The University of Chicago’s Department of Neurobiology seeks to recruit tenure-track faculty. Appointments can be made at any rank from Assistant Professor to Professor, and in any area of neurobiology. Candidates must have a doctoral degree and at least two years of postdoctoral training.
Researchers in molecular/cellular neurobiology or neural development/plasticity are particularly encouraged to apply.
Successful candidates will benefit from a rich intellectual environment that includes a new Neuroscience Institute and an affiliation with the Marine Biological Laboratory in Woods Hole.
All interested applicants must apply online at: academiccareers.uchicago.edu/applicants/Central?quickFind=53616 by uploading a cover letter, a CV, a statement of research objectives, and the names and contact information of three academic references. Application review will begin October 1, 2014 and continue until the position is filled.
All qualified applicants will receive consideration for employment without regard to race, color, religion, sex, national origin, age, protected veteran status or status as an individual with disability.
The University of Chicago is an Affirmative Action / Equal Opportunity / Disabled / Veterans Employer.
This is the third of four posts relating to the Future of Research symposium which was announced in a previous blog post. Each of these posts will discuss a topic that is the focus of a workshop at the Symposium. Even if you can’t attend, please tweet @FORsymp with suggestions, or follow us to respond to our questions about what YOU, trainee scientists, think is important. The hashtag for this post on the Workforce workshop is:#FORmetrics
Rewards and incentives
What is “good science”? What is it that we want science to accomplish?
Does our current system reward “good science”, our shared goal, or are there perverse incentives that do the opposite? What are the problems and how could we reward scientists and institutions to produce the behaviors (such as collaboration, openness and honesty) that we believe support “good science”?
We all have ideas about what science is supposed to do/achieve, but these outcomes happen at a broad, communal, systems level. Meanwhile, rewards/incentives operate on individual scientists. Often, there is a disconnect between those two.
Many of the current incentives work on individuals or small groups of people (grants, jobs, fame, etc.), while many of the things we want science to do (cure disease, figure out how the world works, etc.) need to happen at a broader systems level. This may contribute to the disconnect between incentives and desired outcomes.
Intrinsic motivators may include curiosity, desire for freedom, etc.; the reasons people got into science in the first place. The extrinsic motivators are things like grant money, lab space, prestige, etc. How can we reinforce or encourage people’s own intrinsic motivations (at least the ones that lead to good science)?
The objective of the Metrics workshop is to ask: How can we fix the current system of incentives so that we reward scientists and institutions for the behaviors that we believe support “good science”?
Key areas where science “breaks”
Reproducibility and negative data
Part of the scientific endeavour is to provide checks and balances and reproduce results, or highlight when reproducibility fails. However, it is difficult to publish the results of replicating experiments or negative data and there is a worrying trend in the lack of reproducibility in some forms of analysis highlighted recently with regards to the widely-used technique of fluorescence-activated cell sorting (FACS, Hines et al., 2014). Some journals have made a call specifically looking for negative data and there are indications that the NIH may be looking to drive more studies reproducing data (Collins and Tabak, 2014) but more work on this area is key.
Publish or perish
Success in grant applications and career progression relies on publications (van Dijk et al., 2014). However this can lead to hyper-competition for “high-imapct” publications and in some recent, sad, cases, a lack of truth in publishing (Sovacoll, 2008; Nosek et al., 2012). Clearly the need to publish needs to be balanced with rigorous and honest scientific communication.
Other examples of broken systems
What can we learn from other folks who have tried to fix broken systems? For instance, the aviation industry has gone from a culture of “blame the dead pilot” to rigorous investigations of aviation accidents, which lead to improvements in the system for everyone. People like Atul Gawande in his book “Better” and Malcom Gladwell in “Outliers” have studied “positive deviants”, people who do their job much better than average in order to try to understand how we could all do better, and how we can change the system so that doing better becomes routine. Could we learn from hospitals that have used real patient involvement to improve safety and health outcomes, or from the aviation industry?
Hospitals using patient engagement to bring about real change have instituted a policy that every time a decision that will impact patients is made, a patient representative will be at the table (including who to hire, what equipment to buy, pricing issues, etc.). In science/universities, who are the stakeholders? Who should be at the table when decisions are made?
Where are the dark corners? Where do we hide our embarrassing failures? Examples could include things like replications failures or professional failures like honesty slip-ups. In the cockpit, everyone is responsible for the safety of the aircraft, and anyone who sees a problem is supposed to speak up; formerly, the all-powerful pilot was not supposed to be questioned. In medicine, whistle-blowing has improved patient outcomes by changing the culture so that everyone in the room is responsible for the outcome and it’s expected for those lower down the power curve to speak up if they see something. In science, how could we learn to talk openly about our mistakes without losing professional standing? How could we reward people for saying “I think (or know) that our paper was wrong/misleading/incomplete”?
Questions – please give us feedback!
How would we like scientists to behave, and what do we think science is supposed to achieve?
What are the current rewards and incentives, and what kind of behaviors and outcomes do they promote?
What else could we do to promote or reward the behaviors/outcomes we’d like to see?
What are the best parts of your job as a scientist? How could the system be changed so that you spend more of your time doing those things?
What prevents you from doing your science and doing it well?
How can we change the culture? How can we all agree on what the goal is?
This post has been written from input provided by the moderators of the workshop on “Metrics and Incentives in Science”.
This is the second of four posts relating to the Future of Research symposium which was announced in a previous blog post. Each of these posts will discuss a topic that is the focus of a workshop at the Symposium. Even if you can’t attend, please tweet @FORsymp with suggestions, or follow us to respond to our questions about what YOU, trainee scientists, think is important. The hashtag for this post on the Workforce workshop is:#FORworkforce
Where do postdocs go?
There has been a tremendous shift in the job market for PhDs/post-docs over the past decades. The only job that PhDs/post-docs are trained for (academic PI positions) are precipitously dwindling. Under such conditions, how do we match the changing job market demand with the supply of rightly trained PhDs/post-docs?
There are high numbers of graduate trainees and postdoctoral researchers in the current academic research system (Alberts et al., 2014). There are now slowdowns, or even contractions, in the ability of academia, the government and industry to take on this excess number of postdocs. The goal of this workshop will be to figure out what the best ways of adapting the system to best reduce the postdoc pool (Bourne, 2013).
As discussed in a prior post, the assumption is that everyone who goes into academia wants to end up an academic. Expectations have been shown to change over time as a trainee progresses, in spite of strong encouragement from advisor, who actively discourage other career paths (Sauermann and Roach, 2012). And the jobs people are actually getting show that, actually, academia is one of the “alternative” careers, and not the default. So the workforce becomes filled with more trainees than are needed to replace current academics.
What is the cause of this mismatch?
The doubling of NIH budget in 1990s made provision for a huge influx of money – particularly soft money which the academic institutions used up for massive expansion (of a large number of PhDs, post-docs, etc.) without taking into consideration the long term effects of this one-time influx. The result is over the past decade, in conjunction with the economic crisis and sequestration, the NIH budget has contracted by 20 %, essentially leaving the trainees high and dry without good job prospects for the (academic) jobs that they were trained for.
What is the current demand on the workforce encompassing the grad students and post-docs?
Although almost all PhDs and post-docs are trained, or directed, to become PIs in academia, the current reality is that these academic jobs account for less than 15 % of the job market. Due to The Great Recession, jobs in industry are also in a downward flux. However, there are many new job opportunities that are opening up in fields that initially were considered alternative careers and looked down upon. These include: consulting for life sciences and pharmaceutical industries; sales; marketing and field specialists of high tech and technologically advanced products; science policy; communications; patent law and intellectual property; and many more.
There is a perception that there is actually a shortage of Science, Technology, Engineering and Math (STEM) graduates in the United States. However a recent report by the Center for Immigration Studies using US Census data (Camarota and Ziegler, 2014) is one of a chorus of recent publications suggesting that this is in fact not the case, and that STEM graduates are actually struggling to get jobs. Are we producing too many STEM graduate or too few? Bizarrely, the answer may be “both”, as recently discussed, and that we simply have too many graduates, who are trained for the wrong careers.
Questions – please give us feedback!
How did we get to this situation and what is perpetuating the problem?
What are possible ways of changing the structure of the workforce to relieve the pressure?
This post has been written from input provided by the moderators of the workshop on the “Structure of the Workforce”.
We invite applicants for a tenure-track faculty position in Developmental Biology. We welcome colleagues whose research activities will complement our existing focus areas in neurosensory systems, neurodegeneration, cancer and other diseases, epigenetics or pathogenesis. Potential model systems of interest are mouse, zebrafish, fruit fly or stem cells to study development and disease. The successful applicant is expected to address fundamental questions in developmental biology using varied experimental approaches such as advanced imaging, biochemistry, genomics, genetics and/or gene transfer for translational studies. The position requires teaching graduate and undergraduate courses, mentoring research students and contributing to the mission of the department.
Applicants must have a Ph.D. in Biological Sciences or related discipline and at least 2 years of postdoctoral experience. We expect to fill academic year appointments at the Assistant Professor level, but appointments at the Associate level will be considered.
The Department has over 50 faculty conducting research in a wide range of fields (www.bio.purdue.edu/). We are committed to the success of all new faculty: we assign faculty mentors and offer competitive startup packages. Opportunities abound to use advanced research facilities across campus, such as the Bindley Bioscience Center (www.purdue.edu/discoverypark/bioscience/).
Applications must be submitted electronically to http://hiring.science.purdue.edu as a PDF with a detailed curriculum vitae, contact information for three referees, a 2-3 page summary of research interests and a teaching statement. A clear description of your major research accomplishments and future plans is more valuable for our evaluation than the impact factors of the journals in which you have published. Direct inquiries to search@bio.purdue.edu. Review of applications will begin October 1, 2014 and continue until the position is filled. A background check will be required for employment in this position. Purdue University, West Lafayette, IN is a dual career friendly employer.
Purdue University is an EEO/AA employer. All individuals, including minorities, women, individuals with disabilities, and protected veterans are encouraged to apply.
Postdoctoral positions are available in the laboratory of Systems Stem Cell and Developmental Biology (PI Aryeh Warmflash) at Rice University (Lab website) . We use human embryonic stem cells as a model for studying cellular communication, self-organization, and spatial patterning during embryonic development. The lab utilizes a combination of quantitative experimental techniques and mathematical modeling to address fundamental questions in stem cell and developmental biology (for more details on the work, see Warmflash et al. Nature Methods 11:847, 2014; Sorre et al Developmental Cell 32:332, 2014).
Rice provides an exciting atmosphere for working at the interface between biology and physics with the Center for Theoretical Biological Physics and a new graduate program in Systems, Synthetic and Physical Biology hosted on campus as well as ample opportunity for collaboration with faculty at Baylor College of Medicine and MD Anderson Cancer Center.
Applicants should hold a PhD in Biology, Physics, or other related field by the time of employment, have a track record of success in research, and a strong desire to work at the interface between quantitative and biological sciences. Applications from both experimental and theoretical scientists are welcome. Interested applications should send a one page letter highlighting previous research accomplishments and future research interests and a CV to Dr Aryeh Warmflash aryeh.warmflash [at] rice.edu
Waddington, whose writings on the epigenetic landscape continue to influence developmental biology to this day, called the developing embryo “the most intriguing object that nature has to offer”(Waddington, 1966). The mechanisms of pattern formation and morphogenesis have fascinated biologists for centuries. One question that is difficult to answer is what are the minimal requirements for generating an embryo? To ask a more specific question, at gastrulation stages, the cells of the mammalian embryo separate into three germ layers in a precise spatial arrangement. Is the information for this arrangement present in the cells themselves or do they need cues from the surrounding environment? With the ability to culture mammalian embryonic stem cells (ESCs), we can now begin to tackle these questions by asking how much of embryonic development we can recapitulate with ESCs in a culture dish.
Nearly every paper on ESCs touts their potential for regenerative medicine and there is a large literature that seeks to differentiate ESCs to pure populations of useful cells for just this purpose(e.g. Chambers et al., 2009; D’Amour et al., 2005; Kattman et al., 2011; Ozair et al., 2012). However, embryos, and by extension ESCs, are not in the business of making pure populations but have evolved to self-organized into patterns. Thus, given the right conditions, it is possible that embryonic patterns can be generated spontaneously in culture starting from ESCs. In a recent paper in Nature Methods(Warmflash et al., 2014), we asked whether the embryonic organization of the germ layers in the very early embryo can be achieved in a culture dish beginning with pluripotent human cells.
To begin to explore this idea, we began by simply adding BMP4, a part of the embryonic signaling cascade that induces gastrulation and patterning(Arnold and Robertson, 2009), to cultures of human ESCs grown under standard conditions. The cells readily differentiated, however, the results lacked reproducible organization at the level of fate. This negative result raised the question – why do ESCs fail to generate reproducible patterns in contrast to their embryonic analogs? One difference between the embryonic and culture environments is that the embryo provides a reproducible geometric structure while typical stem cell colonies take on arbitrary sizes and shapes (Figure 1A). We thus aimed to see how much of embryonic patterning could be restored simply by controlling the colony geometry (Figure 1B). To do so, we used micropatterning technology to grow cells on 1mm or smaller patches of cell adhesion molecules (Matrigel or Laminin) surrounded by a lawn of an inert substance to which cells and protein will not adhere. While such assays had been previous employed to evaluate the effects of varying colony size on cell fate(Bauwens et al., 2008), this work did not report any spatial structure in the differentiated colonies. To our astonishment, simple geometric confinement was sufficient to almost fully restore patterning. Two days after the cells were induced to differentiate with BMP4, four distinct cell populations emerged in concentric rings along the axis of the colony. From the outside in, these layers are extraembryonic, endoderm, mesoderm, and ectoderm (Figure 1C). This is precisely the arrangement found in the mammalian embryo at the conclusion of gastrulation.
A. Scan of a large region of a standard hESC culture showing variable colony organization. B. Scan of a large region of micropatterned hESC culture. C. A single micro patterned colony (corresponding to the largest colonies in B) stained for markers of trophectoderm (Cdx2), mesoderm(BRA), and ectoderm (Sox2).
A. Scan of a large region of a standard hESC culture showing variable colony organization. B. Scan of a large region of micropatterned hESC culture. C. A single micro patterned colony (corresponding to the largest colonies in B) stained for markers of trophectoderm (Cdx2), mesoderm(BRA), and ectoderm (Sox2).
Since the differentiating signal is added at high levels to the medium, it is presented homogenously to all cells. Thus, the presence of four cell-fate territories implies that induced signaling between cells overwhelms exogenously supplied signals. How do cells use these induced signals to define position inside a colony 1 mm in size and containing approximately 2000 cells? A clue emerged from examining smaller colonies patterned onto the same slide. The smaller disks lost the inner-most fate, but preserved the outer three at the same position as measured from the colony edge! Thus, cell fates are defined using the colony border as a reference.
What signals are responsible for this long ranged communication? The direct response to BMP4 is measured by the nuclear accumulation of the activated transcription factor Smad1. Persistent BMP signaling is only seen in a narrow ring at the border of the colony and specifies the extraembryonic tissue found there. Inside the colony, a broader ring of activated Smad2, an upstream component of the Nodal signaling pathway, is responsible for inducing mesendoderm. The inner ectoderm territory is defined by the absence of both signals. Several decades of work in developmental genetics have revealed the identity of the inhibitors that restrict BMP and Nodal signaling in the embryo. When the combinations of inhibitors that give a robust embryonic phenotype(Bachiller et al., 2000; Perea-Gomez et al., 2002) are knocked down, cell fate markers spread in the expected way. Thus the boundary is defined by the loss of BMP inhibitors, while Nodal inhibitors restrict the range of the mesendodermal territory. This conclusion is reinforced by differentiating the colonies in mini-wells with high walls: when the inhibitors are prevented from escaping, the entire colony adopts the central ectodermal fate.
Beyond simply providing a reliable geometric structure, colony confinement also has profound effects on the fates that form. Unpatterned colonies form mesoderm and extraembryonic tissue (consistent with previous studies on BMP4 differentiation) but largely fail to form endodermal and ectodermal tissues. We speculate that the increased cell density that results from confinement is necessary both to achieve high enough levels of Nodal signaling to form the endodermal region, and high enough levels of inhibitors to establish the ectoderm.
But what is the relevance of a circularly symmetric colony to an embryo? The ring of mesodermal cells defined by Brachyury expression becomes mounded and displays a collection of markers such as Snail and activated Erk that are characteristic of the primitive streak (PS). Thus the micropatterned colonies appear to form a radially symmetric PS region. This is reminiscent of mutant mouse embryos that are unable to initiate the formation of an anterior-posterior (AP) axis and show a proximal and radially symmetric primitive streak(Nowotschin et al., 2013).
These experiments raise many questions. In the mouse embryo, the signals that initiate AP axis induction reside at the distal end of the visceral endoderm, the extraembryonic epithelial layer, and move to the future anterior side during gastrulation. Will the same signals, presented locally to a micropatterned colony, restrict the PS-streak like region and induce an axis? How does endogenous signaling compete with exogenously supplied BMP4 in the culture medium? The effects of secondary signals can be directly examined by growing cells an imposed flow with microfluidics to remove such signals(Moledina et al., 2012; Przybyla and Voldman, 2012). It is typically very hard to study how signals spread in intact embryos at endogenous levels, so synthetic systems permit one to test hypothesis and then return to the embryo with more targeted experiments.
Much of what we know about the mechanics and biochemistry of signaling comes from cell culture experiments, which have at best only a temporal dimension. But the spatial patterns formed by morphogens and their secondary inhibitors are a biologically relevant phenotype, for which a simple in vitro assay has been lacking until now. Our system is very amenable to long term time lapse imaging, and it will be possible follow markers for cell fates as they emerge in time, and ask, for instance, whether the speckled patterns seen with immunofluorescence are merely cells following the same trajectories but at different rates. It is also an ideal system in which to quantify signaling in space and time and thus in a more predictive way engineer tissues and organs.
Waddington, C. H. (1966). Principles of Development and Differentiation. New York: Macmillan.
Warmflash, A., Sorre, B., Etoc, F., Siggia, E., & Brivanlou, A. (2014). A method to recapitulate early embryonic spatial patterning in human embryonic stem cells Nature Methods, 11 (8), 847-854 DOI: 10.1038/nmeth.3016
This is the first of four posts relating to the Future of Research symposium which was announced in a previous blog post. Each of these posts will discuss a topic that is the focus of a workshop at the Symposium. Even if you can’t attend, please tweet @FORsymp with suggestions, or follow us to respond to our questions about what YOU, trainee scientists, think is important. The hashtag for this post on the Training workshop is:#FORtraining
Who is a “trainee”?
By training, we are referring to the period between obtaining an undergraduate degree getting a first faculty-level position: graduate students and postdocs.
At this point in time, the common thread to training consists of the accumulation of research experience, culminating first in a dissertation for defending a PhD thesis, and then to produce science (most directly in the form of publications) during postdoctoral research, designed to be a more independent research environment under the scope of a Principal Investigator. The goal is then for a trainee to be able to find an academic position, beginning their own lab.
How many trainees are there, and how has this changed over time?
The NIH Biomedical Research Working Group Report (2012) makes for sobering reading. The proportion of PhDs moving into tenured or tenure-track positions decreased from 34 % in 1993 to 26 % in 2012, whilst the proportion of non-tenured faculty has remained constant but increasing in terms of actual numbers. The number of graduate students has doubled in the same time. Numbers of postdocs are harder to track, because nobody seems to, but estimates are that these numbers have also doubled (the NIH Biomedical Research Working Group Report implies that even this could be under-estimated by as much as a factor of 2). Research Training in the Biomedical, Behavioral, and Clinical Research Sciences, by the National Research Council (2011), suggests that as of 2011 there may have been just over 50,000 trainees in the biomedical research system.
How long is one a trainee?
In the US system, PhDs can take anywhere between 5 and 10 years. Postdoctoral training has been identified by the NIH as only being 5 years on average, but the number of people doing up to 8, or even 10 and beyond, has been steadily increasing in recent years. Some institutions put a cap on how long someone can be a postdoc, with a requirement to then make them a research associate.
What trainees expect, and where they end up
The assumption is that everyone who goes into academia wants to end up an academic. Is this the case? Expectations have been shown to change over time as a trainee, with a recent study showing that a faculty position tends to become less attractive over the course of a PhD, although by the end it is still the preferred career path for 50 % of trainees (Sauermann and Roach, 2012). This is in spite of strong encouragement from advisor, who actively discourage other career paths (Sauermann and Roach, 2012).
And the jobs people are actually getting (taken from Polka, 2014) show that, actually, academia is one of the “alternative” careers, and not the default. Less than 10 % of entering PhD students become tenure-track faculty: contrast this to the 50 % identified previously as having academia as their primary goal (Sauermann and Roach, 2012).
Taken from Polka, 2014
What are trainees supposed to accomplish?
This workshop is first going to discuss the question, “In what way are trainees unprepared?” and then depending on what attendees come up with, will then ask, “What are the solutions to these problems?” Then attends will try to arrange these according to feasibility/difficulty of implementation. If there is time, the group will discuss how these fit into the overarching themes of making science efficient, and trying to figure out if these help achieve the greater “goals of science”.
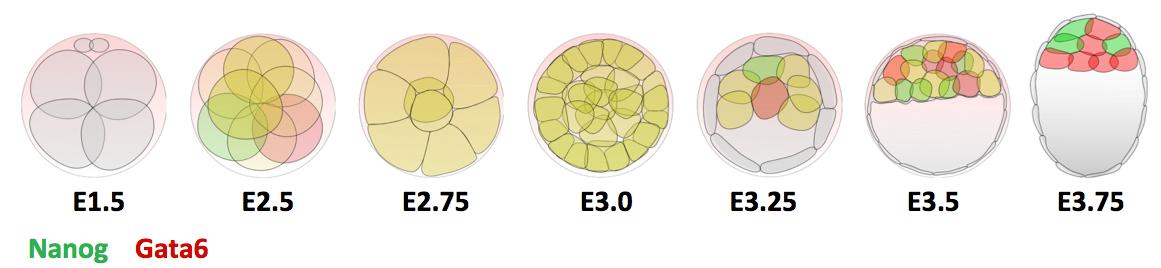
During mouse preimplantation development, the zygote divides and forms three distinct lineages: one embryonic called the Epiblast (Epi) and two extraembryonic called trophectoderm (TE) and Primitive Endoderm (PrE). The first cell fate decision occurs at the morula stage (from 16-cell to 32-cell) between TE and the Inner Cell Mass (ICM) and the second cell fate decision occurs within the ICM that gives rise to the Epi and the PrE in a salt-and-pepper organization during blastocyst formation(Artus and Chazaud, 2014). The complete specification of these three lineages is crucial for correct developmental progression. Moreover, the ICM represents the stem cell niche of Embryonic Stem Cells (ES cells) derived in 1981(Evans and Kaufman, 1981; Martin, 1981). Studying ES cells specification in vivo is important for their better use in regenerative medicine.
Previous research shows that Gata6 and Nanog are key transcription factors for the Epi/PrE cell fate specification. Whereas these two markers are co-expressed in all ICM cells at morula stage, their expression becomes mutually exclusive, asynchronously leading to intermingled Gata6-positive PrE cells and Nanog-positive Epi cells (Figure 1). Moreover, the Fgf/RTK signaling is sufficient and necessary to modulate the salt-and-pepper organisation by controlling Gata6 and Nanog expression (Artus and Chazaud, 2014).
Figure 1 : Expression profile of Gata6 and Nanog during preimplantation development.
We re-examined the role of Gata6 and showed that this transcription factor is required for PrE specification (Figure 2). The PrE defects cannot be rescued by Fgf4 administration, meaning that the cells need Gata6 function to be able to respond to the Fgf signal. These series of experiments also showed that, like in vitro, Nanog expression can be inhibited by the RTK pathway independently of Gata6 function, suggesting a direct repression. Interestingly, by examining Gata6 heterozygotes we also show that the amount of Gata6 is important not only to determine the Epi/PrE ratio but also to regulate the timing of Epi specification and determination. Some of these findings were also reported by Schrode and collaborators (Schrode et al., 2014).
Figure 2: E3.75 immunofluorescence against Cdx2 (blue), Nanog (green) and Gata6 (red).
All these observations suggest that Nanog, Gata6 and the Fgf4/Fgfr pathway act as a regulatory network to specify Epi and PrE cells. Despite progress in understanding the basic principles of stem cell niches, other techniques are required to define the dynamic spatio-temporal induction of salt-and-pepper. One alternative approach is mathematical modeling, based on the translation of biological interactions into mathematical equations. The main feature is to follow the dynamics of factors through time in a defined context.
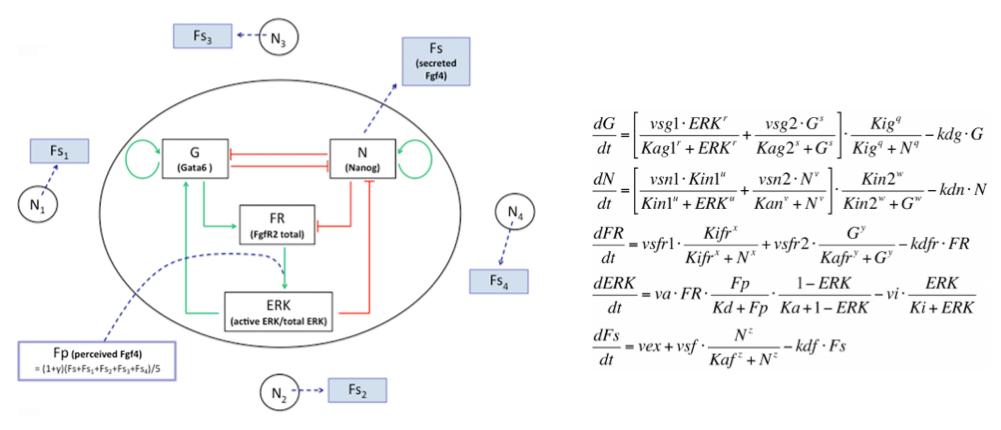
We developed a fruitful collaboration with the Theoretical Chronobiology Laboratory (ULB, Brussels- Belgium). The aim was to develop a mathematical model based on the regulatory network, using published data combined with our results on the analysis of Gata6 mutants (also described in this publication). The model is defined by a set of differential equations. Three of them represent three variables corresponding to the level of intracellular proteins: Gata6, Nanog and Fgfr2. A fourth one represents the activity of Fgfr/Erk signaling, which depends on the extracellular concentration of Fgf4, the fifth variable of the system (Figure 3).
Figure 3: Representation of the regulatory network and its corresponding equations.
The model uncovers three stable steady states (tristability): an ICM-like state where Gata6 and Nanog are co-expressed, a PrE-like state where only Gata6 is expressed and an Epi-like state where only Nanog is expressed.
Simulations show that the model is sufficient to observe Epi and PrE cell specification and resume in vivo findings: 1/ Epi and PrE cells are organized in a salt-and-pepper manner, 2/ this organization is obtained by an asynchronous mechanism, 3/ Epi cells are specified earlier compared to PrE cells and 4/ Epi/PrE ratios obtained fit with in vivo results. The model also recapitulates cell behaviours in the different mutants (Nanog-/-, Gata6-/-, Gata6-/+) (Frankenberg et al, 2011; Bessonnard et al, 2014).
After reaching the ICM state, a subset of cells favours Nanog expression and acquire the Epi-like state. In response to Nanog, cells secrete Fgf4 and modify their local concentration in the neighboring intercellular space. The local increase of Fgf4 is sufficient to push neighboring cells to favour Gata6 expression and reach the PrE-like state. Interestingly, simulations show that only one Nanog-expressing cell could be sufficient to induce the salt and pepper pattern.
The heterogeneity of Fgf4 distribution is crucial to resume the salt-and-pepper organization. It is inherent to the mechanism and causes the asynchrony in cell specification. Thus the model mathematically explains the in vivo observation in which Fgf4 mutants cultured in presence of recombinant Fgf4 cannot restore the salt and pepper pattern and act in a binary fashion (Kang et al., 2013; Krawchuk et al., 2013).
Even if we do not know how one or few cells favour Nanog expression, the following steps of the model are deterministic. Several hypotheses on how Nanog expression is promoted are proposed. The model suggests that it is the consequence of a pErk decrease, following the diminution and dilution of an early Fgf4 expression that was observed by Guo and collaborators (Guo et al., 2010). Alternatively, differences in pErk or Nanog expression could be influenced by cell position (Morris et al., 2010) or stochastically induced (Ohnishi et al., 2014; Yamanaka et al., 2010). More experiments will be required to answer this highly debated question.
In conclusion, this close collaboration enabled us to propose new insights for the salt-and-pepper induction in vivo that will be also useful for stem cell research. More generally, our study is a good example to show that experimental biology and mathematical modeling can synergize to provide insightful results and hypotheses.
Acknowledgments: I thank Dr. Claire Chazaud, Dr. Prudence Donovan and Pr. Albert Goldbeter for reading the article.
Artus, J., & Chazaud, C. (2014). A close look at the mammalian blastocyst: epiblast and primitive endoderm formation Cellular and Molecular Life Sciences, 71 (17), 3327-3338 DOI: 10.1007/s00018-014-1630-3
Bessonnard, S., De Mot, L., Gonze, D., Barriol, M., Dennis, C., Goldbeter, A., Dupont, G., & Chazaud, C. (2014). Gata6, Nanog and Erk signaling control cell fate in the inner cell mass through a tristable regulatory network Development, 141 (19), 3637-3648 DOI: 10.1242/dev.109678
Evans, M., & Kaufman, M. (1981). Establishment in culture of pluripotential cells from mouse embryos Nature, 292 (5819), 154-156 DOI: 10.1038/292154a0
Frankenberg, S., Gerbe, F., Bessonnard, S., Belville, C., Pouchin, P., Bardot, O., & Chazaud, C. (2011). Primitive Endoderm Differentiates via a Three-Step Mechanism Involving Nanog and RTK Signaling Developmental Cell, 21 (6), 1005-1013 DOI: 10.1016/j.devcel.2011.10.019
Guo, G., Huss, M., Tong, G., Wang, C., Li Sun, L., Clarke, N., & Robson, P. (2010). Resolution of Cell Fate Decisions Revealed by Single-Cell Gene Expression Analysis from Zygote to Blastocyst Developmental Cell, 18 (4), 675-685 DOI: 10.1016/j.devcel.2010.02.012
Kang, M., Piliszek, A., Artus, J., & Hadjantonakis, A. (2012). FGF4 is required for lineage restriction and salt-and-pepper distribution of primitive endoderm factors but not their initial expression in the mouse Development, 140 (2), 267-279 DOI: 10.1242/dev.084996
Krawchuk, D., Honma-Yamanaka, N., Anani, S., & Yamanaka, Y. (2013). FGF4 is a limiting factor controlling the proportions of primitive endoderm and epiblast in the ICM of the mouse blastocyst Developmental Biology, 384 (1), 65-71 DOI: 10.1016/j.ydbio.2013.09.023
Martin, G. (1981). Isolation of a pluripotent cell line from early mouse embryos cultured in medium conditioned by teratocarcinoma stem cells. Proceedings of the National Academy of Sciences, 78 (12), 7634-7638 DOI: 10.1073/pnas.78.12.7634
Morris, S., Teo, R., Li, H., Robson, P., Glover, D., & Zernicka-Goetz, M. (2010). Origin and formation of the first two distinct cell types of the inner cell mass in the mouse embryo Proceedings of the National Academy of Sciences, 107 (14), 6364-6369 DOI: 10.1073/pnas.0915063107
Ohnishi, Y., Huber, W., Tsumura, A., Kang, M., Xenopoulos, P., Kurimoto, K., Oleś, A., Araúzo-Bravo, M., Saitou, M., Hadjantonakis, A., & Hiiragi, T. (2013). Cell-to-cell expression variability followed by signal reinforcement progressively segregates early mouse lineages Nature Cell Biology, 16 (1), 27-37 DOI: 10.1038/ncb2881
Schrode N, Saiz N, Di Talia S, & Hadjantonakis AK (2014). GATA6 levels modulate primitive endoderm cell fate choice and timing in the mouse blastocyst. Developmental cell, 29 (4), 454-67 PMID: 24835466
Yamanaka, Y., Lanner, F., & Rossant, J. (2010). FGF signal-dependent segregation of primitive endoderm and epiblast in the mouse blastocyst Development, 137 (5), 715-724 DOI: 10.1242/dev.043471
Here are the highlights from the current issue of Development:
Modelling fate decisions in the early mouse embryo
In the early embryo, the first fate decision separates the trophectoderm from the inner cell mass (ICM). Subsequently, the ICM segregates into epiblast (Epi) and primitive endoderm (PrE), but how do cells decide which of these two fates to adopt? Claire Chazaud, Geneviève Dupont and co-workers address this question in the early mouse embryo (see p. 3637), using both experimental and modelling approaches. It is known that the FGF signalling pathway is involved in this fate decision and also that, while Epi cells express and depend on Nanog, PrE cells are defined by Gata6 expression. The authors complete the interaction network by demonstrating that Gata6 is not only expressed by the PrE but is also required for PrE specification. To understand the network in greater detail, the authors build a mathematical model incorporating the known interactions between these players. The model exhibits tristability, with steady states corresponding to cells displaying ICM, Epi or PrE fate, and is able to recapitulate experimental manipulations of the system, such as the removal of Nanog or Gata6 activity or the effects of altering FGF signalling. Moreover, this approach highlights a number of key features of the system – most notably the requirement for heterogeneity in FGF signalling levels and the self-organising nature of the process of fate determination in this context.
First bird born via ICSI
Intra-cytoplasmic sperm injection (ICSI) is a method of fertilisation that involves the injection of a single sperm into an oocyte. The technique has been applied successfully to generate viable offspring in human, mouse and other vertebrates, but never before has a live bird been produced using ICSI due to difficulties in mimicking avian polyspermic fertilisation. Now, on p.3799, Tomohiro Sasanami and colleagues report the birth of a live quail chick using ICSI and identify some critical parameters that hitherto prevented the successful application of this technique. The authors inject a single sperm together with sperm extract, of which they identify three different components essential for full egg activation and development: phospholipase Cζ, aconitate hydratase and citrate synthase. Importantly, the authors use this system to analyse Ca2+ dynamics during egg activation and show that, in contrast to mammals, avian egg activation requires two distinct patterns of Ca2+ flux: an initial transient rise and a long-lasting spiral-like oscillation. These findings represent an exciting step forward in the study of vertebrate egg activation and will provide new opportunities for the production of transgenic and cloned birds.
Brain keeps a stiff upper lip during development
Physical properties such as tissue stiffness have been shown to influence stem cell fate in vitro. It is possible that spatiotemporal changes in stiffness may influence tissue formation during development; however, it has been difficult to analyse this due to the technical limitations of measuring stiffness in embryonic tissues. In this issue (p. 3793), Yoichi Kosodo and colleagues overcome this problem and provide a novel approach to measuring changes in stiffness using atomic force microscopy combined with immunostaining in the embryonic mouse cerebral cortex during development. By combining these two techniques, the authors show that all layers in the developing cortex undergo significant changes in stiffness throughout the embryonic stages. In the ventricular and subventricular zones, stiffness gradually increases throughout development, while the intermediate zone and cortical plate show an initial increase in stiffness followed by a decrease before birth. Deconstruction of the role of cell and matrix in modulating stiffness in the developing brain reveals that tissue stiffness cannot be solely determined by the stiffness of the cells that constitute the tissue. This study not only sheds light on the spatiotemporal dynamics of stiffness in the developing mouse brain, but also provides a possible approach for understanding the global profile of physical properties in other developing organs.
Mesoderm gets a move on during Xenopus involution
Gastrulation occurs when a single-layered blastula transitions to a gastrula containing all three germ layers: ectoderm, endoderm and mesoderm. Many aspects of vertebrate gastrulation have been well-characterised using the model vertebrate Xenopus, but surprisingly little is known about how the dorsal mesoderm moves from the outer surface to the interior of the gastrula. Now, on p. 3649, Rudolf Winklbauer and colleagues analyse the cellular and molecular dynamics that are central to this involution process and provide several insights into the mechanism of Xenopus gastrulation. First, the authors show that the blastopore lip is capable of autonomous shape changes and that different domains, for example those marked by Goosecoid and Brachyury (Xbra), show differences in cellular migration patterns. Second, the authors investigate the molecular requirements during these events and show that the tyrosine kinase ephrin receptor EphA4 is transiently required to maintain appropriate levels of Xbra expression, but that this occurs indirectly via p21-activated kinase 1 (Pak1). These data highlight a role for Xbra during mesoderm involution in Xenopus and shed light on the general features of dynamic cell migration during gastrulation.
DMRT6 gives spermatogenesis the ‘go-ahead’
The mitotic-to-meiotic transition during spermatogenesis is essential for generating haploid spermatid cells from diploid spermatogonial cells. After differentiation and proliferation of spermatogonial cells, two rounds of meiosis are required to generate spermatid cells, but how the complex transition from mitosis to meiosis is coordinated remains unclear. In this issue (p. 3662), David Zarkower and colleagues investigate the role of theDoublesex-related gene Dmrt6 in male mouse spermatogenesis and show that without DMRT6 this process is severely compromised. By creating a null mutation in two different genetic backgrounds, the authors provide strong evidence that DMRT6 is required for spermatogonial differentiation (C57BL/6J strain) and meiosis (129Sv strain) and that inappropriate gene expression and chromatin events occur when DMRT6 activity is absent. Subsequent mRNA profiling and chromatin precipitation experiments suggest that DMRT6 functions both to suppress genes that promote spermatogonial differentiation and to activate important meiotic regulatory genes. These data provide novel insight into the molecular regulatory events that control and coordinate the mitotic-to-meiotic transition during mammalian spermatogenesis.
PLUS…
Adult neurogenesis: taking stock in Stockholm
In May this year, Stockholm hosted a Keystone Symposium on Adult Neurogenesis, attracting scientists from around the world despite the lack of customary snow. The symposium offered an extraordinary program, covering diverse topics that ranged from the neural stem cell lineage and regulation of neurogenesis to functional aspects of neurogenesis in homeostasis and disease, and even computational modeling. Here, Nambirajan Govindarajan and Gerd Kempermann describe some of the exciting presentations and emerging themes from the symposium, which reveal how much this young field has matured. See the Meeting Review on p. 3615
Poised chromatin in the mammalian germ line
< Poised (bivalent) chromatin is defined by the simultaneous presence of histone modifications associated with both gene activation and repression. This epigenetic feature was first observed at promoters of lineage-specific regulatory genes in embryonic stem cells in culture. More recent work has shown that, in vivo, mammalian germ cells maintain poised chromatin at promoters of many genes that regulate somatic development, and that they retain this state from fetal stages through meiosis and gametogenesis. We hypothesize that the poised chromatin state is essential for germ cell identity and function. Here, Bluma Lesch and David Page propose roles for poised chromatin in the mammalian germ line and discuss these roles in the context of recently proposed models for germline potency and epigenetic inheritance. See the Hypothesis article on p. 3619
Ultradian oscillations and pulses: coordinating cellular responses and cell fate decisions
Biological clocks play key roles in organismal development, homeostasis and function. In recent years, much work has focused on circadian clocks, but emerging studies have highlighted the existence of ultradian oscillators – those with a much shorter periodicity than 24 h. Accumulating evidence, together with recently developed optogenetic approaches, suggests that such ultradian oscillators play important roles during cell fate decisions, and analyzing the functional links between ultradian oscillation and cell fate determination will contribute to a deeper understanding of the design principle of developing embryos. Here, Akihiro Isomura and Ryoichiro Kageyama discuss the mechanisms of ultradian oscillatory dynamics and introduce examples of ultradian oscillators in various biological contexts. See the Review on p. 3627
The conference gives a chance for researchers in various fields, but with the common model system of Xenopus, to discuss their work. The meeting this year took a new direction in not having themed sessions, but rather mixing talks from very different fields into each session. This had the dual purpose of keeping everyone’s attention by switching between different subjects rapidly, but also ensuring people were more likely to stay around for talks outside their field. There was also a strong focus on getting junior faculty and postdocs to speak rather than just the usual senior researchers that are well-known. For these reasons, I personally found this the most engaging conference I have been to.
Keynote talks
There were special lectures given by John Gurdon (Gurdon Institute, University of Cambridge) and Marc Kirschner (Department of Systems Biology, Harvard Medical School), both senior figures in the Xenopus community. John’s talk in particular gave an historical perspective on the beginnings of Xenopus as a model organism, including the transport of frogs from South Africa to the UK by Lancelot Hogben. Apparently, in the early days he was forced to keep his frog colony in the crypt of a church and managed to balance the books by charging hospitals to use frogs as pregnancy tests: when injected with the urine of pregnant women, the hormones present would induce the release of eggs from the female frogs. And from such humble beginnings, the research community has grown, with of course achievements such as John’s Nobel Prize. John speculated on the possible future for research in the frog and the aspiration to be able to track the behavior of individual proteins, within a cell, in real time.
John Gurdon’s special lecture: a historical perspective on the field of Xenopus research.
The keynote talk was given by Rebecca Heald of University of California, Berkeley. Rebecca’s lab focuses on questions to do with size and scaling of cells. In particular, looking at frogs there are basic questions about different species and different stages of development. Xenopus laevis eggs are nearly twice as big as Xenopus tropicalis and yet they are both single cells and, upon fertilization, undergo cleavage in the same way. How does this relate to the DNA content of the cells and how is this remarkable feat of engineering, in an cell so large, achieved? Also, as frog embryos develop, cells continuously divide whilst the embryo as a whole does not grow. What are the implications of maintaining a similar mechanism of cell division, starting in a cell as large as 1 mm in diameter and ending up at a fraction of the size? Work in frog egg and embryo extract is a significant part of the Xenopus field but not often universally used by frog researchers and it was an overview of some of the amazing work that can be achieved with the simple yet versatile extract system.
Rebecca Heald’s tribute to the “Founding Froggers”. Photo courtesy of Cat Vicente.
The community
The frog community also has numerous resources that we simply couldn’t do without: you can check out all things frog at Xenbase and there are three resource centres around the world: the National Xenopus Resource (NXR) in Wood’s Hole, MA; the European Xenopus Resource Centre (EXRC) at the University of Portsmouth, UK; and the National BioResource Project (NBRP) in Hiroshima, Japan which have stocks of wild type and transgenic frogs as well as components such as DNA clones and antibodies. All of these groups presented updates on work they are doing to support the community, such as the genome-editing workshop planned for November 2014 at Wood’s Hole.
Two special workshops were also arranged. One for trainee researchers looking for career advice both in academic and non-academic areas; and one for junior faculty looking for advice on getting tenure. I attended the PhD/postdoc session (as did the Node’s Cat Vicente – as a speaker for non-academic careers) and although the outlook is, as ever, sobering, it was still good to hear some advice specific to people who had worked on frog and some key things to keep in mind. Especially that it’s never too early to be thinking about the next step.
These are both key examples of why I’ve enjoyed my scientific training in the Xenopus community. I started off after finishing my Master’s in Chemistry and moved into a frog lab, and have worked in 3 frog labs/projects since. The sense of community and support provided by the Xenopus field is really quite unique and many who have passed through frog meetings have commented on this. There are some key figures driving the support of more junior faculty and trainees, which is sadly not the norm across all fields of academia.
Looking forward
Work in the model organism Xenopus seems to be really taking off because of all the different techniques that are available to frog researchers. Genetics and genome editing, with TALENs and the CRISP/Cas system, have expanded recently, particularly using the diploid genome of Xenopus tropicalis, but even the pseudoallotetraploid genome of Xenopus laevis, for so long considered a disadvantage, is by its nature of being so unique and outbred generating a lot of questions that other inbred model systems simply can’t answer. The use of Xenopus extracts for in vitro biochemical work was showcased in particular by Rebecca Heald’s lab and the questions that can be answered by the cell-in-a-test-tube system that Xenopus provides still leaves many frog researchers surprised. The rapid development and easy manipulation of frog embryos also allows many simple but elegant in vivo experiments. So, in reality, frogs can be used to answer a whole lot of questions rapidly (and cheaply). I will try to expand upon some of these themes in the near future; needless to say I don’t have time here except to mention what an adaptable model system is across many fields, from bioengineering to biochemistry to biophysics.
Frog friends! Photo courtesy of Atsushi Suzuki, from the National BioResource Project, Japan.
This was my third Xenopus conference; each time I’m unsure if I’ll be at the next one, so I always make sure to enjoy the company of my froggy friends; we always make sure to have a good time!
If you want to keep up with frog-related updates, you can follow various twitter accounts for Xenbase @Xenbase, the National Xenopus Resource at the Marine Biological Laboratory at Wood’s Hole @XenopusNXR and I also blog at bewareofthefrog.tumblr.com and @BiophysicalFrog.
 (1 votes)
(1 votes) (No Ratings Yet)
(No Ratings Yet)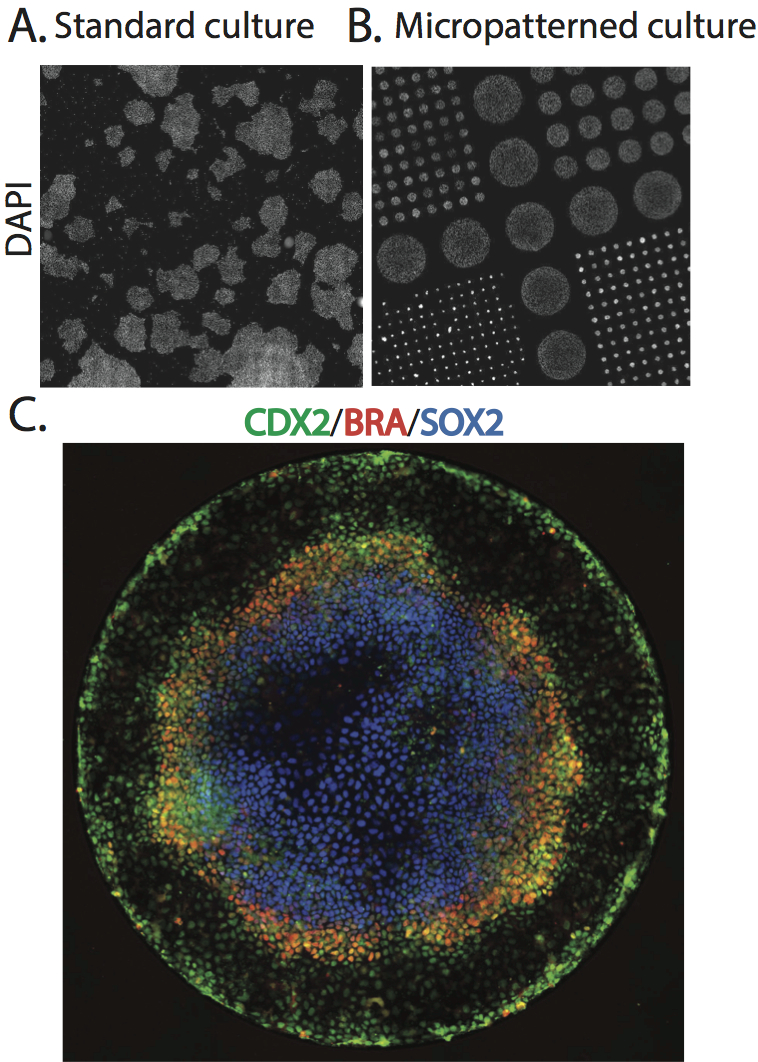
 (7 votes)
(7 votes)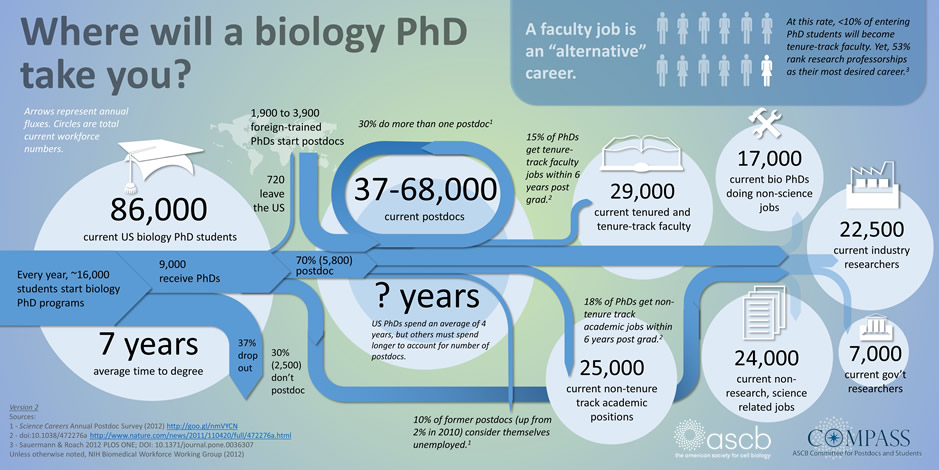

 In the early embryo, the first fate decision separates the trophectoderm from the inner cell mass (ICM). Subsequently, the ICM segregates into epiblast (Epi) and primitive endoderm (PrE), but how do cells decide which of these two fates to adopt? Claire Chazaud, Geneviève Dupont and co-workers address this question in the early mouse embryo (see p.
In the early embryo, the first fate decision separates the trophectoderm from the inner cell mass (ICM). Subsequently, the ICM segregates into epiblast (Epi) and primitive endoderm (PrE), but how do cells decide which of these two fates to adopt? Claire Chazaud, Geneviève Dupont and co-workers address this question in the early mouse embryo (see p.  Intra-cytoplasmic sperm injection (ICSI) is a method of fertilisation that involves the injection of a single sperm into an oocyte. The technique has been applied successfully to generate viable offspring in human, mouse and other vertebrates, but never before has a live bird been produced using ICSI due to difficulties in mimicking avian polyspermic fertilisation. Now, on p.
Intra-cytoplasmic sperm injection (ICSI) is a method of fertilisation that involves the injection of a single sperm into an oocyte. The technique has been applied successfully to generate viable offspring in human, mouse and other vertebrates, but never before has a live bird been produced using ICSI due to difficulties in mimicking avian polyspermic fertilisation. Now, on p. Physical properties such as tissue stiffness have been shown to influence stem cell fate in vitro. It is possible that spatiotemporal changes in stiffness may influence tissue formation during development; however, it has been difficult to analyse this due to the technical limitations of measuring stiffness in embryonic tissues. In this issue (p.
Physical properties such as tissue stiffness have been shown to influence stem cell fate in vitro. It is possible that spatiotemporal changes in stiffness may influence tissue formation during development; however, it has been difficult to analyse this due to the technical limitations of measuring stiffness in embryonic tissues. In this issue (p.  Gastrulation occurs when a single-layered blastula transitions to a gastrula containing all three germ layers: ectoderm, endoderm and mesoderm. Many aspects of vertebrate gastrulation have been well-characterised using the model vertebrate Xenopus, but surprisingly little is known about how the dorsal mesoderm moves from the outer surface to the interior of the gastrula. Now, on p.
Gastrulation occurs when a single-layered blastula transitions to a gastrula containing all three germ layers: ectoderm, endoderm and mesoderm. Many aspects of vertebrate gastrulation have been well-characterised using the model vertebrate Xenopus, but surprisingly little is known about how the dorsal mesoderm moves from the outer surface to the interior of the gastrula. Now, on p.  The mitotic-to-meiotic transition during spermatogenesis is essential for generating haploid spermatid cells from diploid spermatogonial cells. After differentiation and proliferation of spermatogonial cells, two rounds of meiosis are required to generate spermatid cells, but how the complex transition from mitosis to meiosis is coordinated remains unclear. In this issue (p.
The mitotic-to-meiotic transition during spermatogenesis is essential for generating haploid spermatid cells from diploid spermatogonial cells. After differentiation and proliferation of spermatogonial cells, two rounds of meiosis are required to generate spermatid cells, but how the complex transition from mitosis to meiosis is coordinated remains unclear. In this issue (p.  In May this year, Stockholm hosted a Keystone Symposium on Adult Neurogenesis, attracting scientists from around the world despite the lack of customary snow. The symposium offered an extraordinary program, covering diverse topics that ranged from the neural stem cell lineage and regulation of neurogenesis to functional aspects of neurogenesis in homeostasis and disease, and even computational modeling. Here
In May this year, Stockholm hosted a Keystone Symposium on Adult Neurogenesis, attracting scientists from around the world despite the lack of customary snow. The symposium offered an extraordinary program, covering diverse topics that ranged from the neural stem cell lineage and regulation of neurogenesis to functional aspects of neurogenesis in homeostasis and disease, and even computational modeling. Here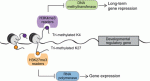 Poised (bivalent) chromatin is defined by the simultaneous presence of histone modifications associated with both gene activation and repression. This epigenetic feature was first observed at promoters of lineage-specific regulatory genes in embryonic stem cells in culture. More recent work has shown that,
Poised (bivalent) chromatin is defined by the simultaneous presence of histone modifications associated with both gene activation and repression. This epigenetic feature was first observed at promoters of lineage-specific regulatory genes in embryonic stem cells in culture. More recent work has shown that, 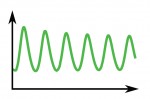 Biological clocks play key roles in organismal development, homeostasis and function. In recent years, much work has focused on circadian clocks, but emerging studies have highlighted the existence of ultradian oscillators – those with a much shorter periodicity than 24 h. Accumulating evidence, together with recently developed optogenetic approaches, suggests that such ultradian oscillators play important roles during cell fate decisions, and analyzing the functional links between ultradian oscillation and cell fate determination will contribute to a deeper understanding of the design principle of developing embryos. Here, Akihiro Isomura and
Biological clocks play key roles in organismal development, homeostasis and function. In recent years, much work has focused on circadian clocks, but emerging studies have highlighted the existence of ultradian oscillators – those with a much shorter periodicity than 24 h. Accumulating evidence, together with recently developed optogenetic approaches, suggests that such ultradian oscillators play important roles during cell fate decisions, and analyzing the functional links between ultradian oscillation and cell fate determination will contribute to a deeper understanding of the design principle of developing embryos. Here, Akihiro Isomura and

