It’s getting to that time of year when we think about the year to come and the things we hope it will bring: love, health, money… and maybe a little bit of help with our research.
There are many things that could make the year 2016 better for a developmental scientist: new equipment, better protocols, an easier way to maintain a model organism, or quite simply some more time to do experiments! So, in the spirit of the festive season, this month we are asking:
What is your research wish list for the New Year?
Share your thoughts by leaving a comment below! You can comment anonymously if you prefer. We are also collating answers on social media via this Storify. And if you have any ideas for future questions please drop us an email!
Here is some developmental biology related content from other journals published by The Company of Biologists.
Genome-wide lacZ profiling in the mouse
Tuck, Estebel and colleagues have produced an open access adult mouse expression resource with the expression profile of 424 assessed in up to 47 different organs, tissues and substructures using a lacZ reporter gene. Read the paper here [OPEN ACCESS].
A method for labelling leukocytes infiltrating the mouse retina
Sim and colleagues describe a method to image myeloid cells infiltrating the mouse retina in vivo using a depot injection of indocyanine green dye (ICG). Read the paper here [OPEN ACCESS].
Urine-derived stem cells predict patient response to cholesterol-lowering drug
Si-Tayeb, Idriss and colleagues have shown that cells derived from patient urine samples can be reprogrammed to generate patient-specific models of hypercholesterolemia, providing a tool to predict patient response to cholesterol-lowering drugs. Read the paper here [OPEN ACCESS], and read the press release here.
Mass and drought resistance linked in seedlings
Seedlings of large-seeded plants are considered to withstand abiotic stress more efficiently. Meng and colleagues show that the integration of drought stress response into the regulation of mass is mediated by the transcription factor ARF2. Read the paper here.
New role for eIF6 in myofibroblast differentiation
Myofibroblast differentiation is regulated by TGF-β1. In this paper, Yang and colleagues show that eukaryotic initiation factor 6 can modulate myofibroblast differentiation by altering the occupancy of the TGF-β1 promoter by H2A.Z and Sp1, affecting TGF-β1 transcription. Read the paper here.
High-salt exposure increases cardiovascular defects in early chick embryos
In chick, embryonic mortality at early stages is usually due to vascular malformations. Wang and colleagues show that high-salt exposure results in angiogenesis and heart defects, possibly due to excess generation of reactive oxygen species (ROS). Read the paper here.
The effects of rearing amphibious fish out of water
Wells, Turko and Wright rear an amphibious fish in and out of water, and find that embryos reared in aqueous environments consume more energy than their faster developing terrestrial counterparts. Read the paper here.
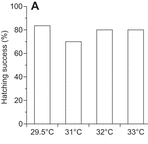
Hatching success does not decrease with higher nest temperatures in flatback turtles
Elevated nest temperatures caused by climate change could harm the viability of ectotherm eggs. Howard and colleagues show that high nest temperatures do not decrease hatching success in flatback turtles, and observe a high pivotal sex-determining temperature in these turtles. Read the paper here.
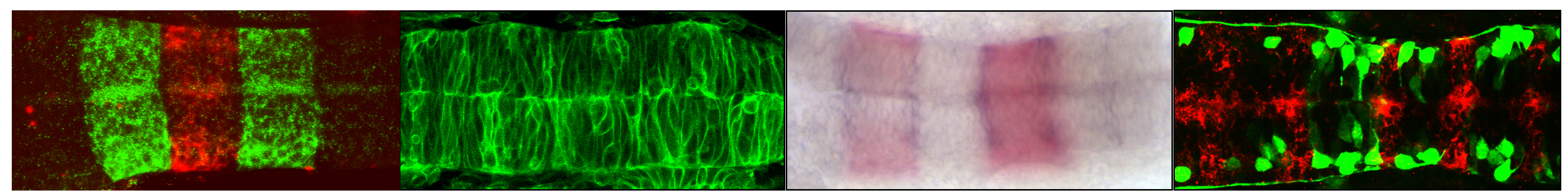
The electroporation technique is widely used in developmental biology to deliver foreign DNA into cells and study gene function. The chick embryos exhibit a remarkable easy access to perform electroporation and follow in ovo development.
Electroporation of limb somites allows the misexpression of genes in limb somite derivatives, like myogenic and endothelial cells, while electroporation of the lateral plate targets other limb cells like cartilage, bone and tendons.
We previously performed limb somite electroporation using two plasmids, one containing the gene-of-interest and one containing the reporter gene. However, this approach does not allow analysis at the cellular level, which constitutes a limitation of the technique.
To overcome this limitation and perform analysis at the cellular level, we aimed to design bicistronic vectors to misexpress, in the same cell, the gene-of-interest and the reporter gene. To do this, we took advantage of the 2A peptide that allows expression of a bicistronic mRNA. In these vectors, a single peptide is produced by the bicistronic mRNA, and auto-cleavage of the 2A peptide subsequently produces equal amounts of the two proteins. To test our vectors, we used control plasmids expressing two fluorescent proteins, Tomato and GFP, separated by the 2A peptide. We further used the Tol2 transposon system to allow genomic integration of the construct and enable analysis at late developmental stages.
One focus of our research is skeletal muscle and tendon formation. Therefore, we designed a set of stable bicistronic vectors containing different promoters to target muscle cells at different stages of differentiation. After performing limb somite electroporation using these vectors, we observed a simultaneous cellular expression of Tomato (membrane) and GFP (nuclei) at the different stages of muscle differentiation. In electroporated limbs, the ubiquitously expressed CMV/βactin promoter targeted both muscle progenitors (Pax7+ cells) and differentiated cells (myosin+ cells). The p57MRE/βactin promoter, which drives expression in differentiated myoblasts, targeted mononucleated (myosin-) cells and muscle fibres (myosin+). Finally, the MLC (myosin light chain) promoter targeted differentiated cells (myosin+). Lateral plate electroporation with the vector containing the CMV/βactin promoter allowed Tomato and GFP co-expression in cartilage, tendons and connective tissue of the limbs, but never in myogenic (Pax7+ or myosin+) cells.
We believe that this set of tools can be used to efficiently misexpress genes at different time points of myogenic cell differentiation and analyse the consequences for muscle development. Moreover, because these vectors can be integrated into the genome, the analysis at late developmental time points can be performed. Finally, the combination of limb somite and lateral plate electroporation can provide us with a tool to study the molecular and cellular interactions between the different components of the musculoskeletal system.
We are seeking to appoint a Post-doctoral Bioinformatician to undertake research on the bioinformatics of eye cancers with Prof. Shin-ichi Ohnuma and Dr. Mandeep Sagoo.
The work aims to understand the mechanism of cancer initiation and progression through bioinformatics data analysis of whole genome sequences and mRNA expression profiling in combination with single cell biology. The position is available for 32 months in the first instance.
This project builds on the previous basic cancer biology works of Prof. Ohnuma and on the patient-based clinical retinoblastoma research of Dr Sagoo.
Applicants should have a PhD and research experience in bioinformatics. Previous experience in analyses of next generation genome sequence data and mRNA expression profiling is preferable. Applicants must beself-motivated, havetheability to plan and interpret experimental studies as well as have excellent communication skills and ability to write well.
The laboratory is well equipped and is part of UCL Institute of Ophthalmology. The Institute comprises about 40 research groups spanning a research area encompassing visual development, stem cell based study, and therapy of retinal diseases. UCL has active neuroscience, developmental biology, and stem cell research communities and the group has established collaborative links within the institute and UCL, as well as with national and international colleagues.
UCL Reference: 1519787
Applicants should apply online through UCL Job search webpage.
15—17 March 2016
St. Catharine’s College, Cambridge, UK
The aim of this Biochemical Society focused meeting is to explore the mechanisms of BMP signal transduction and regulation of signalling, and discuss how genetic and epigenetic alterations result in aberrant signalling and how this leads to cancer. The BMP signalling pathway is a key therapeutic target and we will consider how to manipulate the pathway with small molecules or biologics. This conference will bring together researchers from academe and industry, and will be a great opportunity for those working on BMP signalling, cancer, stem cells, structural biology and drug discovery to share data and interact in a vibrant, but informal environment.
Organizers
Caroline Hill (The Francis Crick Institute, United Kingdom)
Alex Bullock (University of Oxford, United Kingdom)
Gareth Inman (University of Dundee, United Kingdom)
Topics
Mechanisms of BMP signalling
Deregulation of BMP signalling in cancer and therapeutic approaches
Genetic and epigenetic alterations in BMP pathway components
BMP signalling in disease
Details of the event and a list of speakers can be found here.
There will be opportunities for abstracts to be elevated to short oral talks, particularly from early-career scientists.
We are seeking for highly motivated and enthusiastic candidates interested in understanding how tissue compartmentalization and cell fate decisions take place in the Central Nervous System during embryonic development. The project will focus in understanding the genetic mechanisms involved in the control of the cellular fate using zebrafish embryos as a model. Experience on molecular biology, imaging, and developmental biology will be an advantage.
We are located in the Department of Experimental and Health Sciences of the Universitat Pompeu Fabra in Barcelona. Interested candidates are encouraged to contact Cristina Pujades for more information. To apply, send a letter of interest, CV, a short description of your research background, and contact details of 2 referees to cristina.pujades[at]upf.edu http://pujadeslab.upf.edu/
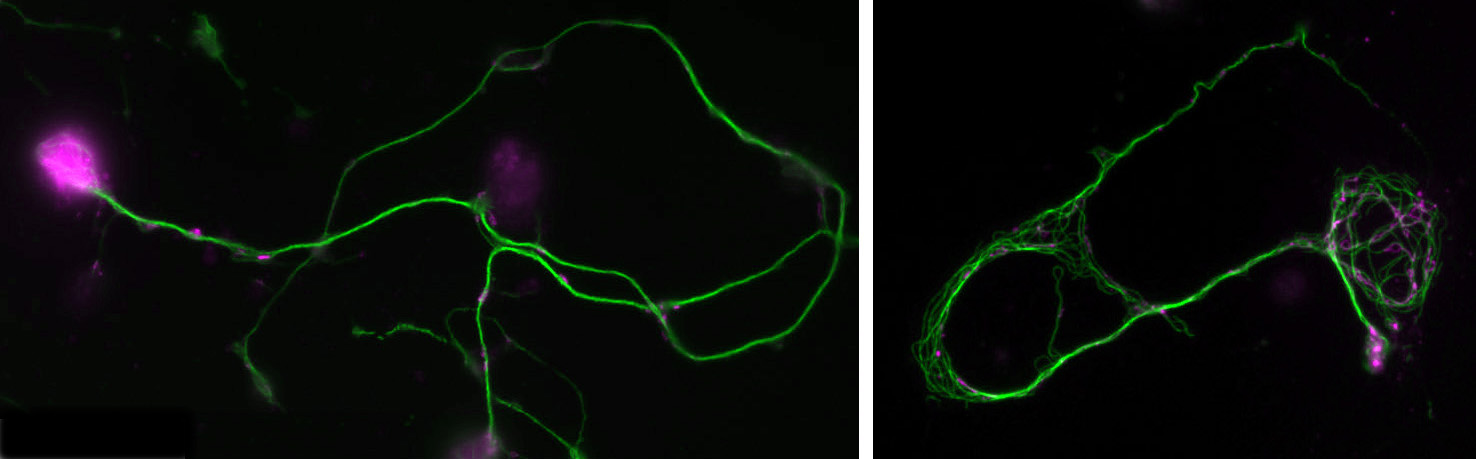
Two positions are available as part of two Research Council-funded doctoral training programmes at The University of Manchester, the first one supported by the BBSRC and the second one by the MRC. Both projects involve work on the fruit fly Drosophila as a highly efficient and relevant model organism to study fundamental mechanisms of neuronal ageing and degeneration with unique detail and depth and delivering understanding of high biomedical applicability. Further information about fruit flies as a model organism is available here. Note that Andreas Prokop, who supervises on both projects, drives active programmes of science outreach and public engagement (see here), and students will have unique opportunities to develop transferable skills in science communication, which is of increasing relevance in modern science and an important category on a researcher’s CV. Full details on how to apply for these positions can be found on the UoM BBSRC DTP website.
Normal and mutant primary Drosophila neurons in culture.
Position 1:Advanced imaging and mathematical modelling of ageing and neurodegeneration in the nervous system
Project: Do you have mathematics skills, are keen to combine them with biological research and to work on a novel, highly interdisciplinary project investigating processes of ageing and degeneration in the nervous system? To carry out this exciting research you will be trained in and apply advanced live imaging, electron microscopy (EM), tomography, genetics and mathematical/computational modelling.
You will study axons, which are the cable-like (~1mm in diameter, metres-long!) extensions of neurons wiring the nervous system. These delicate structures are maintained for many decades in humans. They are key lesion sites in spinal cord injury, trauma and many neurodegenerative diseases (e.g. spastic paraplegia), and we lose 50% of our axons during healthy ageing. The essential structural backbone of axons is formed by continuous bundles of filamentous protein-polymers called microtubules. Disorganisation of these microtubule bundles leads to axon swellings correlating with axon decay, but the underlying mechanisms preventing/causing disorganisation are not at all understood.
To gain this understanding, you will genetically induce MT disorganisation, and use imaging to capture volume and space relationships (EM) as well as the dynamics of MT disorganisation processes (live imaging). Based on your collected data and existing algorithms for MTs, you will develop computational/mathematical models describing MT disorganisation and deduce the underlying rules.
Supervision:
Outstanding supervision and training will be provided through an interdisciplinary consortium of specialists with longstanding expertise in the research areas and technologies/strategies involved in this project:
Andreas Prokop (developmental and cellular neuroscience, Drosophila genetics, live imaging, electron microscopy)
Simon Pearce (mathematical modelling of biological systems)
Karl Kadler (electron microscopy, tomography, high pressure freezing techniques)
Note, that Andreas Prokop and Matthias Heil are also members of the Wellcome Trust-funded PhD programme “Quantitative & Biophysical Biology” providing further opportunities in this area of research.
Dementia causes enormous personal hardship and costs the UK ~£23 billion every year. The second most common form is Frontotemporal lobar degeneration (FTLD). About 40% of FTLD cases have genetic causes, with >8% involving abnormal intronic GGGGCC hexanucleotide repeat expansions in the C9orf72 gene which can additionally cause motor neuron disease (OMIM #105550). These pathological expansions are actively transcribed and, via bidirectional repeat-associated non-ATG (RAN) translation, generate 5 different aggregate-forming GA, GR, PR, GP and AP dipeptide repeat proteins (DPRs).This project will gain new understanding of this type of FTLD by unravelling neurodegenerative pathomechanisms of DRPs through using interdisciplinary approaches. We will focus on the hypothesis that toxicity is caused by DRP structure, comparable to amyloid plaques in Alzheimer’s disease. The project will capitalise on the complementary expertises of the three supervisors, and a readily available, unique set of 4 GFP-tagged constructs with high, pathologically relevant repeat numbers. The detailed aims and outcomes are:
To generate purified DRPs and perform biochemical and biophysical analyses, in order to understand the reasons for their toxicity and identify useful therapeutic strategies which will benefit patients and their families.
To generate transgenic Drosophila fly stocks to obtain primary neurons expressing the four DRPs. We will use powerful fly genetics and well established cell biological approaches to identify the neuronal death pathway and of the DRP-induced pathomechanisms upstream.
There is substantial proof-of-principle for the use and translational potential of Drosophila To validate identified DRP pathomechanisms in mammalian contexts, we will carry out complementary experiments using well established DRP models in SH-SY5Y cells and inducible neuronal cell lines.
This project is highly interdisciplinary, with the analysis of the dipeptide repeat proteins spanning systems from in vitro solvents to human and fly cell lines. Students will receive a broad training in characterisation of polypeptide structure and aggregation, cell culture, fly and human genetics, and cell biology, all in the context of genetic disease and neurodegeneration.
Andreas Prokop (developmental and cellular neurobiologist; pioneered primary neurons of the fruit fly Drosophila as a powerful model to decipher the fundamental cell biology of neuropathologies)
Andrew Doig (biochemist specialised on the structural and mechanistic analysis of protein aggregation in the context of neurodegeneration)
In 2014, the British Society of Developmental Biology (BSDB) has initiated the Gurdon Summer Studentship program with the intention to provide highly motivated students with exceptional qualities and a strong interest in Developmental Biology an opportunity to engage in practical research. Each year, 10 successful applicants spend 8 weeks in the research laboratories of their choices, and the feedback we receive is outstanding. Please, read the student reports, kindly sent to us by Oliver Davis from Brighton and Sussex Medical School who was hosted in summer 2015 by Jean-Paul Vincent at the Crick Institute.
Dying for a pattern
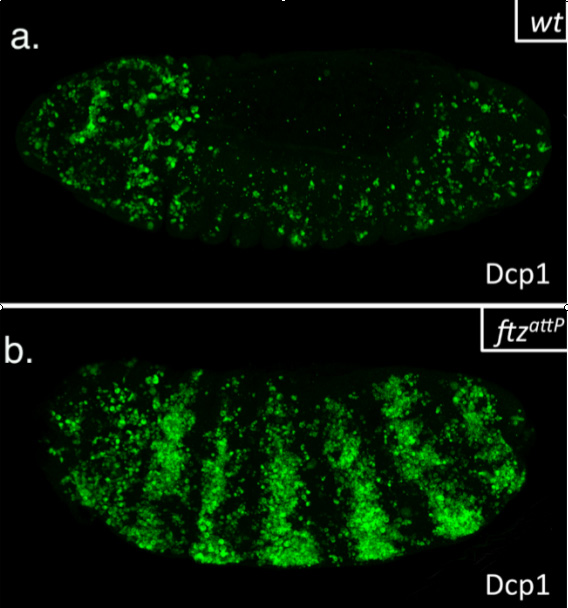
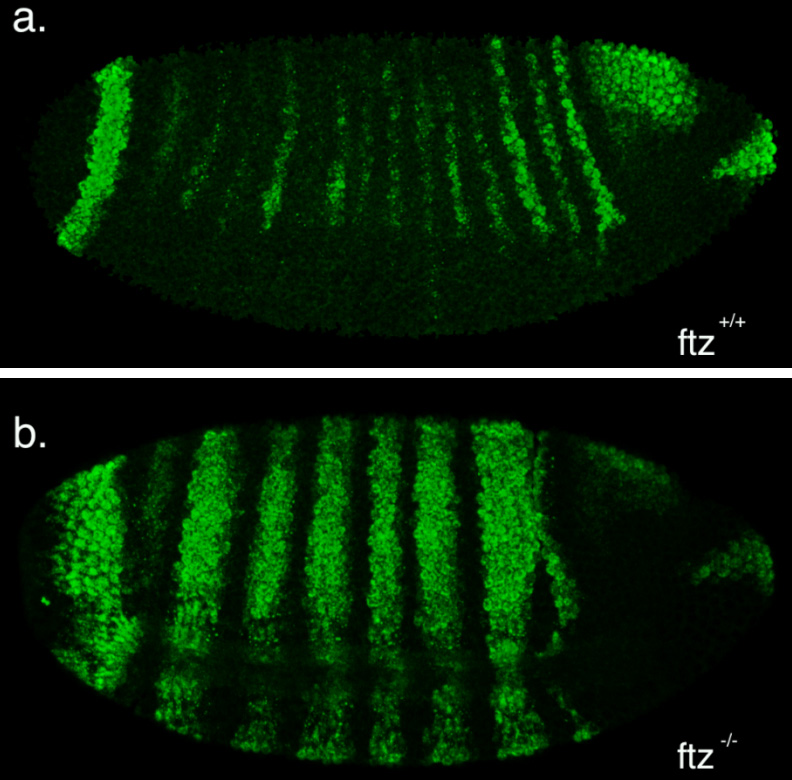
This summer, I had the glorious opportunity of undertaking a BSDB funded research project in the laboratory of Jean-Paul Vincent at the Francis Crick Institute in Mill Hill, London. My project took place in the lab of Dr Jean-Paul Vincent (www.jpvincentlab.com) under the patient and inspiring tutelage of one of his PhD students, Sam Crossman. Our project investigated the mechanism of apoptosis in the model organism Drosophila melanogaster.Apoptosis is a form of programmed cell death and is an important process in the development of all multi-cellular organisms. Understanding why cells die in certain situations and not in others is of relevance to many areas of health, including embryological disorders and cancer, and my overall research aim was to investigate the role of apoptosis in the developing fly embryo. To do this, I worked with strains carrying mutations in genes required for patterning the anterior-posterior axis. Mutation of these so-called patterning genes can trigger extensive apoptosis in the embryonic epidermis (figure. 1) and therefore provides a useful model to investigate the apoptotic machinery in Drosophila.
Figure 1: Increased apoptosis is observed upon the mutation of many patterning genes. Minimal cell death is observed when wild type embryos are stained with the apoptotic marker cleaved-Dcp1 (a), whilst embryos mutant for the patterning gene fushi tarazu (ftz) have increased apoptosis (b).
The cause of the ectopic apoptosis observed in patterning mutant embryos is not fully understood. One previously suggested explanation is that the cells of the epidermis can sense their ability to adopt the correct fate and undergo apoptosis if they lack the required patterning inputs to do so (Werz et al, 2005). However, if this were the case, there would have to exist an unknown machinery that would allow individuals cells to detect patterning errors and initiate apoptosis as a result.
In order to determine if cells are truly capable of detecting patterning errors, I planned to use a light inducible form of Cre recombinase to clonally remove a lox flanked allele of the ftz gene in a small subset of cells within each segment. If these small clones survive in an otherwise wild type embryo, it would argue against a cell-autonomous system where individual cells monitor their ability to differentiate correctly and would suggest that an alternative mechanism could be in play.
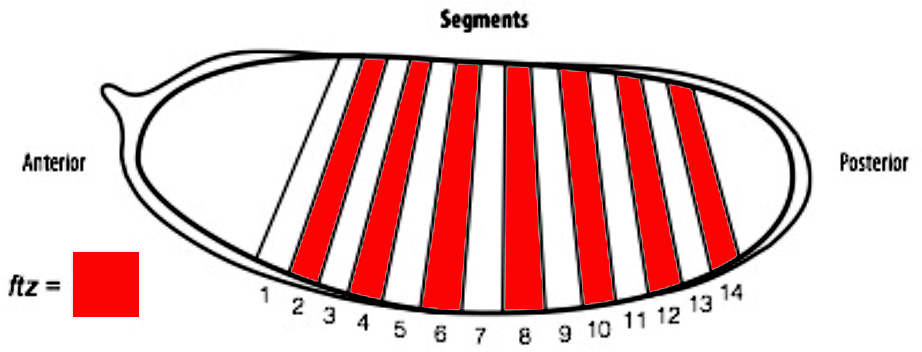
Figure 2. A diagram of a Drosophila embryo with 14 segments subdividing its anterior-posterior axis, which is achieved through the expression of patterning genes.
One difficulty with this plan is that ftz is activated very early on in embryogenesis. As a result, I set out to generate an early acting form of Cre, which could be used to remove ftz before it has carried out its function. To achieve this I spent the first part of my project cloning the Cre enzyme into a plasmid containing the actin promoter. As actin is an important protein in every cell, it is expressed from very early stages of embryogenesis. As a result, we hoped that by using the actin promoter to drive expression of Cre, we could produce the enzyme early enough to remove our lox flanked ftz allele in a timely manner and create mutant cells.
Unfortunately, cloning proved frustrating. Fortunately, it also proved educational. I learnt a lot about the way that experiments work, and how progress in science is more staccato than smooth. One issue I faced was the purification of my final plasmid using an Invitrogen maxiprep column. Each time I purified the plasmid I ended up with a lower yield than required, as I needed enough DNA to send to a company that would use it to generate a transgenic fly. I adjusted a parameter each time, but in the end it may have just been a plasmid with a low copy number as during my last attempt I purified it from a much larger bacterial culture, which finally gave me a sufficient amount of DNA.
The second part of my project was spent optimising a fluorescent in-situ hybridisation (FISH) protocol. I planned to use FISH to label cells expressing the pro-apoptotic gene hid in a series of patterning mutant embryos to characterise the regions where cell death occurs. As a control, I first conducted the protocol with a probe against the segment polarity gene wingless, which is expressed in a row of cells in every segment. This probe was made by someone else in the lab and is known to work, so I used it to learn the steps of the in-situ protocol.
Figure 3: Fluorescent in-situ images of wild and mutant ftz embryos using a wingless probe. a) a wild-type embryo with fourteen stripes; b) a mutant ftz embryo with seven stripes.
My FISH experiments with the wingless probe worked like a treat (figure 3), but difficulties soon followed when I attempted to make a new probe to label cells expressing hid. The stainings I conducted with the hid probe I had made repeatedly failed to work, and every attempt to generate a new probe proved unsuccessful. Disappointingly, I reached the end of my project before I managed to develop a protocol that worked, but I at least learnt plenty of science along the way!
My time in J.P.’s lab has been great for a number of reasons. I’ve come to love the problem solving nature of science and the freedom you get to explore what really interests you. It really is an adventure! However, I’ve also realised how difficult a career in science can be. Whilst this project has inspired me to become a scientist, I wonder if there’s a route into academia that would better suit my background as a medical student and my wider interests in medicine.
Sources
(1) Werz C, Lee TV, Lee PL, Lackey M, Bolduc C, Stein DS, Bergman A. Mis-specified cells die by an active gene-directed process, and inhibition of this death results in cell fate transformation in Drosophila. Development, 2005. 132(24): p. 5343-5352.
Applications are invited from highly motivated and creative individuals who are interested in studying fundamental mechanisms of neuronal migration and axon guidance. The main focus of our research is to understand the molecular and cellular mechanisms underlying the development of neural circuits using the embryonic spinal cord as a model system (http://www.ucmm.umu.se/english/research/sara-wilson/). The fellowship is currently funded for two years and is available immediately. The laboratory is located at the Umeå Centre for Molecular Medicine (UCMM), Umeå University, Sweden. UCMM is an interdisciplinary department, which focuses on questions in basic medical sciences and developmental biology and provides an interactive modern environment with good core facilities.
Requirements: Individuals with a background in developmental biology, neuroscience, molecular and cell biology or related discipline and with a keen interest in developmental neuroscience are encouraged to apply. The successful candidate will have or about to receive a Ph.D. in a relevant discipline and be proficient in written and spoken English.
Technical experience with imaging, vertebrate embryonic model systems – especially chick or mouse electroporation, mouse handling and genetics is a big advantage although training will be given. Experience with molecular, cellular and/or evolutionary biology will be positively considered. The most successful candidate will have a high level of motivation, be creative, organized and rigorous and have the ability to work both independently and within a team.
Please submit your application (reference 2015SW100) by 16th December 2015 to sara.wilson@umu.se by sending the following documents as pdf files:
1) A short cover letter (not more than 1 page) to include a description of your research experience and suitability for the position.
2) Curriculum Vitae including: publication list, technical expertise, names and contact information for three referees.
Informal enquiries may be directed to Dr. S.I. Wilson (sara.wilson@umu.se).
As we develop from wads of cells to fully formed humans, each of our organs goes through intricate processes to achieve the right combination and number of cells arranged in the proper way.
Research published in PLoS Biology by Yung Hae Kim and her colleagues looks at the development of our hormone-oozing pancreas, which assists digestion. They wanted to know how the cells become different cell types. Cells in developing organs begin without specificity and over time turn into various cell types upon receiving the right signals.
The beauty of this research, a part of the puzzle of how we form, is encompassed in the video below. This moving three-dimensional image is really hundreds of two-dimensional images merged together. The cells shown are a chunk of the pancreas, lit up in white, green, blue, and magenta. Our cells are constantly churning out proteins that they use to carry out their functions. It is these proteins that the colours are marking. And the colours carry meaning. Blue and magenta indicate two different kinds of cells, distinguished by a specific protein that only that type of cell makes.
From this colorful three-dimensional clip, Kim and her team looked at whether the cells of the pancreas are ones that can produce hormones, or cells that have not specialized to this function. Further down the line, knowledge about when and how the cells become hormone-secreting ones could be important for diabetes research. It is cells of the pancreas that produce the insulin we need to control blood sugar levels. Perhaps we could incite cells that haven’t specialized to make insulin, to secrete it.
This research is from the lab of Anne Grapin-Botton, Danstem, part of the HumEn consortium, working towards making fully functional beta cells for diabetes therapy.
 (2 votes)
(2 votes)




 (No Ratings Yet)
(No Ratings Yet)