The University of Cambridge is exceptional in the depth and diversity of its research in Stem Cell Biology and has a dynamic and interactive research community that is ranked amongst the foremost in the world. Applications are now being accepted for places on the Wellcome Four-Year PhD Programme in Stem Cell Biology and Medicine, to commence in October 2021. Applicants are welcomed from any country, culture, socio-economic background or university and should demonstrate both a passion for scientific research and a desire to learn about Stem Cell Biology. By bringing together members of both the Schools of Biology and Medicine, this four-year PhD programme enables students on the programme to take advantage of the strength and breadth of stem cell research available in Cambridge, plus the opportunity to complete their research in one of the 25 host laboratories in Cambridge.
Course Outline
The first year MRes degree gives the students an opportunity to complete three different rotation projects in research laboratories, under the supervision of a dedicated Principal Investigator. In addition, students are required to attend weekly Stem Cell Discussion Courses, participate in both presentation sessions and an annual symposium and complete a Personal Researcher Development Training programme. New postgraduate students at the CSCI will become members of the Postgraduate School of the Life Sciences, which organise and promote many further training opportunities. Towards the end of the third rotation project, students are expected to choose a laboratory for their PhD thesis research. Students will prepare a PhD Research Proposal which will form part of the assessment towards the MRes in Stem Cell Biology. Successful completion of the MRes will then lead onto the fully-funded three-year PhD, in a host laboratory.
These fully-funded studentships commence in October 2021. The funding includes the University Composition Fee (at Home (UK) rate), a travel and consumables budget and a stipend at the Wellcome rate (see here). International students can apply for a Cambridge Trust Scholarship during the application process to cover the difference between the ‘Home’ and ‘Overseas’ Fee rate. Other funding opportunities are also available. Further information can be found here: https://www.postgraduate.study.cam.ac.uk/funding/university-funds
The closing date for receipt of applications is: 7 January 2021. Short-listed applicants will be invited to interview the week commencing 25 January 2021. You will be interviewed by a panel from the Cambridge Stem Cell Institute.
Here you’ll find the three talks and their live Q&A sessions, moderated by John Wallingford, one of the guest editors of the issue. The talks will be taken down on Thursday 26 November.
Department/Location: Sainsbury Laboratory, University of Cambridge UK
Salary: £41,526-£52,559
Reference: PT24318
Closing date: 1st December 2020
Applications are invited for a David Sainsbury Career Development Fellowship in the Sainsbury Laboratory, Cambridge University. The Fellowships provide an opportunity for talented junior investigators to develop their own research programme and become creative leaders in the field of quantitative plant developmental biology. We are searching for a candidate who is developing an understanding of design principles that can be used to construct predictive models for complex behaviours in living systems. Applicants are likely to be using theoretical and/or quantitative experimental approaches. Examples include transcriptional modelling, computational morphodynamics, single molecule/cell imaging, systems or synthetic biology and biophysics. The position fits within the overarching goal of the Sainsbury Laboratory to develop a quantitative, dynamic and predictive understanding of plant development, but no previous experience of plant science is necessary. Rather, it is critical that the appointee is willing to engage in the Sainsbury Laboratory’s collaborative research environment.
The Sainsbury Laboratory, Cambridge University is a research institute located in central Cambridge, providing an outstanding scientific environment. It has excellent resources and core facilities, with state-of-the-art equipment including growth rooms, glasshouses, microscopy and computing facilities. It provides a collaborative research atmosphere where researchers can take risks in a supportive environment. We are seeking to appoint David Sainsbury Career Development Fellows who can both contribute to and benefit from this environment. In particular we are looking for scientists with imaginative research ideas that will contribute to our quantitative understanding of plant development by including theoretical and computational approaches.
Applicants from all nationalities are welcome to apply and will typically have postdoctoral experience in a field related to the overall scientific focus of the Laboratory. No prior experience with plants is required – researchers wishing to bring their current wet lab or modelling experience to open novel questions in quantitative plant developmental biology are welcome to apply. Applicants should be able to provide strong evidence of their potential to develop an independent research programme.
The maximum duration of the Fellowship is five years. Successful applicants will initially receive three years of funding, with an extension for a further two years contingent on a successful review and will hold an appointment with the University of Cambridge. Funds include full salary, ranging from £41,526 to £52,559, an additional 30% Gatsby distinction award, benefits and £50,000 p.a. of unrestricted research support. As they develop into independent researchers, Fellows will be encouraged to apply for external funding.
The Laboratory provides a welcoming and collaborative environment with a wide range of family-friendly benefits and development opportunities. More about the Sainsbury Laboratory and details of what the University offers to employees, can be found at https://www.slcu.cam.ac.uk/
Fixed-term: The funds for this post are available for up to 5 years.
Please ensure that you also upload your application as a single pdf file including a full CV, a research proposal for five years, and the names of at least three referees.
Further details about the post and guidance and on how to apply are available below.
The second webinar in the Development presents… series coincides with the November release of our Special Issue: The Origins and Mechanisms of Developmental Disorders. The Special Issue showcases the role of developmental biologists in addressing developmental disorders – the leading biological cause of death for children of all ages – and the talks in our November webinar come from researchers at the forefront of this field. The webinar will be chaired by John Wallingford, Professor of Molecular Biosciences at the University of Texas at Austin, who is one of the guest editors of the Special Issue together with Sally Dunwoodie, Professor of Medicine at the Victor Chang Cardiac Research Institute.
The webinar will be held in Remo, our browser-based conferencing platform – after the talks you’ll have the chance to meet the speakers and other participants at virtual conference tables. If you can’t make it on the day, talks will be available to watch for a couple of weeks after the event (look out for details on the Node).
For more information about what to expect in Remo, go to
Location: Chinese Academy of Sciences, the Guangzhou Institutes of Biomedicine and Health, 190 Kaiyuan Avenue, Guangzhou Science Park, Guangzhou, China
Salary: Competitive with benefits ($45k +), subject to skills and experience
We are seeking highly motivated and collaborative postdocs in studying stem cell lineage of early mammalian embryos.
The purpose of this position is to characterize temporal and spatial mapping of mammalian early embryo and translational applicant of functional stem cells/Organoids. We have developed methods to systematically survey the spatial organization of tissue cell types in single-cell level and identified the temporal and spatial regulation for cell fate commitment in early mouse embryos (Nature, 2019; Nature Genetics, 2019; Dev Cell, 2016). We aim to further understand the mechanisms of stem cell specification and lineage segregation of early mammalian embryos through dynamic, in situ and quantitative approaches.
We seek candidates who are energetic, focused, and productive with a desire to work and lead within a wet and dry mingled lab. Good organizational, analytical, and communication skills are essential. A track-record of original research in biomedical science is required. Experiences in single-cell and spatial omics, bioinformatics are highly desirable.
Project scope
Gunagdun Peng’s laboratory focuses on understanding the mechanism of lineage specification in mammalian early embryo and developing cutting-edge technology of multi-omics, and is keen on stem-cell-based treatment for human diseases.
Research techniques used in the laboratory include: molecular biology, advanced imaging, 2D/3D culture, genome modification, genome-wide lineage tracing, single-cell sequencing, spatial multi-omics, bioinformatics.
About us
Guangzhou Institutes of Biomedicine and Health, Chinese Academy of Sciences (GIBH, CAS) aims to achieve the goal of “Healthy China” and tackle human health challenges.
GIBH has successfully hosted “Guangzhou International Stem Cell and Regenerative Medicine Forum” for ten consecutive years, which invited many world-famous stem cell researchers including Nobel Laureates. GIBH is featured in its influential stem-cell researches.
Interested candidate should apply via email: peng_guangdun@@gibh.ac.cn.
A crucial step in vertebrate muscle development is the activation of myogenic regulatory factors (MRFs) that direct myogenesis. A new paper in Development investigates the roles of Fgf signalling and Tbx transcription factors in zebrafish MRF induction. We caught up with the paper’s two first authors, Daniel Osborn and Kuoyu Li, and their supervisor Simon Hughes, MRC Scientist and Professor of Developmental Cell Biology at King’s College London, to hear more about the story.
Daniel, Kuoyu and Simon (L to R)
Simon, can you give us your scientific biography and the questions your lab is trying to answer?
SH After studying biochemistry as an undergrad and doing a PhD on rhodopsins with Martin Brand in Cambridge, I did a postdoc with Martin Raff in UCL on cell lineage in the optic nerve. In 1987 it was difficult to study cell lineage in vivo in the central nervous system, and the molecules that controlled cell fate were completely mysterious. But MyoD was discovered that year, so I switched to skeletal muscle, doing a second postdoc with Helen Blau at Stanford on how myoblasts generate slow and fast muscle fibres in specific patterns in each muscle. All three advisors taught me so much about both science and life. Before I moved back to London, I worked for a few months in David Botstein’s large group. We were sequencing the yeast genome, a precursor to the Human Genome Project and one of the first high-throughput ‘big science’ projects in biology; really exciting, but not my kind of science. In 1992, I joined the Medical Research Council Biophysics Unit at King’s College London, where Franklin and Wilkins had done their DNA structure work. It was an ideal place for me – Nigel Holder and Roger Patient had just set up a Developmental Biology Research Centre, and my lab linked that to the MRC Unit. We initially worked mainly on mouse and chick muscle, but Nigel soon inveigled me into zebrafish and I fell in love with the simplicity and ability to watch tissue development in real time. I was really lucky to meet, learn from, and collaborate with Monte Westerfield and Phil Ingham and their colleagues, whose differing approaches helped me combine genetics with our developmental cell biology. We have remained true to our original question of understanding the molecular genetics of muscle tissue patterning, though much of our work now focuses on later developmental stages, when zebrafish muscle is growing from committed muscle stem cells. I am really pleased with Dan and Kuoyu’s paper because, although it had a 20 year gestation, I think it fills in a missing link between patterning of early mesoderm and muscle.
Daniel, how did you come to work in Simon’s lab?
DO I started in Simon’s lab as a research technician back in 2001 where my primary role was running the then MRC/KCL zebrafish facility as well as contributing to the group’s research. It was a fantastically encouraging environment for a young biology graduate and I was immediately immersed in exciting developmental biology, the bright lights of London and my first salaried position. I loved the lab work and to fuel my interest Simon offered me a part-time PhD and KCL agreed to waive my bench fees. This sent me down the academic research career path and I haven’t looked back since. My thesis looked at the regulation of myogenic bHLH proteins during zebrafish slow muscle development. A substantial amount of work came out of it, contributing to four papers, and it is now with great pleasure that this collaborative effort has produced a fifth, and perhaps final, paper stemming from my thesis, which was completed 12 years ago.
How are each of you coping in the current COVID-19 pandemic?
DO I am now based at St George’s University of London where I run my own zebrafish group (my group is still very much interested in muscle development, but I am more of a gene hunter these days). Everything is now in stasis until the pandemic passes. It has been difficult to be away from the lab – we have a number of manuscripts in their final stages that need experiments completing before submission and it is frustrating not being able to get these done. Although experiments have stopped, I am still supporting undergraduate students and courses that are now running online, and it’s the time of year for marking dissertations. I also have three young children thrown into my daily mix. They are kindly humouring me by allowing me to explain to them genetic variation using Lego (different coloured bricks for different traits), perform crude DNA extractions from strawberries, and help them mount samples from around the house/garden on slides for viewing down our microscope. Luckily the weather has been surprisingly good, so we have been out in the garden exercising, home-schooling and indulging in plenty of fresh air.
KL I now work in a laboratory in the China Zebrafish Resource Center in Wuhan, having moved back at the end of 2010. By chance, I left Wuhan 2 h before the city was shut, to unite with my family for Spring Festival. At that moment, we didn’t realise how serious this pandemic would be. In our small town in Hubei we self-quarantined at home for 49 days. We ordered supplies online and volunteers brought them to our door every 2-3 days. The institute was shut, with no staff in the labs. Part-time staff came briefly every 4 days to feed the fish. Now, I am back in Wuhan and preparing to return to work next week (30 March). We hope the city will be back to normal after 2 months cold shock.
SH Experiments have been completely shut down and one of my team is recovering from COVID-19, but our many lines of fish are still happy, as far as I am aware. Just like Kuoyu’s, our Biological Services staff are doing a heroic job in very difficult circumstances. My family has escaped to rural Wales, so for me it’s email, FaceTime and Microsoft Teams between spring birdsong and isolated walks – quite idyllic. I’ve volunteered to run PCRs in local hospitals, but no call yet. I feel for my colleagues stuck in small flats in London as we await the approaching medical storm.
Let’s get back to the paper then – what led you to study the roles of Fgf and Tbx in myogenesis?
SH I had known about the importance of Fgf in mesoderm patterning since the work of Kimelman and Kirschner in 1987, and then the finding by my KCL colleagues Kevin Griffin and Nigel Holder that Fgf signalling was important in zebrafish trunk myogenesis. Kevin and Nigel proposed there was a gene they called ‘no trunk’ (by analogy with ‘no tail’, which is what the zebrafish T gene was called at the time). Kevin went as a postdoc to Dave Kimelman’s lab and showed that no trunk really existed; it was tbx16. Tbx16 is mainly known for controlling gastrulation movements at trunk levels; it was originally discovered as a mutant called spadetail that lacks most dorsal trunk tissue, and Sharon Amacher and Chuck Kimmel had shown this nicely. During the course of our study, both Dave Kimelman’s and Sharon Amacher’s labs showed that various Tbx genes collaborate in the formation of dorsal mesoderm. And Stephen Devoto’s lab had also shown that Tbx6, a close relative of Tbx16, is a negative regulator of presomitic mesoderm myogenesis. So it was really a no-brainer to examine this in our myogenic context, particularly with Fiona Wardle as a neighbour.
DO When the work began nearly 20 years ago almost all the MRF loss-of-function analyses had been done in mouse. So it was unclear how general the role of MRFs was in vertebrate myogenesis, particularly bearing in mind that Myod is not required for most myogenesis in flies. Our early experiments used morpholinos because that was the cutting-edge technology at the time; nowadays, we would do it by CRISPR genome editing. I was excited to see that zebrafish MRFs were initiated in the absence of Hh signalling in anterior somites, just as our lab had seen working with Betsy Pownall in Xenopus and as Andy McMahon had shown in smoothened mutants in mouse. Strikingly, just as Anne-Gaëlle Borycki showed in mouse, I saw that Hh signalling was more important in more caudal somites, so I asked what other signalling might be important to turn on the MRFs in anterior somites. SU5402 had been found to block Fgf signalling and it worked. Monte Westerfield’s lab found and published the same thing before us, which was both disappointing and encouraging, and meant we had to do more to publish something meaningful. So, I had shown with loss- and gain-of-function experiments that Fgfs in the tailbud and in the base of the notochord were important for myogenesis. It was one chapter in my thesis – I left at this point and Kuoyu took over the project.
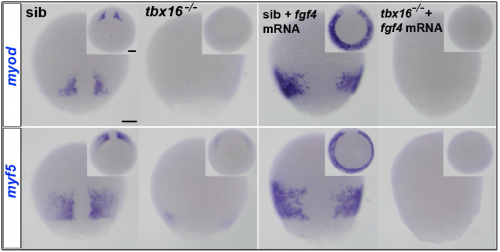
KL When I joined Simon’s lab, we wanted to pursue two aspects of Dan’s thesis. I looked downstream of MRFs at how they drove slow myogenesis; the first paper Dan and I published together was on how Cdkn1c (p57) cooperates with Myod to drive slow myogenesis. But I also began looking at how Fgf might activate MRF transcription. I found that MRFs were no longer induced by Fgf signalling in tbx16 mutants and in morpholino conditions, and that Fgf signalling cooperates with Tbx16 to drive MRF expression, which again argued that both Fgf and Tbx16 are needed within the presomitic tissue to initiate MRF expression and myogenesis.
SH At that point, the project came to a halt. But luckily, Fiona Wardle and colleagues had data on Tbx16 and Tbxta binding to myf5 and myod that moved the project forward.
MRF expression in tbx16 mutant embryos, with or without injected fgf4 mRNA.
Can you give us the key results of the paper in a paragraph?
The key result is that, in addition, to their role in paraxial mesoderm migration, Fgf signalling from the tailbud midline triggers Tbx16 to bind and directly activate the myf5 and myod genes. The same goes for Tbxta on the myod gene, and that initiates slow muscle fibre formation adjacent to the base of the forming notochord. Combined with Andrew and Fiona’s data on Tbx binding to the MRF loci, this encouraged Steve Cutty to use the glucocorticoid receptor-Tbx16 fusion protein plus dexamethasone to show that binding triggers myf5 and myod transcription directly, without new protein synthesis from other genes.
What do you think explains the differences in transcriptional targets between Tbx16 and Tbxta?
DO, KL & SH That’s an interesting question. The consensus binding sequences don’t seem to differ, so presumably their differential binding at certain sites in myf5 and myod has to do with collaborating accessory proteins and/or local chromatin structure. But another issue is whether they are competing for binding at sites where we detected both bound; in the embryo tbx16 and tbxta are expressed in only partially overlapping cell populations, so they could do different things in separate cell types. There is also the possibility that cell signalling and post-translational modifications could make them differentially active, even when bound to the same site. There’s a lot to work out if one wants to fully understand it.
What do your findings suggest about the evolution of vertebrate musculature?
DO, KL & SH It’s now clear that Tbxt/6/16 family genes are required in all major chordate lineages for myogenesis in the body and tail (but not the head). We think this is likely the ancestral way muscle was made in vertebrates. But several kinds of muscle are made in the somites of all vertebrates, and we think diversification of this Tbxt/6/16 gene family allowed the evolutionary diversification that gave vertebrates their advantage. For example, Tbxt genes are famous now for specifying notochord and thus controlling midline Hh expression. We think it is no coincidence that Hh induces both muscle and motoneuron diversification and that in the most primitive extant chordate, Amphioxus, notochord has muscle character. Perhaps the evolutionary origin of notochord was a special kind of dorsalmost muscle.
When doing the research, did you have any particular result or eureka moment that has stuck with you?
DO For me, the eureka moment that catalysed this project was finding that blocking Fgf signalling inhibited early myf5 and myod induction. I had spent a great deal of time analysing myf5 and myod expression in young but not older adaxial cells of Hh mutants. I was determined to find what midline-derived signals might be involved in regulating the initial Hh-independent MRF expression, and the localisation of Fgf signals made them particularly attractive candidates. Then finding that overexpression of either Fgf4 or Fgf6a could ectopically induce myf5 and myod was the icing on the cake – we knew we were onto something.
KL I think the best was finding that Tbx16 was essential for initial myf5 expression. I found it first with a morpholino; the mutant confirmed it. And then when Fgf4 overexpression could not rescue in the tbx16 mutant background.
And what about the flipside: any moments of frustration or despair?
DO Nothing that really stands out: although there are often such moments in science, it’s just the way it is. For this project it probably revolved around cloning – coming in early to find no colonies on my plates. That’s why it feels so good when experiments go to plan or give an unexpected exciting result. The bitter makes the sweet taste so much better!
KL I was very frustrated not to finish this Fgf story before leaving for China in 2010 due to my (or rather the UK Government’s) problem with visas. At that time, there was a temporary MRC funding hiatus caused by the 2008 financial crisis and the new Government was clamping down on immigrants like me. The result was that I had to leave the country before the end of the year. I finished what experiments I could over Christmas and flew back to China on 31 December 2010. Simon offered me a PhD place to continue the project, but my wife (also a scientist) was back in Wuhan starting her own lab, so in the end I stayed here. I now work in the China Zebrafish Resource Center as a member of the technical staff. My prime duty is to keep the aquarium running properly; no more experiments in my life, which is a great sadness to me.
The bitter makes the sweet taste so much better!
Finally, let’s move outside the lab – what do you like to do in your spare time in London and Wuhan?
DO Family is very important to me, especially with my children being so young (6-10 years). Most of my spare time revolves around taxiing between clubs. However, I have recently got into rock climbing (which the kids do too) and I love tinkering around on my motorbike when given half a chance.
KL I spend most of my spare time with my daughter. She is 4 years old. I teach her English and tell her stories about my life in London. She likes Pocoyo and learns a lot from this cute blue guy.
SH Though I enjoy London’s cultural offerings, my favourite spare time is spent in the hills, either the small ones surrounding our Welsh cottage, or larger ones in places like Argentina; they clear your head with a different kind of excitement and provide space to think. It has never been more important for people to (re-)connect with nature.
The Department of Biological Sciences at Louisiana State University invites applications for a tenure-track Assistant Professor position in all areas of stem cell and regenerative biology, including neurobiology, developmental biology, and molecular-cell biology. Biological Sciences is a large and dynamic department, with research ranging across all levels of biological organization, and the successful candidate will complement these strengths. Researchers utilizing non-traditional model organisms and those who would appreciate joining diverse and interactive faculty are especially encouraged to apply. In addition to the Department of Biological Sciences, opportunities for collaboration research are available at the LSU School of Veterinary Medicine, the LSU College of the Coast & Environment, the LSU College of Agriculture, and the LSU Pennington Biomedical Research Center. Successful candidates will be expected to establish and maintain a vigorous, extramurally funded research program and to contribute to undergraduate and graduate teaching. Applicants should have a Ph.D. in Biological Sciences or related field, a successful track record of productive research and publication, and postdoctoral experience.
Our department is dedicated to the goal of building a culturally diverse and pluralistic faculty, and we strongly encourage applications from women, minorities, individuals with disabilities, veterans, and other members of groups underrepresented in science. We seek candidates whose research, teaching, or service has prepared them to contribute to diversity and inclusion in higher education. Candidates should include a statement describing how they will promote an inclusive learning environment, and how their scholarship and mentoring practices support a diverse academic community.
The Sumigray lab in the Department of Genetics at Yale School of Medicine invites applications for a postdoctoral scientist to work on an exciting project at the interface of cell biology, morphogenesis and stem cell biology. The successful applicant will have a strong background in cell and developmental biology, genetics and/or stem cell biology and a track-record of original and impactful research in biomedical science.
The Sumigray lab studies the molecular and cellular processes that drive intestinal crypt morphogenesis and the importance of crypt architecture on intestinal stem cell activity/function. We use a combination of in vivo and in vitro models combined with live imaging, confocal microscopy and next-generation sequencing. For more information about the lab’s research, please see sumigraylab.org.
Applicants should possess a Ph.D. in molecular biology, cell biology, biochemistry, genetics, or a related field. Experience in mammalian cell culture is preferred but not required.
Please submit i) a cover letter with a brief description of your research experience and motivations for joining the lab, ii) CV and iii) contact details for 2-3 references to Dr. Kaelyn Sumigray (kaelyn.sumigray@yale.edu).
The Giraldez laboratory at Yale University is seeking to recruit a highly qualified Associate Research Scientist as a long-term scientist in the laboratory (www.giraldezlab.org). Prerequisites for appointment on the research scientist track include a doctoral degree and relevant postdoctoral experience.
The successful candidate will be a highly-motived scientist with excellent organizational, mentoring and leadership skills. They will be responsible for coordinating the overall scientific operations of the Giraldez lab and will provide critical training and mentoring to individual lab members. In addition, the successful candidate will have the opportunity to participate in multiple research projects and drive a scientific project aligned with the major interests of the laboratory. The successful candidate will have the following attributes:
A doctoral degree and relevant postdoctoral experience
Excellent interpersonal and communication skills
Excellent organizational skills and attention to detail
Solid publication record and the ability to drive long-term, successful research projects
Expertise in one or more of the following: molecular biology, chromatin biology, developmental
biology, genomics, and/or imaging
This appointment can be renewed indefinitely provided the need for the position continues, the funding for the position is available.
Tenure-track position in the Division of Biological Sciences
The Center for Craniofacial Molecular Biology (CCMB) of the Herman Ostrow School of Dentistry of the University of Southern California is recruiting outstanding candidates for a tenure-track position at the rank of Assistant Professor in the Division of Biomedical Sciences to conduct cutting-edge research in the areas of cell and developmental biology, tissue regeneration, cell signaling, gene regulation, computational modeling and human diseases using genetic and genomic approaches. At the CCMB, these disciplines focus on craniofacial biology and there are significant resources to support all aspects of our research. The University of Southern California offers an exciting, and highly supportive environment to conduct collaborative basic, clinical, and translational research. At the health science campus, faculty members at CCMB have access to all research centers and graduate students in all programs.
Candidates must have a Ph.D. in developmental biology, stem cell biology, or molecular biology. Candidates with both a Ph.D. and either a D.D.S. or D.M.D. degree are encouraged to apply. Candidates must have a strong record of high-quality research with significant publications in their field. Candidates with K99/R00 or other independent support are strongly encouraged to apply. For more information: https://usccareers.usc.edu/job/los-angeles/assistant-professor-of-dentistry/1209/17867982
Consideration of applicants will begin immediately and will continue until the position is filled. USC is an equal-opportunity educator and employer, proudly pluralistic and firmly committed to providing equal opportunity for outstanding persons of every race, gender, creed, and background. The University particularly encourages women, members of underrepresented groups, veterans, and individuals with disabilities to apply. USC will make reasonable accommodations for qualified individuals with known disabilities unless doing so would result in an undue hardship. Further information is available by contacting uschr@usc.edu.
 (No Ratings Yet)
(No Ratings Yet) (1 votes)
(1 votes)