers, and then Woods Hole, and recently Villefranche-sur-Mer – forty summers of marine labs. And I’m still having fun: give me another thirty years!”
And maybe they wouldn’t find this surprising, but I don’t think I’m ever really going to grow up! I started going to Bermuda in the summers, and then Woods Hole, and recently Villefranche-sur-Mer – forty summers of marine labs. And I’m still having fun: give me another thirty years!
Can you tell me your scientific biography leading up to this work? When did you first get interested in craniofacial development?
I first got interested in the skeleton when I was a technician at the biotechnology company Amgen, working on an osteoporosis therapy. After deciding to pursue my own scientific questions I enrolled in graduate school at the University of California, Los Angeles studying Notch signalling in cell culture with Geraldine Weinmaster, that’s where I fell in love with imaging cells. After my Ph.D., I wanted to move from cells in a dish to an in vivo genetic system. Chuck Kimmel’s laboratory at the University of Oregon was the ideal fit for me because studying zebrafish craniofacial development perfectly satisfied my interest in the skeleton, my love of imaging live cells, and powerful vertebrate genetics. Plus, Chuck is the very best mentor anyone could ever ask for, his lab is a really great place to be.
The team behind the paper includes labs from across the States: how did these labs come together for the work?
I love the collaborative nature of modern science. I think that, within the zebrafish community especially, people are eager to share their unpublished findings and reagents to help each other out and that’s what happened in this case. This open communal approach to science is not only more efficient, but also more fun since you get to work with friends from all over the place.
“I love the collaborative nature of modern science… within the zebrafish community, people are eager to share their unpublished findings and reagents to help each other out and that’s what happened in this case.”
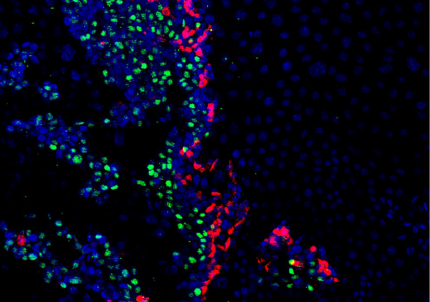
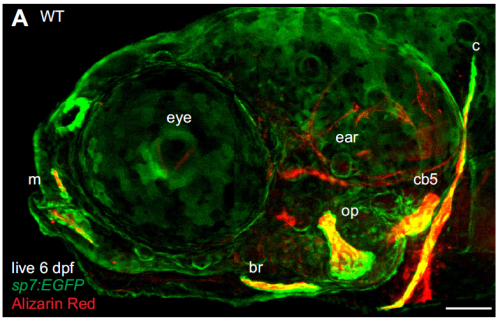
A live 6 day post-fertilisation zebrafish with osteoblasts in green and bones in red, from Figure 1 in the paper
Many researchers, confronted with a highly variable phenotype, would run for the hills or move on to another mutation. What made you guys stick it out?
While this mutant phenotype is notably variable, most mutant phenotypes are more variable than that of the wild type and no one knows why. It is tempting to run for the hills, or just report a ‘representative phenotype’, but I think we are glossing over a lot of interesting biology when we do that. Because zebrafish breed frequently and give large numbers of offspring, and the allele we focused on was behaving just plain weird, we felt like we had a good system to address some important questions about variability. Also, it was very exciting to feel like we were making progress on a real mystery that has intrigued biologists for a long time, I think that kept it fun.
“It is tempting to run for the hills, or just report a ‘representative phenotype’, but I think we are glossing over a lot of interesting biology when we do that.”
Can you sum up the key results of the paper in a paragraph?
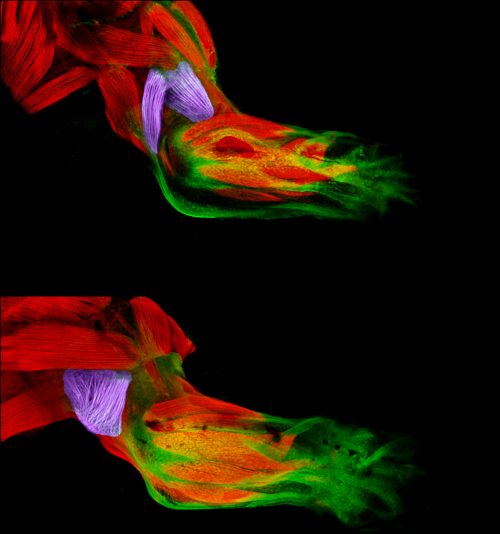
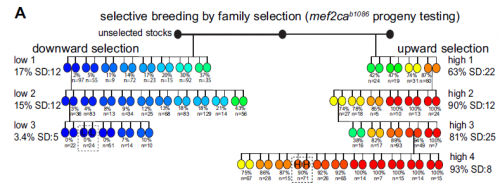
We started with an extremely variable gain of bone phenotype that some really smart people in Chuck’s laboratory had made good progress on in the past. However, this mutant had always been problematic because the extra bone phenotype would occasionally disappear. For example, when we moved the mutation onto a transgenic background to label bone cells, the mutant animals would no longer develop extra bone! In this work, we discovered that the ectopic bone was developing because of a transfating event, where bone forms instead of a previously undescribed ligament. We then showed that we could shift the penetrance of transfating up or down through selective breeding, suggesting variability is heritable. When we compared mutants from our selected high- and low-penetrance strains, we found that the high-penetrance strain had more mutant transcripts than the low-penetrance strain. These data suggested that a high level of the mutant transcript was making the phenotype more severe, which we experimentally confirmed by manipulating the transcript. These findings were a real surprise because it suggested that the allele was acting like an antimorph, or a dominant negative, yet it’s completely recessive to the wild-type allele. So it’s an unusual type of allele, a recessive antimorph. We then found that there was a transposon right at the start of the mutant gene, and this transposon was epigenetically different in the two strains. These data suggested that heritable variability in epigenetic silencing might underlie heritable phenotypic variation. Perhaps not just in this context, but in many others as well.
Selective breeding with upward and downward selection, from Figure 5 in the paper
And your selection experiments were based on those done in the early to mid twentieth century? Did you feel like you were addressing the same issues these researchers faced decades ago?
I do, and those studies certainly shaped how we interpreted our results. C.H. Waddington’s work has enjoyed a tremendous resurgence in interest lately, and his genetic assimilation studies really influenced our thinking. Waddington unveiled cryptic variation in his lines that he could selectively breed by heat shocking his flies. We revealed cryptic variation that we could selectively breed by mutagenizing a transcription factor in our fish. Also, the important experiments from Sewall Wright in which he hybridized his inbred guinea pig strains that had an extra toe or not, just like we hybridized our inbred zebrafish strains that had extra head bones or not. Wright interpreted his results as evidence for threshold characters, where if you have enough of some unknown ‘stuff’ you develop a particular phenotype. This led us to look for levels of ‘stuff’ which turned out to be mutant transcripts. It’s so fun to read the old literature and realize that it applies to what we are seeing in the lab; I love the classics!
“It’s so fun to read the old literature and realize that it applies to what we are seeing in the lab; I love the classics!”
Does your work suggest anything about the evolution of new traits?
I think it does, the idea that all this beautiful cryptic variation is just hiding under the surface, unseen in the wild type but ready to be revealed and selected upon is very attractive to me. When conditions are stable, it’s beneficial to have reproducible development. But if conditions change, it’s beneficial to have abundant variation for natural selection to act upon. It’s like having the best of both worlds, consistent development when things are good, but also the ability to provide raw material for evolution when times get tough.
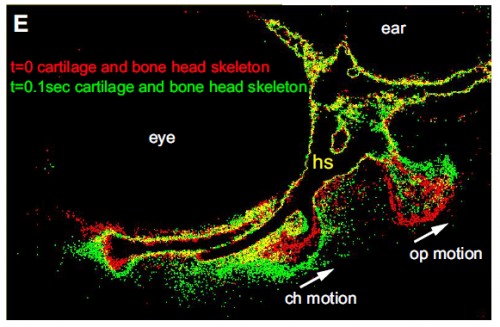
Skeletal movement revealed by overlaying images collected 0.1s apart, from Figure 1 in the paper
When doing the research, was there a particularly exciting result or eureka moment that has stayed with you?
Absolutely, when we were able to visualize the new ligament for the first time. We suspected it was there, but had no evidence either from us or anywhere in the literature of its existence. Convincing ourselves that it was really there was extremely exciting.
And what about the flipside, any moments of frustration or despair?
We struggled for some time with the concept of a recessive antimorph. How can an allele behave as a ‘dominant negative’ when it’s recessive to the wild type? When we found evidence in the literature from Arabidopsis that this sort of unusual allele exists in other systems it was a big relief.
You’ve just recently started as an Assistant Professor in Denver. How has the transition from Eugene been? What are your plans for the next few years?
I’m living the dream! It took me many years to get to this point and setting up my own laboratory has been everything I’d hoped it would be. There are fantastic colleagues here at the University of Colorado Denver, including some world-class zebrafish laboratories, that have welcomed me and are helping me to get started. Importantly, I have an amazingly supportive wife that enables my science habit. I’m surrounded by wonderful people and that has eased the transition.
“I’m living the dream! I’m surrounded by wonderful people and that has eased the transition.”
In the next few years I plan to follow up on the link between epigenetic variability and mutant phenotype variability. I’ve got some nice preliminary data that strengthens this, plus a whole bunch of results I don’t understand just yet. I’ve got plenty to keep me busy.
The author with a Colorado rainbow trout
What do you like to do when you’re not playing with fish?
When I’m not playing with fish in the laboratory, I’m playing with fish outside the laboratory; I’m an aquarium and pond hobbyist and an avid angler. There are some terrific fly fishing opportunities nearby, and I’m enjoying learning my new home waters.
Here are the highlights from the current issue of Development:
Syndecan 4 lets lymphatic endothelial cells go with the flow
Fluid flow is known to play a role in the development and remodelling of both blood and lymphatic vessels. But how is fluid flow sensed and transduced into a response? Here, Michael Simons and colleagues identify a role for syndecan 4 (SDC4) in regulating flow-induced remodelling of the lymphatic vasculature in mice (p. 4441). They first show that Sdc4−/− mice exhibit lymphatic vessel remodelling defects during the late stages of embryonic development. Notably, the alignment of valve-forming lymphatic endothelial cells (LECs), and hence valve formation, is perturbed in these mutants. The authors note that these defects are similar to those seen in mice mutant for Pecam1, which encodes a known flow-sensing molecule, but that Sdc4−/−; Pecam1−/− double knockouts exhibit a more severe phenotype, suggesting that SDC4 and PECAM1 act via distinct pathways. Following on from this, the researchers demonstrate that SDC4 acts by regulating the planar cell polarity protein VANGL2; SDC4knockdown LECs express increased levels of VANGL2 in response to flow and fail to align under flow, whereas the reduction of VANGL2 levels in these cells restores flow-induced alignment. Together, these findings uncover new regulators of flow-mediated remodelling in the lymphatic vasculature.
Optimized inducible gene knockdown and knockout in hPSCs
Human pluripotent stem cells (hPSCs) are emerging as an attractive model for studying human development and disease. However, functional studies of these cells are limited due to a lack of efficient methods for manipulating their gene expression. Here, Alessandro Bertero and co-workers devise platforms that allow for the inducible knockdown or knockout of specific genes in hPSCs and their derivatives (p. 4405). They first validate the ROSA26 and AAVS1 loci as genomic safe harbours that can be engineered in hPSCs to support stable transgene expression in a large panel of mature cells obtained from hPSCs. The authors then develop single-step optimized inducible knockdown (sOPTiKD) – an inducible shRNA-mediated approach for gene knockdown. This method allows for strong inducible expression of shRNAs, resulting in efficient gene knockdown even following hPSC differentiation. It also uses an optimized tetracycline-responsive repressor protein that eliminates leaky shRNA expression. Importantly, the authors show that this method can be used to knock down individual and multiple genes to study developmental mechanisms. They also develop a conditional knockout system based on CRISPR/Cas9 technology, named single-step optimized inducible knockout (sOPTiKO), and show that gene knockout using this system is possible in hPSCs and mature cell types. Given their robustness, high efficiency and scalability, these platforms promise to be valuable tools for the field.
Transposing from ligament to bone
Phenotypic variation among mutant animals is common, with some mutants displaying dramatic phenotypes while their genetically similar siblings seem less affected. Why is this? Here, on p. 4430, Charles Kimmel and colleagues reveal that phenotypic variation, in the case of zebrafish mef2cab1086mutants, can be caused by a fate-switching event during development. Zebrafish mef2cab1086 mutants express a truncated form of Mef2c – a protein involved in skeletal development – and are known to develop variable ectopic bones in their heads. The researchers now reveal that these bones arise due to a fate-switching event during development, such that cells that are normally destined to be ligament variably turn into bone. Selective breeding demonstrates that the penetrance of the bone phenotype is heritable. The authors further show that the mef2cab1086 transcript is differentially expressed in low and high penetrance strains. Finally, they report that a transposon that resides upstream of the mef2calocus exhibits differential levels of DNA methylation; in high penetrance strains, which express high levels of the mef2cab1086 transcript, DNA methylation of the transposon is significantly reduced. These findings lead the authors to propose that variable epigenetic silencing of transposons underlies the variable mef2cab1086 phenotypes and could explain other cases of phenotypic variability.
Bringing in fresh blood: SOX7, RUNX1, AP-1 and TEAD4
Despite being the focus of intense research in recent years, the precise mechanisms that regulate the development of haematopoietic stem and progenitor cells (HSPCs), which give rise to all differentiated blood cells, remain unclear. In particular, it is not clear how various transcription factors function together to drive the emergence of HSPCs from haemogenic endothelium (HE) during development. In this issue, two papers attempt to tackle this problem.
In the first paper (p. 4341), Valerie Kouskoff and co-workers examine how SOX7 and RUNX1 regulate haemogenic fate in the yolk sac of mouse embryos. These two factors are thought to play opposing roles: RUNX1 acts as a master regulator of endothelial-to-haemogenic transition (EHT) while SOX7 downregulation is needed for this event. Now, the authors report that, when overexpressed in ESC-derived HE, SOX7 inhibits the expression of RUNX1 target genes but has no effect on the expression of RUNX1 itself. They further reveal that SOX7 and RUNX1 are co-expressed in the yolk sac and dorsal aorta HE of mouse embryos and, importantly, can physically interact with each other via their respective HMG and RUNT domains. This interaction, the authors report, inhibits the transcriptional activity of RUNX1; the binding of SOX7 to RUNX1 prevents RUNX1 from interacting with its co-factor CBFβ and with its target DNA sites. Together, these findings highlight how direct protein-protein interactions between endothelial and haematopoietic transcription factors can regulate cell differentiation programmes during development.
In a second paper (p. 4324), Nadine Obier, Constanze Bonifer and colleagues investigate how AP-1 transcription factors regulate cell fate during the differentiation of mouse embryonic stem cells (ESCs) into haematopoietic cells. They demonstrate that the global inhibition of AP-1 factors (using inducible overexpression of a dominant-negative FOS peptide) affects various stages of ESC differentiation, as cells transition from haemangioblasts (HB) into haemogenic endothelium (HE) and haematopoietic cells. In particular, inhibition at the HB stage enhances cell proliferation and affects the balance between smooth muscle and blood cells, shifting cells towards a blood cell fate. Finally, the authors reveal that AP-1 factors bind to target genes involved in vasculogenesis; these target sites colocalize with binding motifs for TEAD transcription factors, and the authors further show that AP-1 factors are required for the de novo binding of TEAD4 to these genes. In summary, these results suggest that cis-regulatory elements that bind both AP-1 and TEAD4 act as ‘hubs’ that integrate multiple signals to regulate specific gene expression programmes during haematopoiesis.
CHK2 mediates DNA damage in adult stem cells
Adult stem cells are often exposed to genotoxic stress, whether directly from the environment or from within their own stem cell niche. DNA damage accumulates in the stem cells of aged tissues and has been proposed to accelerate both cellular aging and cancer formation, yet the mechanism through which this occurs is not well understood. Now, on p. 4312, Ting Xie and colleagues investigate this issue and demonstrate that DNA damage disrupts germline stem cell (GSC) self-renewal and lineage differentiation in a checkpoint kinase 2 (CHK2)-dependent manner. The authors use an inducible system to generate widespread double-stranded breaks (DSBs) in the GSCs of the Drosophila ovary. These DSBs resolve over time but leave the tissue with significantly fewer GSCs. By contrast, the number of GSC daughter cells initially increases then remains constant, suggesting that differentiation is blocked. The authors go on to identify a role for CHK2, showing how the induction of DSBs in flies lacking CHK2 is sufficient to prevent damage-induced GSC loss. Finally, the authors provide some evidence to suggest that the loss of GSCs may be partly due to reduced BMP signalling and cell adhesion. This study offers insight into how DNA damage might affect stem cell-based tissue regeneration and provides a mechanistic target – CHK2 – for further investigation.
PLUS:
An interview with David McClay
David McClay is the Arthur S. Pearse Professor of Biology at Trinity College of Arts and Sciences, Duke University, North Carolina. His lab works on the transcriptional control of morphogenesis in the sea urchin embryo. We caught up with David at the 2016 Society for Developmental Biology – International Society of Differentiation joint meeting in Boston, where he received the Lifetime Achievement Award. Read the Spotlight article on p. 4289.
A common framework for EMT and collective cell migration
It has long been considered that epithelial cells either migrate collectively as epithelial cells, or undergo an epithelial-to-mesenchymal transition and migrate as individual mesenchymal cells. Here, Kyra Campbell and Jordi Casanova hypothesise that such migratory behaviours do not fit into alternative and mutually exclusive categories. Rather, they propose that these categories can be viewed as the most extreme cases of a general continuum of morphological variety. See the Hypothesis article on p. 4291.
Cycling through developmental decisions: how cell cycle dynamics control pluripotency, differentiation and reprogramming
A strong connection exists between the cell cycle and cell fate decisions in a wide-range of developmental contexts. Terminal differentiation is often associated with cell cycle exit, whereas cell fate switches are frequently linked to cell cycle transitions in dividing cells. In recent years, progress to address the connection between cell fate and the cell cycle has been made in pluripotent stem cells, in which the transition through mitosis and G1 phase is crucial for establishing a window of opportunity for pluripotency exit and the initiation of differentiation. Here, Abdenour Soufi andStephen Dalton summarize recent findings in this area and place them in a broader context that has implications for a wide range of developmental scenarios. See the Review on p. 4301.















 (2 votes)
(2 votes)

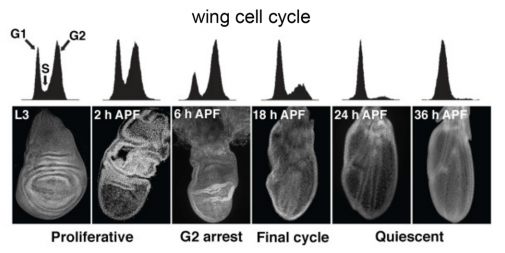




 (No Ratings Yet)
(No Ratings Yet)













 It has long been considered that epithelial cells either migrate collectively as epithelial cells, or undergo an epithelial-to-mesenchymal transition and migrate as individual mesenchymal cells. Here,
It has long been considered that epithelial cells either migrate collectively as epithelial cells, or undergo an epithelial-to-mesenchymal transition and migrate as individual mesenchymal cells. Here,  A strong connection exists between the cell cycle and cell fate decisions in a wide-range of developmental contexts. Terminal differentiation is often associated with cell cycle exit, whereas cell fate switches are frequently linked to cell cycle transitions in dividing cells. In recent years, progress to address the connection between cell fate and the cell cycle has been made in pluripotent stem cells, in which the transition through mitosis and G1 phase is crucial for establishing a window of opportunity for pluripotency exit and the initiation of differentiation. Here,
A strong connection exists between the cell cycle and cell fate decisions in a wide-range of developmental contexts. Terminal differentiation is often associated with cell cycle exit, whereas cell fate switches are frequently linked to cell cycle transitions in dividing cells. In recent years, progress to address the connection between cell fate and the cell cycle has been made in pluripotent stem cells, in which the transition through mitosis and G1 phase is crucial for establishing a window of opportunity for pluripotency exit and the initiation of differentiation. Here,