Here is October’s round-up of some of the interesting content that we spotted around the internet!
News & Research
– Developmental Biologist Eric Davidson, who was based at CalTech, has sadly passed away. You can learn more about Eric and his career in this Q&A published in Current Biology a few years ago.
– Interesting piece in Nature News considers how biohackers (amateur biologists) are using CRISPR.
– Masayo Takahashi is the winner of the inaugural Ogawa-Yamanaka Stem Cell Prize, awarded by the Gladstone Institutes. Also don’t miss the chance to vote for the 2015 Stem Cell Person of the Year in Paul Knoepfler’s blog.
– How to unboil an egg and how to make a chicken walk like a dinosaur are just some of the winners of this year’s Ig Nobel prizes.
– How scientists fool themselves and how they can stop- preventing bias in science, in Nature.
In multicellular animals and plants, the single-celled zygote develops into the embryo. In the model plant Arabidopsis thaliana, the zygote divides asymmetrically to form a small cytoplasmic apical cell, which is the precursor of the proembryo, and a large vacuolated basal cell, which develops into the extra-embryonic suspensor (Figure 1). Communication between the intra-embryo (proembryo and suspensor) and the inter-embryo (embryo and surrounding tissues) is required for proliferation, pattern formation, and differentiation during embryogenesis1-3. Moreover, it has long been known that in several plant species, including Arabidopsis, disruption of the original embryonic tissue induces formation of newly embryonic cells derived from the extra-embryonic tissue4-6. However, the process by which an embryonic cell is initiated from the extra-embryonic tissue remains unclear. Angiosperm embryogenesis in vivo occurs within multiple layers of maternal tissues within the flower; therefore it is difficult to observe and analyze the intercellular communications in living material in real time. The development of a live-cell imaging system has been long-awaited to reveal how cell fate is specified by intercellular communication during early embryogenesis. Recently, we have developed a live-embryo imaging system to visualize cell division and cell-fate specification in Arabidopsis thaliana. We incorporated two advanced technologies—micro-engineering and optical manipulation—into this system.
Figure 1. Schematic representation of embryogenesis within Arabidopsis flowers. After double fertilization, the embryo and endosperm develop in the ovule. The zygote divides asymmetrically into an apical cell (embryonic cell) and a basal cell (extra-embryonic cell). The apical cell divides vertically to generate the proembryo, which ultimately gives rise to the adult plant. The basal cell divides horizontally to produce the suspensor (which acts like an umbilical cord in mammals), and finally undergoes programmed cell death.
First, we developed an in vitro ovule culture system so that time-lapse imaging could be performed. An in vitro culture system was necessary for real-time observation because embryogenesis could not be observed in vivo on account of the deep tissues. However, we could not observe any cell division in the isolated early embryo7. Therefore, we cultured the embryo within the maternal tissue—the ovules—in vitro. We used liquid medium to prevent the ovules drying up during long-term culture and placed the ovules close to the cover glass. However, stable observation of the ovules in the liquid medium was difficult because of their round shape, so we fabricated PDMS [poly(dimethylsiloxane)] microdevices to adjust the orientation and fix the position of the ovules using micro-engineering techniques.
PDMS has several advantages in the field of developmental biology, such as a high level of biocompatibility, including efficient gas exchangeability and a suitable transparency for observation. PDMS-based microfluidic devices have also been used for animal cell cultures8. Recently, plant researchers have also started to use PDMS-based microdevices for analyzing the growth of root tips, pollen tubes, and seedlings as a lab-on-a-chip9. Initially, we fabricated a PDMS microcage array to fix the ovules in the liquid medium10. To increase the trapping efficiency of the ovules, we modified the PDMS microcage array to a micropillar array11 (Figure 2). Micropillar arrays have a weaker effect on ovule growth because of the physiological flexibility of PDMS micropillars. Thus, we were able to conduct time-lapse imaging of cell division—from the zygote to the heart-stage embryo—for 3 days within the ovules in Arabidopsis embryogenesis.
Figure 2. Arabidopsis ovules on a PDMS micropillar array.
Another advanced technique is optical manipulation. To elucidate the mechanisms of cell–cell communications within the tissues, it is important to carry out a spatio-temporal analysis to determine when, where, and which cells are communicating. Optical manipulation is a powerful tool for such real-time analysis. Because lasers have high optical intensities at the focal point, laser ablation has long been used to disrupt cells. In plants, UV lasers have been used for laser ablation12,13. However, there are limitations to how UV lasers can be used. Short-wavelength lasers, such as UV lasers, are susceptible to refraction, absorption, and scattering from the tissues. Therefore, it is difficult to ablate the cell at the level of a single cell within deep tissues.
Recently, near-infrared pulse lasers have been gradually used to overcome the limitation of UV lasers. Near-infrared pulse lasers are tolerant to scattering within the tissues because of their longer wavelength. In addition, femto-second pulse lasers can induce nonlinear optical effects, such as plasma formation, at a sub-femto-liter volume. Thus it can induce damage even within a cell14. Hasegawa et al. performed the disruption of actin filaments within a tobacco-cultured cell using a femto-second pulse laser in real time15.
We performed laser disruption to our live-embryo imaging system using a femto-second pulse laser11. When an apical cell (embryonic cell) was disrupted by laser irradiation, the cell fate of the remnant basal cell (extra-embryonic cell) was converted, through changes in identity gene expression, into an embryonic cell (Figure 3). In terms of the disruption of basal cells, we found that apical cells were not converted into basal cells. Thus, we succeeded in performing real-time observation of cell-fate conversion for the compensation of an embryonic cell. We propose that the fate and behavior of each individual cell in Arabidopsis embryos is determined via intercellular communication with neighboring cells, and that such flexibility contributes to the robustness of plant embryogenesis.
Figure 3. Time-lapse images of embryonic marker, DRN::erGFP (green, ER localization) after laser irradiation of the apical cell. When an apical cell was disrupted (asterisk), the basal derivative cell underwent DRN expression and divided like an apical cell. Scale bar, 20 µm.
Our imaging system enables direct visualization of the dynamics of cell-fate specification during embryogenesis using micro-engineering techniques. In plants, cell fates are specified via positional information during development. Femto-second laser irradiation, as a form of optical manipulation, enables a spatio-temporal induction of cell-fate conversion that is impossible with genetic ablation. Thus, these techniques will enable novel insights into the molecular mechanisms of cell-fate specification via intercellular communication in plant development. Moreover, these techniques are highly suited to research into developmental biology as well as to plant research, so will be of interest to broad developmental biologists in addition to their applicability to live-cell analysis.
1 Kawashima, T., & Goldberg, R. (2010). The suspensor: not just suspending the embryo Trends in Plant Science, 15 (1), 23-30 DOI: 10.1016/j.tplants.2009.11.002
2 Javelle, M., Vernoud, V., Rogowsky, P., & Ingram, G. (2011). Epidermis: the formation and functions of a fundamental plant tissue New Phytologist, 189 (1), 17-39 DOI: 10.1111/j.1469-8137.2010.03514.x
3 Lafon-Placette, C., & Köhler, C. (2014). Embryo and endosperm, partners in seed development Current Opinion in Plant Biology, 17, 64-69 DOI: 10.1016/j.pbi.2013.11.008
4 Haccius, B. (1955). Experimentally Induced Twinning in Plants. Nature176, 355-356, doi:Doi 10.1038/176355a0
5 Yeung, E., & Meinke, D. W. (1993). Embryogenesis in Angiosperms: Development of the Suspensor Plant Cell, 5 (10), 1371-1381 DOI: 10.1105/tpc.5.10.1371
9 Sanati Nezhad, A. (2014). Microfluidic platforms for plant cells studies Lab on a Chip, 14 (17), 3262-3274 DOI: 10.1039/C4LC00495G
10 Park, J., Kurihara, D., Higashiyama, T., & Arata, H. (2014). Fabrication of microcage arrays to fix plant ovules for long-term live imaging and observation Sensors and Actuators B: Chemical, 191, 178-185 DOI: 10.1016/j.snb.2013.09.060
11 Gooh, K., Ueda, M., Aruga, K., Park, J., Arata, H., Higashiyama, T., & Kurihara, D. (2015). Live-Cell Imaging and Optical Manipulation of Arabidopsis Early Embryogenesis Developmental Cell, 34 (2), 242-251 DOI: 10.1016/j.devcel.2015.06.008
12 van den Berg C, Willemsen V, Hendriks G, Weisbeek P, & Scheres B (1997). Short-range control of cell differentiation in the Arabidopsis root meristem. Nature, 390 (6657), 287-289 PMID: 9384380
13 Higashiyama, T., Yabe, S., Sasaki, N., Nishimura, Y., Kuroiwa, H., & Kuroiwa, T. (2001). Pollen Tube Attraction by the Synergid Cell Science, 293 (5534), 1480-1483 DOI: 10.1126/science.1062429
14 Tirlapur, U., & König, K. (2002). Targeted transfection by femtosecond laser Nature, 418 (6895), 290-291 DOI: 10.1038/418290a
15 Hasegawa, J., Higaki, T., Hamamura, Y., Kurihara, D., Kutsuna, N., Higashiyama, T., Hasezawa, S. & Matsunaga, S. (2014). Vacuole subdivision in plant cell growth by the genotoxic stress inducing DNA double strand breaks. Cytologia79, 467-474, doi:10.1508/cytologia.79.467
This Sticky Wicket article first featured in Journal of Cell Science. You can read the Mole’s visit to the first 5 circles of Grant Hell here. Read other articles and cartoons of Mole & Friends here.
Mole was revising his rejected grant and moaning about how unfair it all is, when the shade of Francis Bacon, the Father of the Scientific Method, appeared to take him on a tour of Grant Hell. Sir Francis (‘Frank’) looks like Dumbledore but talks like a wanna-be hipster but, hey, maybe Mole is just dreaming all this. Anyway, they went to Grant Hell. There, they met lots of wretched souls who were not successful in revising their grants, and had lots of reasons why not.
Oh, one more thing. For some reason, the story is told in triplets that don’t rhyme. Probably they did in some other language, but it would have been too much trouble to translate it and make it rhyme. Mole may be getting lazy. Or maybe he’s up to something? No, most likely he’s just lazy. We join our travelers, having passed the Fifth Circle, and have continued up to the Fourth.
“Here is the Fourth Circle,” said Frank. This was Greed.
“But hey,” I said, “I thought greed is good. That’s what they tell us,
It makes the market work.” Frank shook his head.
“Wait,” I said, “Are you saying that these poor people are here
Because they wanted to do too much? They wanted too many grants?
I mean, what’s wrong with a little ambition if the science is good?
“Listen,” I went on, “I’m getting a little tired of hearing that somebody, who
I think is a really terrific scientist, has ‘too many grants.’ I mean, if they are
Getting the best work done, maybe they deserve the grants. You know?”
“And now,” I think I was on a roll. “Now you tell me that they are here,
In Grant Hell, because they work too hard.”1 “Easy, Big Guy,2” said Frank,
And I looked to see if he was talking to me. He was. Nobody calls me ‘Big Guy.’
He went on. “You’ve got it all wrong. They aren’t here because they wanted
Too many grants. They are here because they confused grant funding and power.
Their greed is not to do more great science but to amass influence.”
“Oh,” I said. “Then they get what they deserve.” Because being ‘powerful’
As a research scientist is a bit like being the King or Queen of Cheese.3
It isn’t worth very much and can smell pretty terrible. Frank cracked up at that one.
We had moved to the Third Circle, where the wretched never stopped writing,
Except to eat the paper they were generating. Or electrons, but I couldn’t see that.
Then they would write some more. “Here is Gluttony,” whispered Frank.
I stopped one poor soul who was eating a page as it came out of a printer.
“Why are you doing this?” I asked. “Oh,” he moaned. “You know how it is,
I don’t get my grants funded, so I write more and more and more.
“I was putting in a grant every cycle, but that didn’t work, so I went to two,
Then three. I don’t have anything new to say, and I spend all my time
Regurgitating my ideas and sending them in. Eventually I’ll get one.” He belched.
“But wait,” I cried, “Maybe you need to stop and actually think about what
You are writing! Maybe if you took some time out, you’d frame the work
In a way that reviewers will see why it is important?” He wolfed down a page.
“No, this is how I do it,” he sputtered between mouthsful. “This is how I roll.”
“How’s that working out for you?” I asked, but he wasn’t interested.4
“Let’s go up,” said Frank. “Not much more to see here.”
We found the Second Circle. Souls were flying in the winds that blew around us.
“This is Lust,” said Frank, watching them flit around us. “Their only crime, I think,
Is to want their grant too badly. They don’t want to change it, because they love it.”
I could understand. Maybe this was my own circle. I really, really want my grant.
I lust for it. The work is lovely, and elegant, and important. But it wasn’t funded.
I could feel myself starting to float off the ground and my clothes began to flap.
“Whoa, there, Compadre,5” said Frank, grabbing my leg and bringing me to earth.
“Get a grip. I mean, its just a proposal for what you want to do. There’s nothing
of value in a grant, except as a way to get the support for the work, right?”
I wasn’t so sure. I mean, when I write a grant, I think about the project,
Really think about it, often for months. The act of writing helps me to formulate,
Consolidate my thoughts. It’s a plan for the next few years of research.
But yes, he was also right. The act of writing a grant, really, is just an approach
To supporting the work that has been formulated. Yes, thinking about it is good,
But actually doing the work is what this is all about. I needed to think.
And found that we had entered the First Circle. Limbo. Nothing to do but wait.
And while we all waited to hear the results of our submission, we were happy.
Because until we get the bad news, we can imagine that we’ll get it.
We emerged in the light of Hope. And there, before us, was a mountain.
“We’ll climb that in a while,6” said Frank. “But first, I know a good place
Near here to get some lunch. You down for that?” Me, I didn’t know down from up.
Footnotes
↵1 Mole is correct here, and despite FB’s words to the contrary, many experienced investigators have their grants fail because of a perception that “they have too much money.” Indeed, funding institutions sometimes place limits on total amounts a program may have, and in those that do not, the reviewers may take this position themselves. But Mole feels that this is inappropriate; some investigators are simply more productive than others and, given the opportunity, more support can result in substantially more impactful research from such investigators. Of course, the opposite is also true – there are investigators who will not be productive, regardless of how much support they have.
↵3 (il re o la regina di formaggio) It is possible that Mole is referring to either King Camembert (1411–1489) or, perhaps, Queen Gouda (1536–1590). Or he may be referring to his friend Weasel who is, indeed, very fond of cheese.
↵4 This approach, of flooding granting institutions with applications in hopes that one may “stick” is becoming much too common. Mole doubts that it is a viable strategy, unless such multiple applications can be done with the same care that goes into only one. As this is rarely the case, the strategy might, at least, be reconsidered. Given that the result is an avalanche of hastily prepared applications converging at any one competition, an exceptionally well-crafted (and well-reasoned) application might stand out and be favored. It’s just a thought.
↵6 The mountain, Grant Purgatory, eventually leads to Grant Paradise, which lasts only until the period of the grant runs out, and a renewal applied for. Mole points out that it may be interesting to note that, whenever a grant is rejected and we find ourselves in Grant Hell, we blame the process (“they killed me”). When it happens that we emerge into the light of funded Grant Paradise, though, we did it all by ourselves (“yes, I got the grant”).
Xenbase (www.xenbase.org) is the Xenopus bioinformatics and genomics resource. Xenopus is a major model for fundamental cell and developmental biology and a model for human disease. Xenbase is a totally free, and globally accessible database, used by Xenopus researchers worldwide, and is funded by the National Institute of Child Health and Human Development. Xenbase has two performance sites: the curation team is based in Cincinnati, OH (PI: Dr. Aaron Zorn) and the developer/database management team is based at the University of Calgary in Canada (PI: Dr. Peter Vize).
Xenbase is seeking to fill 2 full time Curation positions to join the curation team at the Division of Developmental Biology, Cincinnati Children’s Hospital, Cincinnati, OH, USA. Curation positions offer a challenging job away from the wet-lab and research bench, where interpreting, annotating and displaying complex data is our main task. Curators also develop strategies to improve data curation; work to improve data display/querying on the website; interact with our user community at research conferences; develop programming skills; and contribute to Xenbase publications.
Job Description:
Curation and annotation of published Xenopus scientific literature, focusing on gene expression and the extraction of other research data: genes, transgenics, antibodies, morpholinos, phenotypes, genetic interactions, gene product functions and models of human disease.
Import and annotate data from large-scale screens (e.g., loss-of-function morpholino screens, gain-of-function mRNA screens).
Help develop new features: curation and processing of public and directly submitted RNA-seq and ChIP-seq NGS data from Xenopus experiments, curation of mutant phenotypes and transgenics; expanding our an anatomy atlas; implementing GO annotations.
Co-author reports and publications, and give presentations at national and international meetings and workshops.
Qualifications:
MSc or PhD degree in bioinformatics and/or developmental biology, genomics, genetics, molecular biology, zoology, anatomy or related field.
Demonstrated ability to produce scientific papers, reports and presentations
Demonstrated ability to work in a team as well as independently, efficiently (i.e both quickly and accurately) and be self-motivated
Strong interpersonal and communication skills, including excellent written and spoken English
Preference will be given to applicants with:
Experience with a bioinformatics, genomics or model organism database
Experience in data annotation/biocuration, knowledge of relational databases, and familiarity with ontologies.
Experience in a Xenopus or other vertebrate (mouse, zebrafish or chick) developmental biology lab.
Experience in analyzing genomics data, using GRN software, genome browsers and common bioinformatics tools.
How to Apply:
Please submit your application, to: aaron.zorn[at]cchmc.org with the following information:
A cover letter , including a statement of interest/purpose
CV/Resume.
Copy of your degree(s).
List 3 references/referees whom we may contact (please include their postal address, email and phone number).
Salary and Start Date:
Salary will be commensurate with qualifications and experience. Start date is negotiable, but expected to be in early 2016.
The successful applicants will be employees of Cincinnati Children’s Hospital and will undergo background checks, orientation and a 3-month probationary period. Employees are required to receive an annual flu vaccination.
Using chick embryos we recently identified BMP/Smad and Wnt/GSK3b signals as extrinsic cues that control the migration of prospective cardiac cells. We showed that BMP and Wnt pathways converge on a common effector: the transcription factor Smad1 (Song, McColl et al., PNAS 2014). We will use systematic approaches to determine the targets of BMP/Smad1 and Wnt/GSK3b signalling in early mesoderm cells. Identified candidate genes will be assessed for their function in controlling cell migration and fate choice.
A NIH-funded postdoctoral position is available in the laboratory of Dr. Kevin Francis atThe Children’s Health Research Center at Sanford Research (Sioux Falls, SD) to study the consequences of altered cholesterol metabolism on neurodevelopment and neuronal function. The successful applicant will utilize a wide range of experimental approaches, including iPS cell models, electrophysiology, whole genome sequencing, confocal imaging, and genome editing, to answer questions of biological and clinical significance. Current research projects are exploring lipid-protein interactions critical for neurodevelopment, identifying mechanisms underlying neuronal fate choice, and developing iPS cell-based assays for high-throughput small molecule screens.
Candidates with established expertise in electrophysiology are particularly encouraged to apply. Experience in neurobiology, molecular biology, and stem cell biology is also desirable.
Additional information about the Children’s Health Research Center can be found on the center’s website here.
Contact: Candidates should submit a cover letter (including career goals), curriculum vitae, and contact information for three or more professional references to Dr. Kevin Francis at: kevin.francis@sanfordhealth.org.
We are happy to announce the forthcoming workshop on « Tissue mechanics in morphogenesis: Focus on theoretical modeling ». This is an informal workshop open to anyone, theorist or experimentalist, who is developing or is using theoretical models to understand tissue mechanics during morphogenesis.
How tissues acquire and maintain their shape is a crucial question at the crossroad between developmental biology and physics, which requires the joint efforts of experimentalists and theorists. Theoretical modeling plays different roles in this context: it guides experiments, helps data analysis and provides conceptual and predictive frameworks, which are instrumental to understand tissue morphogenesis and homeostasis.
Several models have been developed over the past years, some are published, and some others are in preparation.
This workshop aims at encouraging:
– discussions between researchers in the field
– exchange of published and unpublished information
– discussion of regime of applicability of theoretical frameworks
– link between different scales: molecule, cell, tissue
– studies of dynamical processes in morphogenesis
– new approaches to data analysis
– improvement of existing models
– development of new theory/models
– new collaborations
A postdoc position is available in Jim Smith’s lab at the Francis Crick Institute to study the role of PAWS1 during embryonic development and in cancer. PAWS1 regulates signalling by bone morphogenetic proteins (Vogt et al., Open Biology4, 130210; 2014), and we have recently demonstrated that it influences the activity of additional signalling pathways and modulates a range of different cellular functions, including ciliogenesis. See www.crick.ac.uk/jim-smith.
 (1 votes)
(1 votes)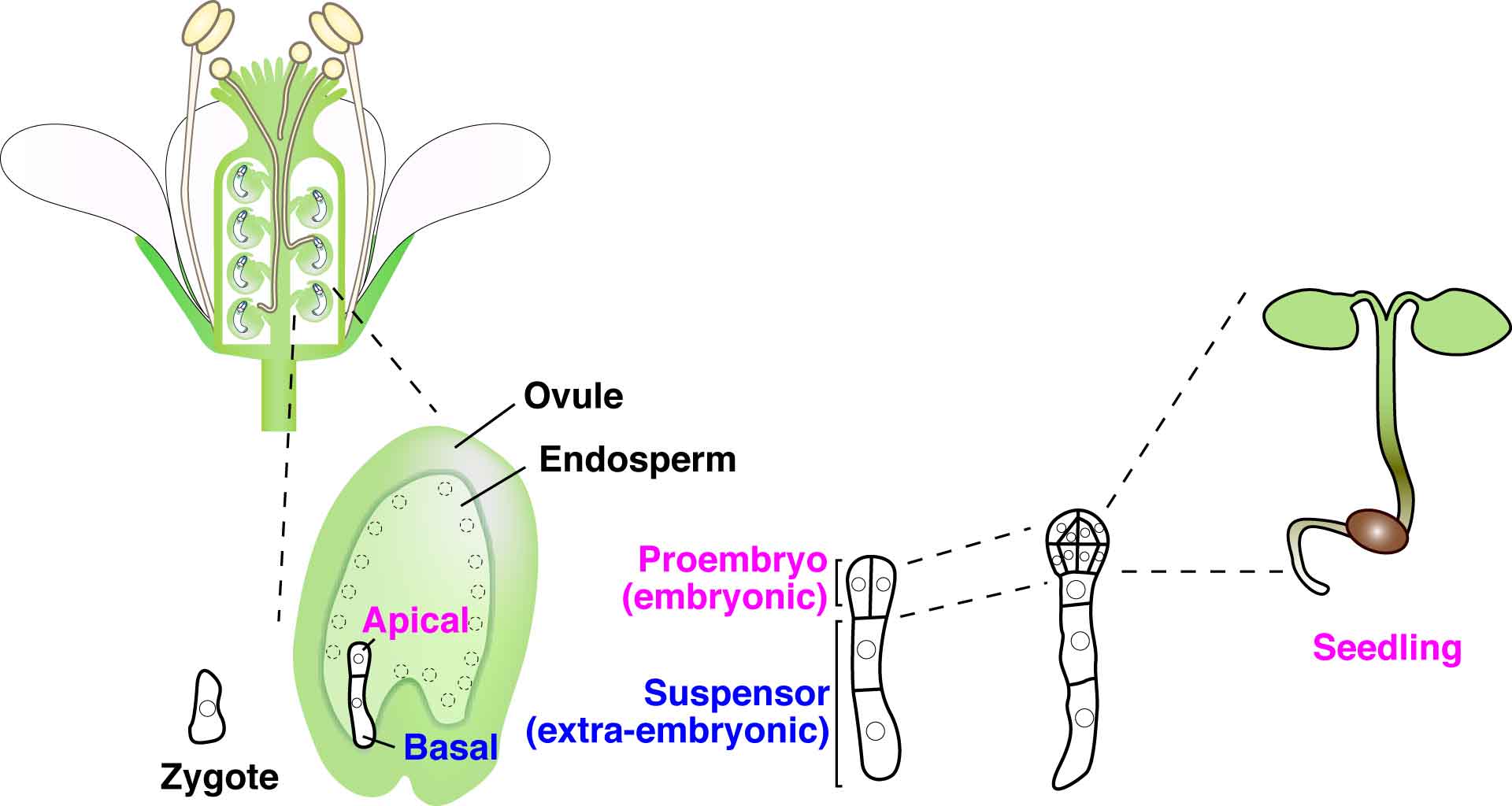



 (No Ratings Yet)
(No Ratings Yet)