Happy New Year everyone! The past year presented us with the usual mix of quirky and interesting developmental biology, and we’ve had posts about new meetings, research, resources and more. Now it’s time to review the most popular posts of 2015!
4- A day in the life of an Axolotl lab– Annie told us what it’s like to work in a lab that uses axolotls as a model system, with plenty of photos and a couple of videos to illustrate!
5. An interview with Mike Levine– A lively chat with the winner of the 2015 SDB Edwin Grant Conklin Medal, including an audio of the famous ring of fire story!
Best rated 2015 posts
1- From the lab to the peak district– Héctor told us about his visit to the Rivolta lab at the University of Sheffield, sponsored by a Development Travelling Fellowship.
2015 was a year of records. We featured more posts than ever before- an incredible 418 posts (more than one per day!) We can also boast an average of over 6,000 visitors every month. Thank you all for making the Node so successful!
In 2015 we also celebrated the Node’s fifth anniversary. Our birthday gift was a new, more modern, look which made the site less busy and more functional. Your feedback was really important in deciding how to revamp the Node, so thank you all for completing our survey. We hope that you had a chance to come to our stall at the SDB meeting for some birthday cake!
As you can see from the list above, our A day in the life series is still going strong (and we have a little surprise for you to collect at conferences this year!), and there were also new additions to the outreach series. We also kicked off a new type of post this year, which we hope will give you more opportunities to discuss interesting and topical issues. Check out our Question of the Month posts to see what topics have featured already, and you can get in touch if you have any suggestions for future discussions.
The Node is your community blog, and it could not exist without your participation. A big thank you to all of you who wrote, commented, rated and read the Node posts in 2015. We are looking forward to another exciting year of developmental biology in 2016!
Used for thousands of years but grafting remains mysterious
For millennia, people have cut and joined different plant varieties or species together by a process known as grafting. By grafting different plants to each other, the chimeric individual acquired desirable properties from both plants, or alternatively, elite varieties could be more easily propagated. It is unknown how people first discovered that different plants could be grafted together, but nature may have provided inspiration since certain plants naturally graft with themselves (such as English Ivy) and occasionally, to other species. The first uses for grafting were likely to propagate desirable varieties of fruits such as oranges, apples, plums, cherries, grapes, figs and olives [1] since the best varieties are often not true breeding (the seeds will not produce fruit identical to the parents). Later, improving plant size or improving disease resistance may have encouraged further widespread use of this technique. A great example of this phenomenon is the collapse of the European wine industry in the 1880s due to a flying insect, phylloxera, which had arrived from North America. Similar to an aphid, it fed on the leaves and roots of European grapes that had no immunity, unlike North American grapes. The solution was to graft a North American rootstock to a European scion (shoot), thus conveying resistance to the roots where the majority of the damage occurs. Grape grafting spread throughout Europe, the Americas, Oceania and the industry was saved. Today, the vast majority of vineyards, orchards and many ornamental plants and vegetables are grafted for disease resistance, ease of propagation or to change plant size.
People have been interested in how plants graft for at least several hundred years. Texts published in the 1500’s and 1600’s described how to graft and which species could be grafted. Interestingly, much of the compatibility information was incorrect [1], and it took several centuries before a more rigourous scientific approach was taken. John Wright performed some of the first studies in the 1890s when he described the events that occurred during grafting between tomatoes, potatoes and geraniums [2]. Later, using scanning electronic microscopes, breaking weight assays and sectioning and staining for cell division and vascular connectivity shed light on the grafting process [3, 4]. These studies described a process whereby tissues initially adhere, followed by undifferentiated tissue formation (callus), and finally by vascular differentiation and reconnection [4, 5]. Cell wall components including pectins were deposited [4], and cytoplasmic channels formed between the two grafted halves [6, 7]. Despite these comprehensive descriptions, we had little idea of the mechanism or developmental process that allows plants to graft. Arabidopsis as a way forward
The majority of previous grafting studies used commercially important species such as grapes and tomatoes but their limited genetic resources made research with these species slow and labour intensive. One solution was to use Arabidopsis, the model plant. Fortunately, an efficient and rapid protocol for grafting Arabidopsis had been developed [8] and used extensively to study the movement of proteins, RNAs and metabolites [9]. We embarked in early 2012 to study how the Arabidopsis hypocotyl (the tissue between the shoot and root) reconnects, the results of which have been published recently [10]. Initially, we focused on the vascular reconnection process since previous work indicated that this was a critical step for successful graft formation [5]. The technique involved grafting seven-day-old seedlings and initially we used attachment assays and transport assays to look for the movement of fluorescent dyes and fluorescent proteins (GFP) across the graft junction. We observed a dynamic process involving attachment of cut tissues, followed by phloem connection, then root growth resumed and finally xylem connection. We performed live imaging and in situ hybridisations of the grafted tissues and observed cell division and cell differentiation occurring first above the junction (scion side) and one to two days later, immediately below the junction (rootstock side). Promoters responsive to the plant hormones auxin and cytokinin were also strongly unregulated in the region of the graft. The functional assays using GFP transport were robust and reproducible which facilitated a reverse genetics approach. Since plant hormones responses were increase and these hormones are known to be important for vascular formation, we assayed 45 genotypes associated with the auxin, cytokinin and ethylene pathways and found four genotypes, all in the auxin response pathway, were important for phloem connection. We further analysed two of these mutants (alf4 and axr1) and, surprisingly, found they were only required below the graft junction and only in the region close to the cut site (Figure 1).
Figure 1. Graft formation in Arabidopsis thaliana. Shoots expressing p35S::GFP-ER [11] were grafted to roots expressing pUBQ10::PM-tdTomato [12] and imaged with a confocal microscope after seven days (left panel). Vascular tissues, seen in the middle of this cross section (right panel), connect in part through the actions of TIR1, AFB2, AFB3, ALF4 and AXR1 [10]. The right panel is a magnified cross section through the hypocotyl of a similar plant as that seen in the left panel.
These results from our paper [10] described the dynamics of graft formation, but also established tools that we hope will be useful for the community to study graft formation. Our results also demonstrated how incredibly robust grafting is in young Arabidopsis hypocotyls: the majority of mutants, some of which are stunted and sick plants, grafted as robustly as wild type plants. The four genotypes out of 45 that affected phloem connection only delayed it by two-fold; none blocked the process. Lastly, it demonstrated an important role for auxin response in re-patterning vasculature and pointed to different mechanisms above and below the graft junction driving vascular connection.
The future for grafting
Grafting is a widely used technique in horticulture that is important for industries worth billions of dollars per year, including the wine and fruit industries. There are several ways research in grafting could benefit agriculture that should be future research priorities. Firstly, many graft combinations even within the same species are not successful and plant breeders have to develop new rootstocks that work well with a given scion. The molecular basis for this incompatibility and self/non-self recognition is unknown, but discovering the mechanism could reduce graft failure and allow wider grafts to be made between species in new combinations. For instance, wild relatives of our modern-day fruits and vegetables have often enhanced stress resistance that could be used as rootstocks to improve resistance in cultivated fruit trees, tomatoes, cucumbers, melons and potatoes. As grafting is widely conserved between dicots and gymnosperms (but interestingly not monocots), discoveries made in Arabidopsis might be conserved in other species. A future goal will be using tomato grafting as a platform to test discoveries made in Arabidopsis. In particular, understanding the mechanism and downstream targets of genes such as AXR1 and ALF4 may present targets to improve grafting efficiency.
Secondly, approximately 1% of flowering plants are parasitic [13]: they infect and attach to other species to draw out nutrients. Many parasitic plants form partial vascular connections including attaching their xylem to the host xylem [14], conceptually similar to graft formation. By better understanding how parasitic plants overcome the host/non-host recognition barriers, we could use this information to improve grafting. Similarly, blocking graft formation pathways may confer resistance to parasitic plants. Grafting is a fascinating biological process. By studying graft formation, we can better understand how plants heal wounds, repair vascular tissue, and distinguish self from non-self. Ultimately, basic research on grafting should improve and expand current grafting possibilities.
The laboratory of Sarah Goetz at Duke University School of Medicine is inviting applications for a postdoctoral researcher.
Our group studies the biology of the primary cilium. We are working to identify the signaling pathways that control the assembly of this essential organelle, as well as to define the roles of primary cilia in embryonic development and in human diseases. We use cell culture as well as mouse models to explore these questions.
Ideal candidates will be accomplished, highly motivated, and creative scientists with a recent Ph.D. in the life sciences (less than 2 years of postdoctoral experience is strongly preferred), or who anticipate completion of their degree prior to starting the position. Previous experience in cell biology, neurobiology and/or mouse genetics is desirable. Applicants should email a brief cover letter describing research accomplishments and future research goals, current CV with list of publications, and contact information for 3 professional references to:
Tunicates are the invertebrates most closely related to us, forming a monophyletic clade with the vertebrates, known as Olfactores. Tunicates, often erroneously referred to as “urochordates” (a junior synonym and thus a taxonomically invalid term), have revealed many insights into the development and evolution of chordate- or vertebrate-specific tissues and organs, such as the notochord, thyroid, liver, craniofacial muscles, heart, spinal cord, and more. Of course, one of the most important vertebrate features is a population of cells called the Neural Crest Cells (NCCs). In all chordates, the central nervous system derives from a dorsal neural plate, which rolls into a dorsal hollow neural tube. In vertebrates, NCCs are specified all along the lateral borders of the neural plate, which meet at the dorsal midline upon neural tube closure. There, NCCs undergo an epithelial-to-mesenchymal transition to form what’s been sometimes called the “Fourth Germ Layer”1. This is because NCCs are a population of stem cell-like progenitors that delaminate and migrate to give rise to a dizzying array of cell types all throughout our bodies and most of the skull: pigment cells, sensory neurons, glia, cartilage, bone, connective tissue, smooth muscle, and chromaffin cells of the adrenal medulla.
Because NCCs are so special to us, there has been much interest in understanding its evolutionary origins. There have been many theories and many disagreements, but a clear picture is starting to emerge. One of the biggest clues pointing to the origin of NCCs is one derivative cell type in particular: pigment cells containing melanin, or melanocytes. In cephalochordates, the other major chordate subphylum, cells along the lateral borders of the neural plate give rise to melanocytes associated with a series of light-sensing organs in the neural tube, known as Dorsal Ocelli2. These are not to be confused with the melanocytes of the frontal eye, which appear to be homologous to the pigment cells of the vertebrate retina instead3. In tunicates, similar dorsal melanocytes arise from the lateral borders of the neural plate and become associated with a light-sensing ocellus and a gravity-sensing otolith4. Recently, the gene regulatory networks specifying and differentiating these cells have been shown to be shared with the NCC-derived melanocytes of vertebrates, strengthening the case for homology5. Therefore, the latest models of NCC evolution propose that the neural plate borders of the pre-vertebrate ancestor already gave rise to one NCC derivative: melanocytes. Presumably, the ability to delaminate, migrate, and differentiate into several different cell types would have been added on to these ancestral melanocyte progenitors6,7.
The major questions stemming from this model are: how did the vaunted multipotency of the NCCs evolve? How did they acquire their amazing migratory capabilities? Were these traits coupled to a singular evolutionary co-option event, or have they been added on, piecemeal, over the course of chordate evolution? To begin answering these questions, one must look carefully at all those other derivatives of vertebrate NCCs Some, like cartilage and bone, are clearly a co-option of an ancestral mesoderm derivative8. Other cell types are likely vertebrate-specific, like chromaffin cells. However, the picture is less clear when looking at NCC-derived neurons, since clear homologs of these have not been found in tunicates or cephalochordates. Although the peripheral nervous systems of tunicate larvae have several sensory neuron subtypes9, none of them have been decisively linked to NCCs, either because they do not arise from the neural plate borders or because they more closely resemble non-NCC-derived sensory cells in vertebrates.
Here I describe and comment on our recent results10, which have further refined our models of NCC evolution. We were intrigued by a particular neuronal subtype that had been recently discovered and described in the tunicate Ciona intestinalis, named the Bipolar Tail Neuron (BTN)9. So called due to their bipolar, or more accurately, pseudo-unipolar morphology, bearing a single axon divided into two long neurites, or processes: an anterior, proximal process extending towards the head of the tadpole, and a posterior, distal process extending to the tip of the tail. We showed that the BTNs receive synaptic inputs from sensory cells embedded in the epidermis of the tail11. Furthermore, we also found that the BTNs synapse onto neurons of the Motor Ganglion, the spinal cord homolog of tunicates. Thus, the BTNs form a relay, part of a neural network that transmits sensory inputs from the periphery to the central nervous system.
This configuration is reminiscent of certain sensory pathways in vertebrates, in which NCC-derived Dorsal Root Ganglia (DRG) neurons relay peripheral sensory inputs from epidermal neurons known as Merkel Cells12, to neurons of the spinal cord and beyond. The shared molecular features are also intriguing, as both DRG neurons and BTNs express the transcription factor genes Neurogenin (Neurog) and Islet 13-15, and mechanosensation-modulating Acid-Sensing Ion Channels (ASICs)16. These parallels prompted us to wonder whether the BTNs, like the melanocytes of Ciona, could represent an evolutionary forebear of the NCCs. We reasoned that further support for this possibility would come from identifying the embryological origins of the BTNs (whether they originate at the borders of the neural plate), and characterizing their development (whether they engage in delamination and migration).
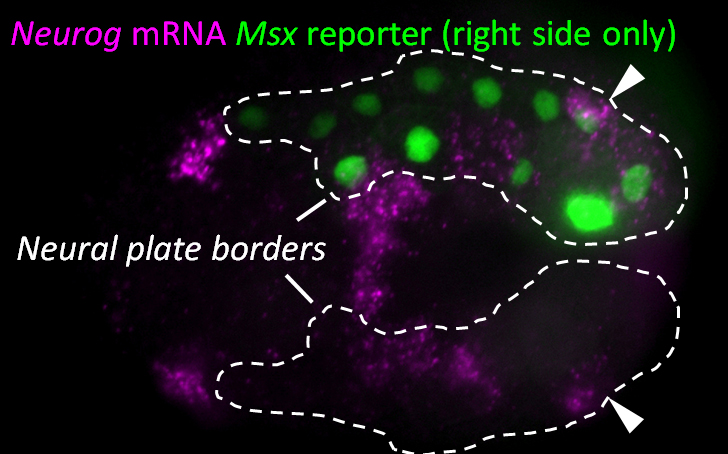
Figure 1. Early neurula stage Ciona embryo, electroporated on the right side with Msx>Beta-Galactosidase revealed by immunofluorescence (green nuclei). Msx+/Pax3/7+ Neural plate borders outlined by dashed lines. Neurogenin (Neurog) mRNA expression detected by in situ hybridization (magenta). Arrowheads indicate Neurog expression in caudal neural plate border cells (bipolar tail neuron progenitors).
Since we knew that the BTNs express Neurog, we looked at the expression of this gene by mRNA in situ hybridization, and found that it is activated in two lateral rows of cells at the very posterior end of the embryo at the early neurula stage (Figure 1). We showed that these cells are at the boundary between the epidermis and neural tissue, and express Msx and Pax3/7, conserved markers of the neural plate borders in all chordates. In vertebrates, NCCs arise from Msx+/Pax3/7+ neural plate border cells, and in fact these genes encode upstream regulators of many NCC specification genes17. Using a Neurog reporter gene to follow these cells throughout development, we found that BTN precursors delaminate shortly after neural tube closure, and migrate as a simple chain of two cells on either side of the neural tube (Figure 2). We found that they migrate outside the neural tube, along paraxial mesoderm, evocative of the migration of NCC-derived DRG neuroblasts along paraxial mesoderm on either side of the vertebrate neural tube. As the BTNs migrate anteriorly to the middle of the tail, their intrinsic polarity is reversed and begin to extend their posterior processes, suggesting a precisely timed re-orientation of cell polarity underlies their distinctive bipolar morphology.
Figure 2. Mid-tailbud stage Ciona embryo electroporated with b-line-specific Neurogenin>YFP reporter plasmid (green) and Nodal b8.18>H2B::mCherry reporter plasmid (magenta nuclei). Nodal b8.18 reporter marks descendants of the b8.18 blastomere. The anterior bipolar tail neuron (aBTN) descends from the b8.18 blastomere, while the posterior bipolar tail neuron (pBTN) derives from the tail tip of the embryo (Nodal-negative), likely descended from the b8.21 blastomere instead. On either side of the bilaterally symmetric embryo, the aBTN and pBTN migrate as a simple chain of two cells along the paraxial mesoderm outside and lateral to the neural tube.
We also investigated the molecular control of BTN specification and differentiation, and found that this is under the control of Neurog itself, which in turn is tightly regulated by FGF/ERK signaling. While we are still working to sort out these steps in detail, our current findings as they stand have some interesting implications for models of NCC evolution. Mainly, we present evidence that the BTNs and the DRG neurons might be homologous, which would imply that last common ancestor of vertebrates and tunicates might have had neural plate borders capable of giving rise to more than one NCC derivative cell type, in this case, pigment cells and sensory relay neurons. Furthermore, our observation of the delamination and migration of the BTNs along the paraxial mesoderm suggests that some aspects of NCC migratory behavior may also pre-date the emergence of vertebrates and bona fide NCCs.
Our new model, building on previous models, posits that the olfactorean ancestor already had neural plate borders capable of generating both pigment cells and sensory relay neurons, which in vertebrates are two derivatives of NCCs. In the highly reduced tunicate embryo, pigment cells and sensory neurons arise from opposite ends of the neural plate borders. However, recall that cephalochordates have serial pigmented dorsal ocelli, so perhaps the original chordate ancestor generated multiple pigment cell and sensory neurons all along the neural plate borders. In this model the real novelty of vertebrate NCCs is not the ability to give rise to multiple cell types, which in our model is an ancestral character. Instead, the key difference would have been the intercalation of a multipotent stem cell-like regulatory program within the ancestral network, downstream of neural plate border specification and upstream of multiple cell type differentiation programs18. This multipotent progenitor state, a hallmark of NCCs, allowed for these cells to undergo a prolonged period of delamination, migration, proliferation, and more dynamic modes of downstream cell fate specification and differentiation. In sum, our findings lend further support to the longstanding idea that vertebrate NCCs were not an abrupt vertebrate innovation, but rather derived from a set of ancestral features, elaborated over the course of chordate evolution19.
The Alberto Monroy Fellowship is awarded to an Italian citizen to support their attendance in the Embryology Course next summer at the Marine Biological Laboratory in Woods Hole, Massachusetts. The amount of the fellowship is 5,500.00 euros (about $6,000.00). The full text of the announcement of the award competition for 2016 can be found at http://www.giomolab.it/AAM/bando%20Monroy%202016.pdf. The deadline for applications is February 1, 2016.
Applications are invited for a postdoctoral position at the University of California San Francisco, Broad Center of Regeneration Medicine and Stem Cell Research, in the laboratory of Dr. Katja Brückner.
The laboratory investigates molecular principles of how the peripheral nervous system and its inputs regulate stem cell niches and tissue microenvironments during animal development and homeostasis. We utilize established Drosophila melanogaster models that address the regulation of hematopoiesis by sensory neurons and extrinsic stimuli. The lab also aims to expand the research program into other organ systems in Drosophila, and complementary systems of murine stem cells and self-renewing tissue macrophages. Projects are federally funded by NIH R01 and other NIH and NSF grants.
The ideal candidate is a talented, determined and creative scientist with a solid background in developmental biology and genetics. Prior work with Drosophila genetics, invertebrate or vertebrate neurobiology or hematopoiesis/macrophage biology is preferred. General basic skills in molecular biology, histology and cell-based assays are expected. Candidates must hold a recent PhD, MD or MD/PhD degree (or anticipate such a degree prior to starting the postdoc), have prior publications and be motivated and hardworking.
UCSF and the Broad Center offer a vibrant scientific environment, competitive salary and benefits. Please send a CV, statement of research interests and names and contact information of three references to: katja.brueckner@ucsf.edu
The three-pound lump under our skulls that allows us to speak, run and function in our daily lives is a mass of dozens of types of minuscule cells joined in an intricate web of communication. We pop out of the womb with our brains ready for us to interact with the world, but throughout our lives, new cells are born and others die, connections form and wither. There is a group of cells that makes this possible. They are our brain’s store of stem cells, which can become all of the different types of cells of the brain, including neurons and glial cells.
The brain’s stash of stem cells is important for both maintenance and healing after injury. Some of them become neurons, others glial cells, and some remain as the trusty store of stem cells. When cells change from the stem cell version to a specific type of cell, this is called activation (in scientist lingo). Ana Martin-Villalba and her team of researchers at the German Cancer Research Center wanted to know what makes a stem cell change from the dormant state to this activated one.
They looked at the SVZ (subventricular zone), which is a part of the brain with a hub of stem cells. Using human cells, they looked at the genes expressed in individual cells. Let that one sink in. We can study the genetics of separate cells, and taken together look at the gene expression of whole groups of cells. Based on the gene expression, they distinguished two groups, quiescent and active. The quiescent cells were broken down further into dormant and primed cells, which are ready to become active. The researchers then compared the dormant and active cells, to see how they acted differently. They found that the dormant cells were churning out proteins, the molecules that cells use to function and communicate, whereas the active cells were busy specifying more into specific cell types (a process known as differentiation).
This month’s image looks like a mosaic with each tile representing a single gene in a single stem cell. The cells in this grid were undergoing a simulated stroke, receiving decreased blood flow and oxygen. Simulating this physiological perturbation and examining the response of the stem cells showed that just as there are subsets of cells under normal conditions, subsets of cells respond differently under injury. Not only that, the response showed activation. This study solves a piece of the puzzle of how the cells are being stimulated to become active. A molecule called interferon gamma helps orchestrate the transition.
Understanding the stem cells in our brain can have important implications for helping our brains heal after brain injury or in neurodegenerative disorders.
The use of green fluorescent protein (GFP) has revolutionised the study of dynamic cellular processes in cells, tissues, and whole organisms. Laboratories throughout the world have exploited the simplicity of GFP as an everyday tool to determine protein localisation in cell lines and whole organisms. Fluorescence microscopy now dominates the imaging field although the attainable resolution of these methods has always been a limiting factor. Alternative techniques such as electron microscopy offer higher resolution but have traditionally been viewed as slow and technically difficult.
An electron microscopic method that provides rapid, simple, high resolution, and quantitative detection of GFP-tagged proteins would represent a significant advance in the fields of cell and developmental biology. Ideally this method would be compatible with the new 3D electron microscopic techniques now available, including electron tomography and serial blockface scanning electron microscopy and not require new or highly specialised equipment. We have now developed a new technique that we believe fulfils all these requirements (Ariotti et al, 2015).
We have shown that we can target a recently developed modified peroxidase, called APEX (Martell et al, 2012), to any GFP-tagged protein of interest by co-expression with a GFP-binding peptide directly attached to the APEX-tag (Ariotti et al, 2015). The APEX peroxidase produces an electron dense reaction product at the site of the GFP-fusion protein that allows its visualisation in the electron microscope. With this method whole libraries of GFP-tagged proteins could be localised at electron microscopic resolution in a few days. Moreover, the method was shown to generate remarkably high-resolution localisation data. We can localise proteins to distinct subdomains of microdomains of subcellular organelles such as individual intraluminal vesicles within endosomes and specific protein subdomains of the neck region of caveolae. This method proves to be a very powerful and versatile technique for EM analysis. It is quantitative by linescan analysis, compatible with tomography and serial blockface electron microscopy for 3D analysis of whole cell protein distribution, and offers a simplistic alternative to involved correlative light and electron microscopy techniques that are currently available in the literature.
Perhaps of most interest to developmental biologists is the application of this technique to localisation of GFP-fusion proteins in whole organisms (Ariotti et al, 2015). The effort and cost required to generate entirely new and separate gene edited lines for fluorescence and electron microscopic analyses (by conventional APEX tagging methods) are prohibitive. We bypassed this bottleneck by generating new zebrafish lines that express APEX-GFP binding peptide in all tissues. We designed two different zebrafish lines under the control of two different promoters. The first line exploits a beta-actin2 promoter for ubiquitous and continual expression of GFP-binding peptide-APEX fusion in all cells. The second zebrafish line allows for temporal control of the expression of the EM-marker, as it was engineered under the control of the hsp70I promoter that, after a brief and mild heat shock, induces rapid expression and accumulation of the APEX-GFP binding peptide in the cytoplasm of all cells within in the animal. With a single cross between our gene edited lines and any zebrafish stably expressing a GFP-tagged protein we could show rapid electron microscopic localisation of proteins including those expressed at endogenous levels in vivo. By the exploitation of the hsp70I promoter it will be possible to track subcellular protein distribution changes during development to a resolution of approximately 10 nm rapidly and easily. To further simplify the system, both lines were engineered with a red lens to denote expression of GFP-binding peptide-APEX fusion protein for easy genotyping of progeny.
We envisage that this method will provide a rapid localisation strategy in cells and whole organisms. It does not require new or expensive EM equipment, but rather is adapted from an established protocol that has been standard in the electron microscopy field for decades. While we have only demonstrated the utility of the method in zebrafish embryos the technique will not be limited to this particular animal model and we predict will be of use in other species.
It’s almost the New Year! The Node will be a bit quieter than usual for the next few days and we might take longer than usual in replying to messages.
But not to worry! If you’re craving some developmental biology, you can always have a look at our latest research posts. And if you still need to find a few last minute presents, make sure to check Cat’s science gift ideas, and let us if you’ve found anything that you’d like to add!
The New Year will bring with it new research, so why not share your 2016 research wish list with us? And while you’re at it, have some fun (and find out how far in development you are) with the Node’s DevBio Quiz!
Happy holidays and a happy New Year from the Node!
 1- The (developmental) biologists reading list– Cat asked about the books that every biologists should read, and you had plenty of suggestions to offer!
1- The (developmental) biologists reading list– Cat asked about the books that every biologists should read, and you had plenty of suggestions to offer!

 (1 votes)
(1 votes)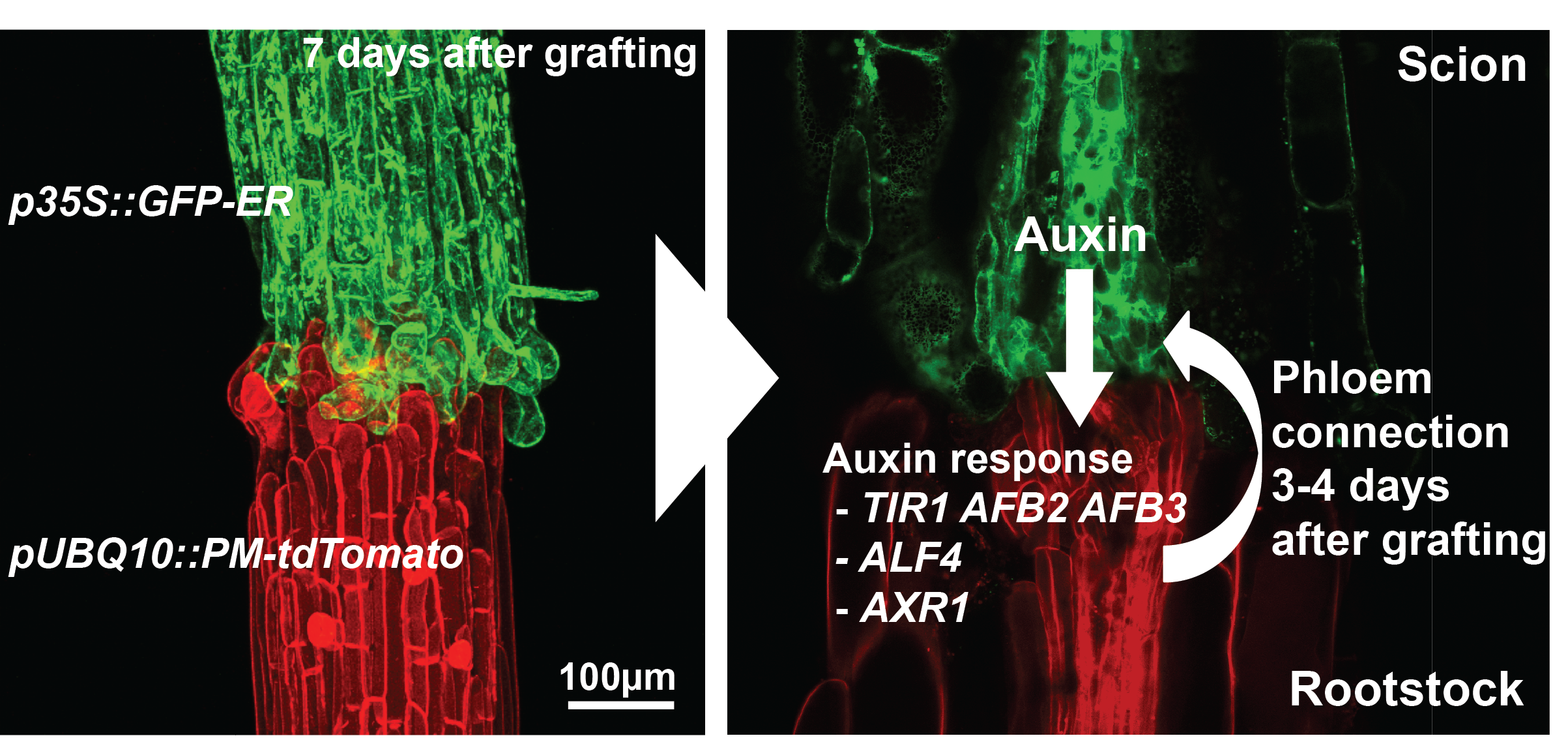
 (2 votes)
(2 votes)