EuroStemCell September 2014 Newsletter: Creative stem cells
Posted by jbarfoot, on 10 October 2014
Super Cells science exhibit launches first leg of 5-year international tour
 There’s the Builder, the Renewer, the Regenerator and a tricky little character known as the Transformer. These four unusual and charming superheroes are the guides in a new science exhibit to be officially launched today at the Sherbrooke Museum of Nature and Science. Their goal? To show museum visitors the power and wonder of stem cells, our body’s master cells. Read more
There’s the Builder, the Renewer, the Regenerator and a tricky little character known as the Transformer. These four unusual and charming superheroes are the guides in a new science exhibit to be officially launched today at the Sherbrooke Museum of Nature and Science. Their goal? To show museum visitors the power and wonder of stem cells, our body’s master cells. Read more
CRG produces stunning art/science exhibition in Barcelona
 The Centre for Genomic Regulation (CRG), Barcelona and partners have produced a stunning and unique art/science exhibition titled “Tree of Life. The complexity of life: from the cell to the living organism“. The exhibition is open to the public at Palau Robert (Passeig de Gràcia, Barcelona) through support by the Banc Sabadell Foundation. It aims to both share the work done at the CRG, and reveal the beauty and uniqueness of the images, showing the parallels and synergies between art and science. Read more
The Centre for Genomic Regulation (CRG), Barcelona and partners have produced a stunning and unique art/science exhibition titled “Tree of Life. The complexity of life: from the cell to the living organism“. The exhibition is open to the public at Palau Robert (Passeig de Gràcia, Barcelona) through support by the Banc Sabadell Foundation. It aims to both share the work done at the CRG, and reveal the beauty and uniqueness of the images, showing the parallels and synergies between art and science. Read more
What’s in a name? Pathways in development
 In this blog, Vanessa De Mello (Hippo pathway enthusiast and PhD student in the Musculoskeletal group, University of Aberdeen) explores the names behind signalling pathways that control stem cells. During growth and development the cells that make up our body need to be precisely controlled. If a ‘stop signal’ is constantly given cells will not grow and divide. But if the lights are continually green, too many cells will grow leading to problems like cancer. It is the ‘developmental signalling pathways’ that control this and give our cells instructions on when to grow, divide, or die. Read more
In this blog, Vanessa De Mello (Hippo pathway enthusiast and PhD student in the Musculoskeletal group, University of Aberdeen) explores the names behind signalling pathways that control stem cells. During growth and development the cells that make up our body need to be precisely controlled. If a ‘stop signal’ is constantly given cells will not grow and divide. But if the lights are continually green, too many cells will grow leading to problems like cancer. It is the ‘developmental signalling pathways’ that control this and give our cells instructions on when to grow, divide, or die. Read more
Researchers grow ‘seed’ of spinal cord tissue in a dish
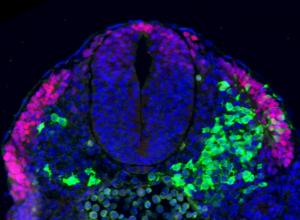 Medical Research Council (MRC) scientists have for the first time managed to turn stem cells into the specialised cells that go on to form spinal cord, muscle and bone tissue in the growing embryo. Their discovery could lead to a new way of studying degenerative conditions such as spinal muscular atrophy, which affects the nerve cells in the spinal column, and may pave the way for future treatments for this and other neuromuscular conditions. Read more
Medical Research Council (MRC) scientists have for the first time managed to turn stem cells into the specialised cells that go on to form spinal cord, muscle and bone tissue in the growing embryo. Their discovery could lead to a new way of studying degenerative conditions such as spinal muscular atrophy, which affects the nerve cells in the spinal column, and may pave the way for future treatments for this and other neuromuscular conditions. Read more
Scientists reset human stem cells to earliest developmental state
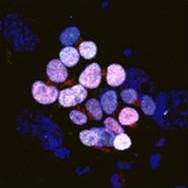 Scientists have successfully ‘reset’ human pluripotent stem cells to the earliest developmental state – equivalent to cells found in an embryo before it implants in the womb (7-9 days old). These ‘pristine’ stem cells may mark the true starting point for human development, but have until now been impossible to replicate in the lab. Read more
Scientists have successfully ‘reset’ human pluripotent stem cells to the earliest developmental state – equivalent to cells found in an embryo before it implants in the womb (7-9 days old). These ‘pristine’ stem cells may mark the true starting point for human development, but have until now been impossible to replicate in the lab. Read more
Film Launch: Cell Fate – Journeys to specialisation. Heidlberg 12th October
 EuroStemCell is delighted to invite members of the public, teachers and schools students are to join us in Heidlberg on the 12th of October (18:00) for the launch of our new dynamic film about stem cells. Produced by award winning science documentary makers, in collaboration with stem cell scientists, this engaging film combines film with animation to create a new perspective on this topic. Read more
EuroStemCell is delighted to invite members of the public, teachers and schools students are to join us in Heidlberg on the 12th of October (18:00) for the launch of our new dynamic film about stem cells. Produced by award winning science documentary makers, in collaboration with stem cell scientists, this engaging film combines film with animation to create a new perspective on this topic. Read more
Continued funding for EuroStemCell
 EuroStemCell is delighted to announce that we have been successful in securing continued funding under the European Commission’s Horizon 2020 funding programme. This means that EuroStemCell, with its strong collaboration of partners, will continue to support the scientific community to engage with citizens, teachers, patients, patient support groups, regulators, policy makers, parlimentarians, journalists…and the many more parties with a strong interest in stem cell research. As well as continuing our current activity, we will be initiating some new and exciting branches of the project with the aim of developing the project and increasing the impact of our activities – watch this space! The EC have produced a news story about the project which can be read here. Read more
EuroStemCell is delighted to announce that we have been successful in securing continued funding under the European Commission’s Horizon 2020 funding programme. This means that EuroStemCell, with its strong collaboration of partners, will continue to support the scientific community to engage with citizens, teachers, patients, patient support groups, regulators, policy makers, parlimentarians, journalists…and the many more parties with a strong interest in stem cell research. As well as continuing our current activity, we will be initiating some new and exciting branches of the project with the aim of developing the project and increasing the impact of our activities – watch this space! The EC have produced a news story about the project which can be read here. Read more
New resources translated into German.
With thanks to the German Stem Cell Network for their support with the translation of these resources and factsheets.
 (No Ratings Yet)
(No Ratings Yet) (1 votes)
(1 votes)


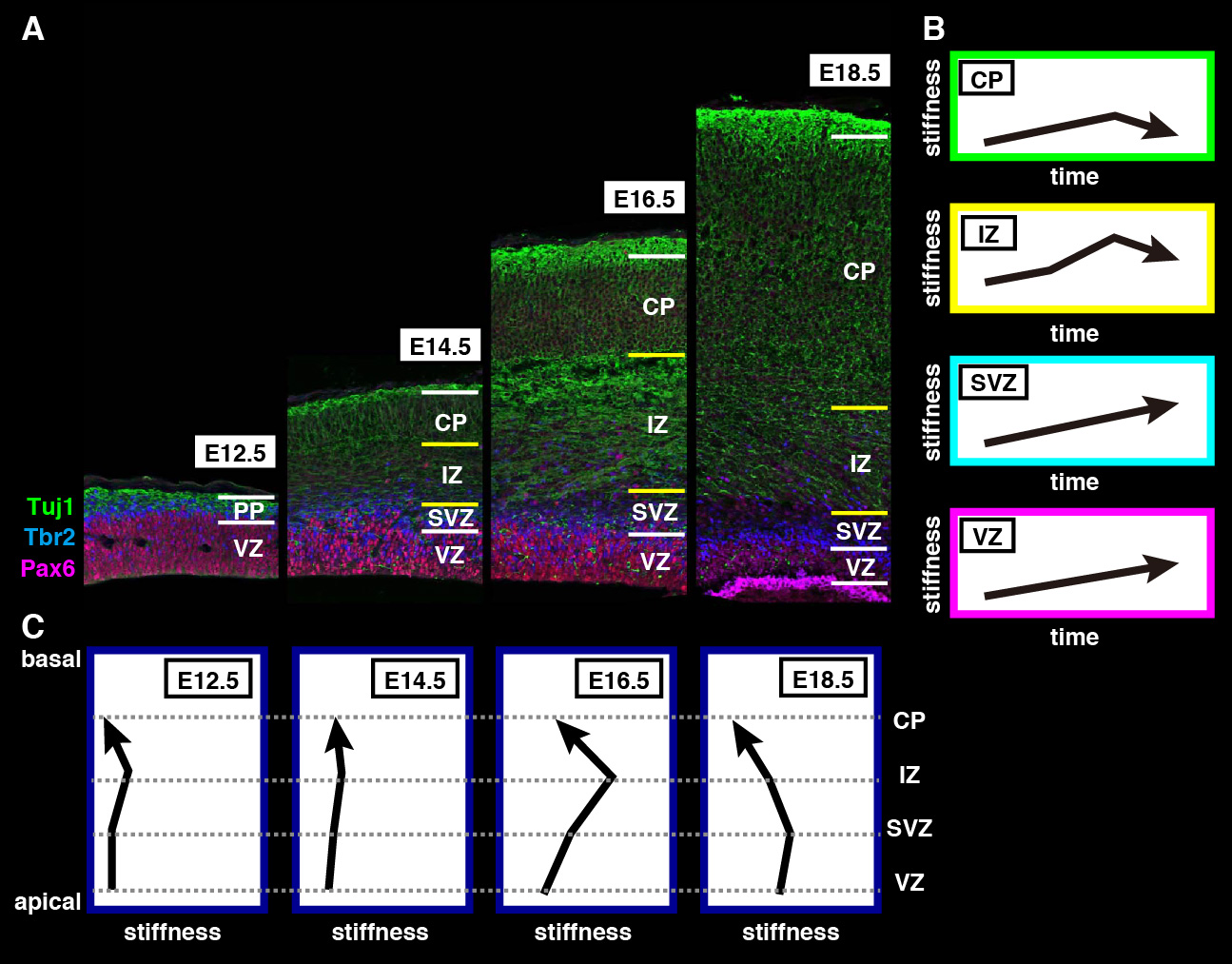
 The repair of cartilage and bone following damage remains a clinical challenge. Current cell-based therapies rely mostly on adult mesenchymal stromal cells, but the expansion of these into correctly differentiated and functionally competent chondrocytes, which give rise to cartilage and then bone, remains problematic. Here, Naoki Nakayama and colleagues develop a small molecule-based approach that mimics the embryonic somitic chondrogenesis programme and can be used to differentiate mouse embryonic stem cells (ESCs) into chondrocytes in vitro (p.
The repair of cartilage and bone following damage remains a clinical challenge. Current cell-based therapies rely mostly on adult mesenchymal stromal cells, but the expansion of these into correctly differentiated and functionally competent chondrocytes, which give rise to cartilage and then bone, remains problematic. Here, Naoki Nakayama and colleagues develop a small molecule-based approach that mimics the embryonic somitic chondrogenesis programme and can be used to differentiate mouse embryonic stem cells (ESCs) into chondrocytes in vitro (p.  The gene orthodenticle homologue 2 (Otx2) encodes a paired-type homeodomain transcription factor that is known to play a role in head morphogenesis. In the mouse, Otx2 is expressed in the anterior neurectoderm, where it is required for the differentiation of anterior neural tissues. Otx2 is also expressed in the anterior mesendoderm (AME) but its role here is unknown. On p.
The gene orthodenticle homologue 2 (Otx2) encodes a paired-type homeodomain transcription factor that is known to play a role in head morphogenesis. In the mouse, Otx2 is expressed in the anterior neurectoderm, where it is required for the differentiation of anterior neural tissues. Otx2 is also expressed in the anterior mesendoderm (AME) but its role here is unknown. On p.  Numerous transcription factors (TFs), including PU.1 and Scl, are known to play important roles during haematopoiesis, but how these act within wider TF networks is unclear. Now, Berthold Göttgens and colleagues use transcription activator-like effectors (TALEs) to manipulate the expression of PU.1 and Scl and determine how these TFs function during developmental haematopoiesis (p.
Numerous transcription factors (TFs), including PU.1 and Scl, are known to play important roles during haematopoiesis, but how these act within wider TF networks is unclear. Now, Berthold Göttgens and colleagues use transcription activator-like effectors (TALEs) to manipulate the expression of PU.1 and Scl and determine how these TFs function during developmental haematopoiesis (p.  Pattern formation during development often depends on the differential regulation of gene expression in response to a morphogen gradient, but how such gradients govern gene expression is unclear. A simplified view suggests that the morphogen activates a transcriptional activator, and that differential gene expression is dependent on the affinity or number of binding sites for this activator within target genes. However, this model does not account for bifunctional transcriptional effectors – those that function as activators and repressors – and has also been questioned by recent experimental results. Here, James Briscoe and colleagues describe a unifying mathematical model of morphogen-dependent gene expression that can explain recent counterintuitive findings (p.
Pattern formation during development often depends on the differential regulation of gene expression in response to a morphogen gradient, but how such gradients govern gene expression is unclear. A simplified view suggests that the morphogen activates a transcriptional activator, and that differential gene expression is dependent on the affinity or number of binding sites for this activator within target genes. However, this model does not account for bifunctional transcriptional effectors – those that function as activators and repressors – and has also been questioned by recent experimental results. Here, James Briscoe and colleagues describe a unifying mathematical model of morphogen-dependent gene expression that can explain recent counterintuitive findings (p.  The T-box family of transcription factors exhibits widespread involvement throughout development in all metazoans. Here, Virginia Papaioannou provides an overview of the key features of T-box transcription factors and highlights their roles and mechanisms of action during various stages of development and in stem/progenitor cell populations. See the Primer on p.
The T-box family of transcription factors exhibits widespread involvement throughout development in all metazoans. Here, Virginia Papaioannou provides an overview of the key features of T-box transcription factors and highlights their roles and mechanisms of action during various stages of development and in stem/progenitor cell populations. See the Primer on p. 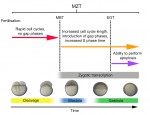 The initial phases of embryonic development occur in the absence of de novo transcription and are instead controlled by maternally inherited mRNAs and proteins. Following this period of transcriptional silence, zygotic transcription begins, the maternal influence on development starts to decrease, and dramatic changes to the cell cycle take place. Here, Steven Harvey and colleagues discuss recent work that is shedding light on the maternal to zygotic transition. See the Review on p.
The initial phases of embryonic development occur in the absence of de novo transcription and are instead controlled by maternally inherited mRNAs and proteins. Following this period of transcriptional silence, zygotic transcription begins, the maternal influence on development starts to decrease, and dramatic changes to the cell cycle take place. Here, Steven Harvey and colleagues discuss recent work that is shedding light on the maternal to zygotic transition. See the Review on p.