Dawei Sun has just finished his PhD in Dr Emma Rawlins’ lab at The Gurdon Institute here in Cambridge, UK. He has remained in the city with his wife for the duration of the pandemic. In the middle of the UK’s second lockdown we met him, virtually, to find out how he is doing, hear about the triumphs and tribulations of his PhD, and discuss his experience of studying in the UK. (more…)
Nuclear-encoded mitochondrial ribosomal proteins (MRPs) are crucial for mitochondrial function and hence cellular energy production. A new paper in Development investigates the function of MRPs in early mouse development, thanks to a battery of knockout mutants. We caught up with first author (and recent graduate) Agnes Cheong and her supervisor Jesse Mager, Associate Professor at the Department of Veterinary and Animal Sciences at the University of Massachusetts, Amherst, to find out more.
Agnes and Jesse at Agnes’ PhD defence – note the mouse cake
Jesse, can you give us your scientific biography and the questions your lab is trying to answer?
JM I did my PhD work with Terry Magnuson at UNC (University of North Carolina at Chapel Hill), studying the Polycomb Group Gene EED and connecting PRC2 with genomic imprinting, X-inactivation and gastrulation defects. I then did postdoctoral work with Rudolf Jaenisch and Marisa Bartolomei, again investigating epigenetic regulation during early development, which I have continued in my own group since 2008 at UMass. We continue to identify novel parent of origin events during gastrulation, as well as characterizing a large number of early lethal phenotypes arising from the Knockout Mouse Project that advance our understanding of the diverse array of genes required for implantation and gastrulation.
Agnes, how did you come to work in Jesse’s lab and what drives your research today?
AC My first research experience involved the study of mammalian implantation and I was really fascinated by the remarkable transformations the embryos undergo during early development. I wanted to continue exploring my interest in embryogenesis, which was the reason why I decided to stay at Jesse’s lab after my rotation. We know so much more than we did decades ago, yet when we take a magnifying glass to zoom into different developmental processes, many blank spaces remain. The desire to comprehend this convoluted process is my motivation to pursue research in development.
When we take a magnifying glass to zoom into different developmental processes, many blank spaces remain
How are each of you coping in the current COVID-19 pandemic?
AC A workday at home is certainly very different from that at the lab. Since I started working from home, I lost the privilege of casually reaching my 10,000 steps daily. Besides my baking hobby, I am otherwise eager to be able to get back to the lab and do some bench work. Fortunately, the Mager lab still has regular virtual lab meetings so that we can present our research ideas and catch up with our lab family, even though we are not able to be physically present together.
JM Since the shutdown mid-March, the lab is now basically just me and the mice…so a bit lonely. But we’ve had lots of Zoom meetings and are trying to keep the lab members busy writing and reading. It definitely took a few weeks to find productive work patterns (and bonus regular lunches with my kids!). Daily evening strolls with my family have been a silver lining – we are fortunate to be able to get outside quite a bit.
Let’s get to your current paper then, which uses mutants produced by the Knockout Mouse Project. What were the aims of that project and your lab’s involvement with it?
AC & JM IMPC/KOMP is a global initiative, and its main goal is to generate a comprehensive and public resource comprising mouse embryonic stem cells and mice containing a null mutation in every gene within the mouse genome. Although the bulk of the effort has focused on adult phenotypes, ∼25% of nulls are lethal and 12% die prior to E9.5. Our involvement with KOMP is to characterize a large number of these early lethal phenotypes since there is no planned pipeline to analyse early embryos. Thus far, we have phenotyped more than 110 gene knockouts, and will grow to ∼250 over the next few years. Check out all our results at http://blogs.umass.edu/jmager/.
Can you give us the key results of the paper in a paragraph?
AC & JM Our paper demonstrates the importance of a functional oxidative phosphorylation (OXPHOS) system during early development. Although there are over 70 mitochondrial ribosomal protein (Mrp) genes in mammals, the absence of a single Mrp gene results in pre-gastrulation developmental arrest. Despite the presence of pluripotent epiblast cells in the Mrp mutants, these embryos fail to initiate EMT and progress through gastrulation. This disruption is the result of a malfunctioning mitochondrial translation system which contributes to a lack of ATP production, and hence results in cell cycle disruption and stalling at the G2/M checkpoint. Our results are aligned with other developmental studies of mitochondrial functioning genes showing that the OXPHOS is indispensable during early embryogenesis.
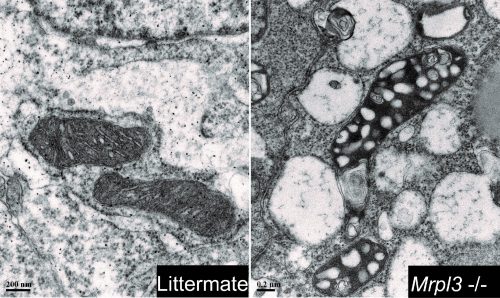
TEM images revealing the different mitochondrial morphologies between mutants and control embryos.
Before developmental arrest, early embryonic development runs normally in Mrp mutants – does this mean the MRPs are doing nothing before then?
AC & JM Not necessarily: since the majority of Mrp genes are also present in the oocyte, maternal Mrp protein could be supporting development through implantation and/or an alternative energy system, such as glycolysis, may be a major factor driving developmental progression prior to gastrulation. A more-detailed study of protein turnover and maternal deletion experiments will provide insights on the precise time point at which Mrp proteins are essential during embryogenesis.
When doing the research, did you have any particular result or eureka moment that has stuck with you?
AC My memorable eureka moment was when I analysed phospho-cdc25c foci and found a significant increase in the mutants, confirming my suspicion that the malfunctioning OXPHOS system leads to cell cycle disruption.
And what about the flipside: any moments of frustration or despair?
The endless sea of genotyping when working with 20+ different knockout lines and needing to genotype every mouse and embryo.
So what next for you after this paper?
AC I recently completed my PhD defence. My plan is to diversify my scientific experience and continue my research career in a biotech or industrial setting.
Where will this work take the Mager lab?
JM Our findings that so many nuclear-encoded mitochondrial functioning genes have such similar phenotypes (gastrulation arrest) are prompting the design of experiments to try to assess the metabolic needs of distinct pre-gastrulation lineages and cell populations. This is a new and exciting area or inquiry for our group. We will also pursue conditional knockout approaches to examine the maternal protein contribution.
Finally, let’s move outside the lab – what do you like to do in your spare time in Amherst?
AC Amherst is a small college town surrounded by many local breweries. When the weather is nice and warm, I enjoy walking to town and reward myself with a hoppy and cold beer. On cold rainy days, I like warming up my apartment with baked goods and sharing them with my friends.
JM I’ll be honest – I’m an exercise enthusiast (to put it mildly)…so if I’m not at work or with my family I’ll generally be found getting those endorphins.
An NSF-supported postdoctoral position is available in the Yakoby Lab https://yakoby.rutgers.edu/ for a highly-motivated and productive individual with an interest in studying the evolution of tissue patterning and morphogenesis in Drosophila oogenesis. The project includes the prediction and validation of cis-Regulatory Modules (CRMs) in the follicular epithelium of several Drosophila species. Applicants with a Ph.D. and a strong background in molecular biology, including the use of CRISPR/Cas9, genetics, and bioinformatics are encouraged to apply. Interested individuals should send a CV, a short statement of research interests, and three names of references to <yakoby@camden.rutgers.edu>. The Yakoby Lab https://yakoby.camden.rutgers.edu/ is in the Biology Department https://biology.camden.rutgers.edu/ and the Center for Computational and Integrative Biology https://ccib.camden.rutgers.edu/ at Rutgers University Camden campus located in the Greater Philadelphia Area.
We are seeking candidates to apply for two upcoming calls for Postdoctoral Researchers from the Spanish Government (Juan de la Cierva).
Candidates should have an interest in Developmental Neurobiology, with expertise ranging from embryology, to stem cell biology, bioinformatics and/or quantitative biology to investigate development of the vertebrate Central Nervous System (CNS) in health and disease. Candidates must have a competitive publication records, since both are competitive open calls.
Candidate will join the Elisa Martí Laboratory (IBMB-CSIC, Barcelona). Please contact Elisa directly emgbmc@ibmb.csic.es the sooner the better (calls are about to open)
Relevant recent publications from the lab
Murielle Saade and cols Cell Stem Cell 2020 Oct 27; doi: 10.1016/j.stem.2020.10.002.
Gwenvael Le Dréau, and cols Elife 2018 Aug 10;7. pii: e37267. doi: 10.7554/eLife.37267
This Obituary by James M. A. Turner, Shantha K. Mahadevaiah, Arthur P. Arnold and Robin Lovell-Badge was recently published in Development.
Paul Burgoyne was an outstanding mouse geneticist and developmental biologist who made fundamental discoveries about the sex chromosomes and their biology. He was born on 10th January 1946, spending most of his childhood living in Dartmoor on a farm with his parents, who were Quakers. The farm was a busy and culturally diverse environment, co-run by a second family (the Haines) and providing residence to foreign students who visited through the International Scholastic Correspondence, an organisation that brought together youngsters from different countries to foster better understanding in the post-war period. Paul considered several academic and non-academic career options, including a farmer, a vet and, at one point, a pilot; the last option he eventually rejected due to a tendency for airsickness. Encouraged by his parents and teachers, he continued to higher education, studying botany and zoology at Portsmouth University. His meticulous and precise approach to science was noted by his college tutor, who persuaded him to take up a technician position at Edinburgh University with Alan Beatty, reproductive biologist and PhD supervisor to the Nobel Prize winner and ‘test-tube baby’ pioneer Robert Edwards. Paul remained in the Beatty lab for his PhD, working on sperm morphology. He loved playing cards, and regularly attended the lunchtime whist club. It was there that he met his beloved wife Kathy, a computer scientist who later helped him analyse his cytological data.
In 1973, Paul moved with Kathy to the USA, where he did a postdoc with John Biggers at Harvard University. During this time, Paul’s interest in sex chromosomes was first piqued, when he discovered that female embryos lacking one X chromosome exhibit impaired development in vitro. Among the highlights of this 3 year stay in the USA was a summer placement at the Jackson labs, where he worked with Wes Whitten, discoverer of the Whitten effect (that male pheromones can stimulate simultaneous oestrus in females), and enjoyed afternoons swimming in the lake. Paul subsequently returned to Edinburgh University to work with Ted Baker, during which period he demonstrated that two X chromosomes are necessary for the development of eggs. He was subsequently invited by eminent developmental biologist Dame Anne McLaren in 1979 to the newly formed MRC Mammalian Development Unit in London. Anne was a huge influence on Paul and was a personal and scientific role model for him. But Anne was equally devoted to Paul; she was delighted when he came to her unit and with the research he did there, and was equally excited about his accomplishments after this.
During his period at the MRC Mammalian Development Unit, and later when he moved (in 1992) to the MRC National Institute for Medical Research in London, Paul made discoveries in sex chromosome biology that have become memorialised in textbooks. He dominated the field of mouse Y chromosome biology, using a diverse array of mouse mutants carrying what he referred to as ‘weird sex chromosomes’ to dissect the function of Y genes in fertility. He was fascinated by meiosis, the point in germ cell development when homologous chromosomes pair and form crossovers. He correctly predicted that the X and Y chromosomes, thought initially to be genetically dissimilar, shared a region of homology through which they exchange genetic information. He named this the ‘pseudoautosomal region’ (PAR), and demonstrated that defective genetic exchange in the PAR was the principle cause of infertility in male mice with abnormal sex chromosomes. His group demonstrated that DNA double-strand breaks, the substrates of genetic exchange, are formed much earlier in meiosis than previously believed, and discovered a novel epigenetic phenomenon, meiotic silencing, in which chromosomes failing to pair are transcriptionally silenced. These studies, marvelled at for their beautiful cytology, revealed to us the intricate surveillance mechanisms that operate in the germ line to ensure that offspring inherit the correct number of chromosomes. Throughout his life, Paul was deeply concerned about overpopulation, and the irony that he worked on infertility was not lost on himself or his friends!
Paul’s love of sex chromosomes also led him beyond reproductive biology, and in a series of papers, most of them published in his favourite journal Development, he made landmark discoveries in sex determination, embryology and sex differences. He found that in chimeric XX-XY mice, most testis cell types could be either XX or XY, but Sertoli cells were exclusively XY. This elegant experiment revealed that the testis-determining factor, not yet identified by the Lovell-Badge and Goodfellow labs as Sry, acted cell-autonomously in Sertoli cell precursors. This became an important criterion to reject or pursue candidates for the gene. He showed that sex chromosomes control the rate of embryonic growth, with a Y chromosome speeding up and a paternal X chromosome slowing down development of the preimplantation mouse embryo. With Art Arnold and Robin Lovell-Badge, he designed and validated a method called the ‘four-core genotypes cross’ that allowed the effects of sex chromosome make-up and gonadal hormones to be uncoupled. Before this point, the impact of sex chromosomes on male-female differences was underappreciated, and it wasn’t until 2014 that the National Institutes of Health mandated the use of both male and females in research studies on animals. The four-core genotypes cross is now a mainstay of research across the globe, revealing how sex chromosomes impact diverse diseases, including autoimmunity, metabolic syndrome, Alzheimer’s and hypertension.
Paul maintained a small research group and was hands-on with respect to science, especially mouse work. He expected to find his trainees in the mouse rooms, peering into cages so that they wouldn’t miss phenotypes. He made twice-weekly visits to the animal house, where he spent the entire afternoon meticulously assessing each of the 70 or so sex-chromosome variant mouse lines. His desire to work on a farm, in close contact with animals, may have motivated this close connection with his mice. Even during a 10-year period when he had become very allergic to mice, he would still trot off to the animal room, looking like an astronaut, with a full face mask and HEPA-filtered air supply. It was during his mouse-room visits that his enthusiasm and genetics prowess were most evident. He would recite complex genetic crosses effortlessly, regaling those shadowing him with witty anecdotes from his life as a trainee. His karyotype preparations were always more beautiful than everyone else’s, and he took great relaxation from sitting in his office, counting chromosomes on his Olympus microscope, a cup of tea by his side and classical music on the radio. He was utterly captivating during scientific presentations, leaving even the most learned geneticists in awe.
Paul was a guileless and modest man who never realised how brilliant or deified he was. He preferred instead to focus on the success of his junior scientists, many of whom now run research groups. He was elected to the Academy of Medical Sciences, but otherwise eschewed formal recognition. To him, research was simply another of his many hobbies, like gardening, hiking, DIY and badminton, all of which he was enviably good at.
Paul is survived by his wife Kathy, daughter Jacqueline, and sons Robert and Thomas.
Acknowledgements
We are grateful to Kathy Burgoyne and Paul’s scientific colleagues for sharing information on which this obituary is based.
The Nerurkar Lab is looking for Postdoctoral Researchers with an interest in the interplay between molecular and mechanical aspects of vertebrate morphogenesis. Using the chick embryo, we
combine live in vivo imaging, embryology and molecular genetics with engineering and physics approaches to study how developmental signals modulate physical forces that shape the embryo,
and how forces in turn feedback on tissue growth and stem cell differentiation. Projects include early morphogenesis and patterning of the gut tube and brain, and organogenesis of the small intestine. Applicants must hold a PhD in molecular biology, development, bioengineering, or related field. Biologists with a strong developmental biology and embryology backgrounds and an interest in building expertise in biophysical and quantitative approaches are particularly encouraged to apply. Part of the Department of Biomedical Engineering at Columbia University, the Nerurkar Lab is located on the Morningside Heights campus of Columbia University in the City of New York. An academic reflection of New York’s excitement and creativity, Columbia offers a rich research environment, with boundless opportunities for collaboration with experts across engineering, biology, and clinical/translational disciplines. Interested applicants should contact Nandan Nerurkar at nln2113@columbia.edu.
The National Institute of Standards and Technology (N.I.S.T.) is recruiting recent PhD’s. The laboratory is located near Washington, DC. Our research program focuses on characterizing the dynamics of gene regulatory networks in human iPSCs and in developing quantitative models that describe the pluripotency landscape. Possible start dates: 6/2021 – 12/2021
We invite interested candidates to apply for Postdoctoral Research Fellowships available through the National Research Council (NRC). These research associateships are two-year appointments with an annual salary of $71,128 (U.S. citizenship is a requirement). These positions are highly competitive and attract outstanding candidates. Research challenges include:
applying state-of-the-art live cell imaging and image analysis and deep learning models to follow gene expression dynamics in single cells
gene editing (i.e. CRISPR-Cas9) to engineer iPSC lines with multiple fluorescent reporters
statistical analysis of time series data to infer gene network interactions
modeling pluripotency regulation
Experience with cell culture, quantitative imaging and image analysis, and/or mathematical modeling is a plus. These postdoc appointments are excellent opportunities for recent Ph.D. graduates who are seeking additional experience prior to accepting a permanent academic or industrial position. Historically, a number of these awardees have also become permanent employees at NIST. Next application deadline is February 1.
Begin the application process by sending your CV and cover letter to Michael Halter (michael.halter@nist.gov).
Job requirements:
-US citizenship
-Within 5 years of obtaining PhD in bioengineering, cell biology, or physical sciences
-PhD is not required at time of application, but is required before beginning Fellowship
-Ability to work in a multidisciplinary team with biologists, bioengineers, computer scientists, physicists, and statisticians
The Marín-Juez laboratory, at the CHU Sainte-Justine Research Center,is recruiting a PhD student and a postdoctoral fellow (4-year fully funded positions). Our laboratory is interested in the cellular and molecular mechanisms regulating cardiac regeneration. The successful applicant will join the Marín-Juez laboratory at the CHU Sainte-Justine Research Center, where he/she will have access to state-of-the-art facilities and technology platforms including Advance imaging platform (light-sheet, spinning-disc confocal, multiphoton, STED super-resolution, etc.), genomics (DropSeq, 10x, Illumina Novaseq) and bioinformatics platforms. CHU Sainte-Justine Research Center provides a thriving scientific environment where the successful applicant will have the opportunity to work with multidisciplinary scientific teams and to collaborate with talented clinicians and researchers.
Research project description
For this project, we are particularly interested in understanding how the cardiac endothelium regulates different aspects of cardiac regeneration and how alterations in the coronary network formation impact the ability of coronary vessels to support tissue replenishment. We have recently found early coronary regeneration as a key determinant of heart regeneration (Marín-Juez et al., PNAS 2016), and identified mechanisms regulating coronary network replenishment to form a vascular scaffold that supports cardiomyocyte regeneration (Marín-Juez et al., Dev Cell 2019). We now seek to define how the different components of the cardiac endothelium regulate tissue replenishment and identify the different mechanisms involved in their regulation of CM proliferation and migration.
Required training and profile
Ph.D. student position: Applicants should have training in vascular biology, molecular biology, cell biology, or related fields. Suitable candidates should be enthusiastic about regenerative and vascular biology. Previous research experience with zebrafish and/or heart regeneration is desired.
Postdoctoral position: We are looking for candidates with a Ph.D. in the biological sciences and laboratory experience in tissue repair/regeneration, cellular, molecular biology, or genetics. Previous experience working with zebrafish, imaging and histology are highly valued but not essential.
Both positions: Candidates with experience in confocal/light-sheet imaging and/or genome engineering are strongly encouraged to apply. Preference will be given to applicants with excellent collaborative and communication skills. The Marín-Juez lab and the CHU Sainte-Justine Research Center subscribe to the principle of equal access to opportunities and encourage women, members of visible and ethnic minorities, persons with disabilities and Indigenous people to apply.
Submit your application
Candidates must send the required documents before 02/2021 to Rubén Marín Juez at ruben.marin.juez.hsj@ssss.gouv.qc.ca
Please provide: Curriculum vitæ, Cover letter and References (2 or 3).
PhD or postdoc project at the Zaidel-Bar lab: A systematic analysis of C. elegans gastrulation in order to uncover molecular mechanisms of cell ingression.
We are currently seeking an independent and motivated research associate to work on a research project led by Dr. Anestis Tsakiridis (https://www.tsakiridislab.com/) and Prof. Peter Andrews at the Centre for Stem Cell Biology, University of Sheffield, UK. The successful applicant will work on the characterisation of candidate human pluripotent stem cell (hPSC)-derived enteric nervous system (ENS) progenitor
populations for use in cell therapy approaches to treat Hirschsprung disease (HSCR), one of the most common congenital diseases affecting the ENS. This project is funded by the MRC and is a collaboration with the group of Conor McCann/Nikhil Thapar at the UCL Great Ormond Street Institute of Child Health in London.
You should have a PhD (or be close to completion/have equivalent experience) in Stem Cell/Developmental Biology and have experience in hPSC culture and neural differentiation as well as in one or more of the following: immunocytochemistry, qPCR, flow cytometry, and Crispr/Cas9 based genetic modification approaches. You must be self-motivated, well-organised, highly independent, and willing to work with other members of the existing research team. Excellent communication, written and interpersonal skills are essential.
We are keen to attract a diverse applicant pool, and we are aware that some under-represented groups are typically less confident about applying for jobs unless they are sure they comfortably meet all the criteria. With this in mind, we encourage
all potential candidates to reflect on their strengths and experience in the broadest sense, including transferable skills where appropriate, when considering their suitability for the position.
We build teams of people from different heritages and lifestyles from across the world, whose talent and contributions complement each other to greatest effect. We believe diversity in all its forms delivers greater impact through research,
teaching and student experience.
We’re one of the best not-for-profit organisations to work for in the UK. The University’s Total Reward Package includes a competitive salary, a generous Pension Scheme and annual leave entitlement, as well as access to a range of learning and
development courses to support your personal and professional development
Closing date: 9th December 2020. Please apply here (Job reference: UOS026765). For informal enquiries email: a.tsakiridis@sheffield.ac.uk
 (3 votes)
(3 votes)
 (No Ratings Yet)
(No Ratings Yet)

