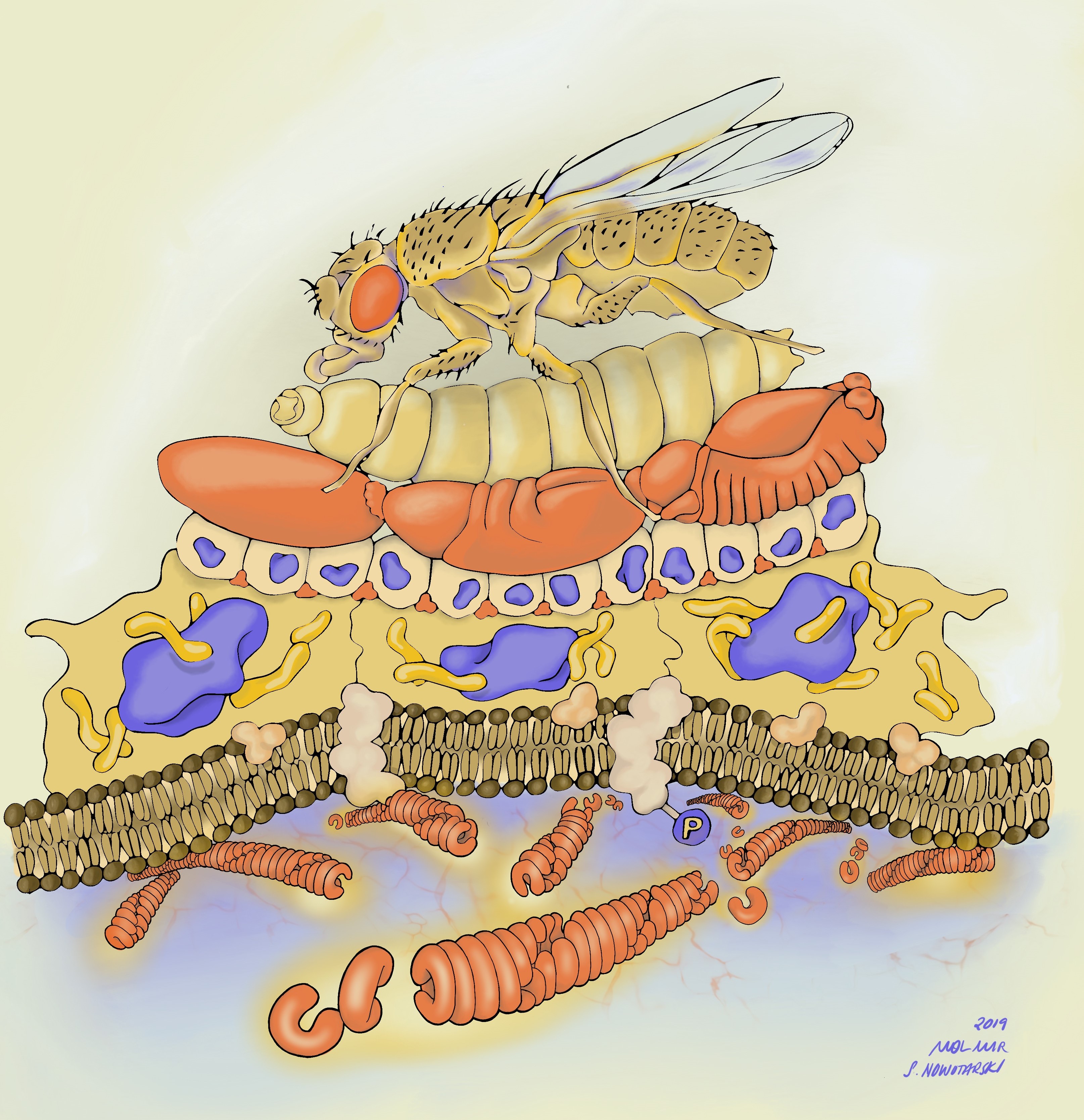
Post 1 requires experience in biochemistry, molecular and cell biology techniques as well as knowledge of transgenic and knockout mouse models to elucidate how NRP1 promotes brainvascularisation.
Post 2 requires experience in human embryonic stem cell culture and transcriptomics analyses to identify novel molecular and cellular markers for vascular progenitors in development and disease.
Post 2 will be co-supervised by Dr Paola Bonfanti at University College London’s GOS Institute of Child Health and the Francis Crick Institute.
Position Title: Research Assistant or Research Associate
Position Summary: The Marine Biological Laboratory seeks a motivated, creative, and innovative Research Assistant or Research Associate to join the laboratories of Kristin Gribble and David Mark Welch in the Josephine Bay Paul Center for Comparative Molecular Biology and Evolution. Our research combines comparative genomics, biochemistry, and life history to study aging, maternal effects, and DNA damage prevention and repair using rotifers, a novel aquatic invertebrate model system for studies of aging, neurobiology, genome evolution, and ecology.
The successful candidate will develop genome editing techniques in rotifers, including CRISPR/Cas9, as part of a broad initiative at the MBL to advance new aquatic and marine models for biological discovery. Research will take place in the Bay Paul Center, with extensive DNA sequencing and bioinformatic resources, and in the NSF-funded Genome Editing Facility in the Marine Resources Center, where MBL scientists are developing new genetic and genomic tools for a wide range of marine invertebrates. We invite individuals with experience in genome editing in other animals to join this expanding program.
Basic Qualifications: Research Assistant applicants should have a B.A., B.S., or Master’s degree in biology, cell/molecular biology, biochemistry, or a related field. Research Associate applicants should hold a Ph.D. or have commensurate laboratory experience. This position requires proficiency and previous experience in molecular biology, microscopy, microinjection, and CRISPR/Cas9 methodology. We are seeking an independent, organized, enthusiastic, and productive individual with robust problem solving skills. Excellent interpersonal skills, attention to detail, and a strong work ethic are essential. Position level and salary will depend upon education and experience.
Preferred Qualifications: The ideal candidate will have working familiarity with RNAi and transgenic protocols. Proficiency in bioinformatics is a plus. Previous experience in established animal model or in non-model systems is preferred.
Physical Requirements: Minimal exposure to biohazardous chemicals. Occasional lifting of heavy objects (<30 lbs).
Special Instructions: Please apply on the MBL website and submit the following three items with your application:
(1) Cover letter describing your experience, research goals, specific interest in joining our group, and what you would contribute to the project
(2) CV/resume
(3) Contact information for 3-4 references (Please do not send letters at this time; we will contact references directly).
PhD candidates on Quantitative Developmental Biology and Mathematical Modeling of Blood Vessel Growth near Tumorsat Leiden University, The Netherlands
Key responsibilities
The PhD projects will be part of an interdisciplinary project that will unravel how modifications of the extracellular matrix, as they can occur for example near tumors, can modify the structure of new blood vessel networks.
In this highly interdisciplinary project experimental biologists will work closely together with mathematical modelers on a daily basis. This approach enables the incremental development and testing of theories of single-cell behavior and collective cell behavior during tumor angiogenesis.
Based on cycles of iterative refinement of the mathematical model, followed by experimental validation, you will unravel aspects of blood vessel growth, and prepare your insights for publication in the biological, biophysical, and/or biomathematical literature.
One mathematical modeling project will focus primarily on the cellular scale and on the molecular mechanisms ofmechanical cell-cell interactions. The second mathematical modeling project will focus oncollective cell behavior, initially simplifying the underlying molecular details. The experimental project will focus on imaging single cell behavior and collective cell behavior during angiogenesis.
Our interdisciplinary team carries out mathematical biology research in close interaction with our recently established experimental lab. More information about the group can be foundhere. This interdisciplinary group is embedded at theMathematical Instituteand theInstitute of Biology Leiden, both at the Faculty of Science at Leiden University, and is physically based at, and embedded within the Cell Observatory of the Faculty of Science.
For more details about the positions and application procedure, see:
A minimum 2 year postdoctoral research position is available in the group of associate professor Thomas Elbenhardt Jensen, PhD at the department of Nutrition, Exercise and Sports, University of Copenhagen, currently #1 in Shanghai academic ranking of Sports Science Schools and Departments. Copenhagen is currently ranked #9 on the Global Liveability Index.
The overall goal of our research is to understand how physical activity and nutrition influence skeletal muscle function in health and disease (insulin resistance and atrophy) and the underlying molecular signal transduction mechanisms mediating these effects. Our model systems range from cell culture to mice to humans, which we combine with a unique mix of state-of-the-art methods within molecular cell biology and physiology.
Project
The currently available project seeks to develop and implement novel tools to study subcellular compartmentalized kinase signaling in adult skeletal muscle. This work will entail design and optimization of novel compartment-specific kinase-reporters, microscopy-approaches and compartment-isolation techniques in adult muscle.
The candidate
Qualified candidates should have strong experimental backgrounds in molecular and cell biology, in particular having expert knowledge within molecular cloning and microscopy. We seek a cell biologist with expertise in complementary, rather than similar, laboratory methods to those of our current team – specifically with know-how in creating new recombinant protein constructs. Therefore, prior research experience in skeletal muscle and mice is not a necessity and applicants without extensive experience in molecular cloning will not be considered.
The candidate should possess excellent written and verbal communication skills and have demonstrated the ability to think critically and innovatively. We are looking for an ambitious, flexible and self-motivated individual who is able to manage multidisciplinary projects both as lead scientist and as part of our research team and mentor junior lab members.
The postdoc’s duties will include research tasks as outlined above, as well as teaching. The post may also include performance of other duties.
The position is open from [1st of June 2020] or as soon as possible thereafter.
The University wishes our staff to reflect the diversity of society and thus welcomes applications from all qualified candidates regardless of personal background.
Terms of employment
The position is covered by the Memorandum on Job Structure for Academic Staff.
Terms of appointment and payment accord to the agreement between the Ministry of Finance and The Danish Confederation of Professional Associations on Academics in the State.
The starting salary is currently up to DKK 434.179 including annual supplement (+ pension up to DKK 74.244). Negotiation for salary supplement is possible.
The application, in English, must be submitted electronically by clicking APPLY NOW below.
Please include
Curriculum vitae
Diplomas (Master and PhD degree or equivalent)
A 1p motivational letter
Complete publication list
Separate reprints of 3 particularly relevant papers
The deadline for applications is March 17th 2020, 23:59 GMT +1.
Interviews will be held on April 6th 2020.
After the expiry of the deadline for applications, the authorized recruitment manager selects applicants for assessment on the advice of the Interview Committee.
Horizontal gene transfer has played a major role in the evolution of bacteria, however, its importance in animals is less clear. Recent evidence suggests that insects have repeatedly acquired genes from symbiotic bacteria that have provided them with key adaptation that has led to their success. Examples of this include, the horizontal acquisition of genes for lignocellulose digestion that has led to herbivory in beetles, and toxin encoding genes that may underlie protection from natural enemies in aphids and vinegar flies. However, it is currently unclear how common the horizontal transfer of symbiont genes is across insect species, and in most cases whether the newly acquired genes actually provide insects with adaptive functions. It is crucial to understand the dynamics of horizontal gene transfer in insects as the acquisition of novel traits from microbes is transforming our view of how arthropods evolve. This is of particular important for insect pests as it may underlie their capacity to colonise new environments and resist efforts to control them.
The successful applicant will use several insect families (e.g. aphids, ants) as models to rigorously test hypotheses on horizontal gene transfer in the evolution of insects. This may include: using whole genome sequence data to explore horizontally acquired genes involved in defence and nutrition across phylogenetically diverse insect species; investigating gene expression profiles to determine gene activity; and performing functional assays to reveal whether horizontally acquired genes provide insects with novel adaptive functions.
• You will have access to large genetic databases, insect collections, in-house live organisms and cutting-edge research facilitties to fuel your investigation.
• You will gain experience of molecular/experimental biology, comparative/metagenomics, bioinformatics, and statistics (e.g. comparative phylogenetics).
• You will be encouraged to develop your own ideas and hypotheses.
Research Environment
The School of Biological and Chemical Sciences at Queen Mary is one of the UK’s elite research centres, according to the 2014 Research Excellence Framework (REF). We offer a multi-disciplinary research environment and have approximately 160 PhD students working on projects in the biological, chemical and psychological sciences. Our students have access to a variety of research facilities supported by experienced staff, as well as a range of student support services.
The successful applicant will enter a vibrant research environment, under the supervision of Dr Lee Henry and co-supervision of Prof Richard Nichols and Dr Chema Martin. The Henry lab is well equipped to carry out the proposed research with world-class facilities in genomics and molecular biology. The student will also have access to ample funds to facilitate the research through Dr. Henry’s external funding (~£1 million in the past ten years). Dr. Henry has an outstanding record with student supervision with a focus on students generating high impact first authorship publications.
Our PhD students become part of Queen Mary’s Doctoral College which provides training and development opportunities, advice on funding, and financial support for research. Our students also have access to a Researcher Development Programme designed to help recognise and develop key skills and attributes needed to effectively manage research, and to prepare and plan for the next stages of their career.
The PhD student will have access to large genetic database, field collected insect samples and live in-house organisms. They will gain experience in molecular techniques, bioinformatics, experimental/field biology, and statistics (e.g. GLMM, comparative phylogenetics).
Eligibility
Applications are invited from candidates with, or expecting to be awarded, at least an upper-second class bachelors degree, or equivalent qualification, in biological or computational sciences (or similar). A masters degree is desirable, but not essential.
Potential candidates may contact Dr Lee Henry with informal enquiries about the project (l.henry@qmul.ac.uk).
Formal applications must be submitted through our online form by the stated deadline and should include your CV, statement of purpose, previous degree certificates and transcripts, and two referees. Applicants are not required to write a research proposal to apply for this position.
This studentship is open to applicants worldwide and is funded by a Queen Mary University of London Principal’s Studentship. It will cover tuition fees, and provide an annual tax-free maintenance allowance for 3 years at the Research Council rate (£17,009 in 2019/20).
“It’s the cutest rodent I have ever seen, even cuter than a cuddly hamster, and it would be fun doing a rotation for the opportunity to work with this animal.” That was my thought, as a first-year graduate student, when I first heard Kim present her research on the development of digit loss in the jerboa. I was completely hooked by the jerboa’s fascinating biology, and the Cooper lab became my second family. I had no idea then how working with the challenges of a non-traditional species would also mature my scientific thinking.
The vertebrate form is incredibly diverse, as evident not only in the body shapes and sizes but also in the limbs. For example, bats have elaborated forelimbs capable of flight while snakes have lost all four limbs. Such diversity has long inspired scientists and provided opportunities to investigate both developmental and evolutionary processes. To understand the developmental programs that explain limb diversity, the Cooper lab uses the bipedal jerboa (Figure 1), a close relative of the mouse with very different hindlimbs. The jerboa hindlimb has lost the first and fifth toes, fused the three central metatarsals into a single bone, disproportionately elongated the foot, and lost all intrinsic foot muscles. These modifications of the limb form allow the jerboa to jump high and rapidly change orientation, an advantage to escape predators in its desert environment with sparse vegetation. The jerboa and mouse have similar forelimb morphology, which serves as a great internal control for comparative anatomy and genomics to identify potential mechanisms specific to hindlimb divergence.
Figure 1: The jerboa with its elongated hindlimb.
Our recent work, Tran et al.eLife 2019, showed that the normal developmental process of muscle loss in the jerboa is similar to pathological muscle atrophy. In this post, I will elaborate on our findings, sprinkled with scientific speculation, and discuss the experimental challenges that we encountered during our exploration of the fascinating biology of this non-traditional laboratory animal model.
In 1979, Susan Berman, then a graduate student at the University of Pittsburgh who later became faculty at College of the Holy Cross, published a description of the hindlimb skeletal and muscular anatomy of many bipedal rodents including several species of jerboas1. When I started my work in 2014, we knew from her work that the adult jerboa has no ‘intrinsic’ muscles within its feet that spread and draw together toes of more basally branching species. Instead, most jerboa species have robust tendons in each location that we presume stabilize the ankle when animals jump and land with great force. However, we knew nothing of the developmental process leading to the evolutionary loss of these muscles.
Jimmy Hu, a graduate student with Cliff Tabin while Kim was a postdoc, had recently shown that limb myoblasts require reception of Shh signal to complete migration into the distal limb, as Smoothened receptor loss-of-function in mouse muscle causes a complete absence of muscles in the hands and feet2. A similar failure of muscle progenitor cells to populate the distal jerboa hindlimb seemed a simple explanation for the adult phenotype. We thought this project might be ‘low-hanging fruit’ for our nascent lab. We were therefore surprised to find that not only do myoblasts reach the foot, they fuse, differentiate, and form sarcomeres before rapidly disappearing in the first week after birth.
We next assumed the muscles must have died. Programmed cell death eliminates ‘extra’ tissue to shape developing structures in many contexts, including the process that carves interdigital webbing away from the fingers and toes like pulling dough away from the edges of a cookie cutter. Convinced this must also be the case for developing muscle that disappears so quickly, I performed multiple assays at all stages of muscle loss fruitlessly searching for signs of compromised cell membrane, fragmented DNA, or immune cell recruitment. I’ve also looked at thousands of muscle cells by transmission electron microscopy, a classic approach used to describe programmed cell death before there were kits and antibodies. Although myofibers don’t look entirely normal (they have large lipid droplets and vacuoles, and the sarcomere disintegrates), the plasma membrane appears intact, the nucleus appears normal, and there is no immune cell infiltration suggesting these cells do not die. I thought that I must be doing something wrong, but I had included all the positive and negative controls I could think of and covered all the stages of muscle loss. The more data I amassed suggesting this simple hypothesis was invalid, the more frustrated I grew. In contrast, my advisor became more and more excited.
We found no evidence of apoptosis, necrosis, or macrophage infiltration in jerboa foot muscles. Left panels show transverse sections of third digit interosseous muscle in the P6 jerboa foot, right panels show positive controls. Figure 3 in Tran, et al. eLife 2019.
If muscle cells did not seem to die, what was their fate? We considered the possibility that muscle cells switch identity, which would explain the disappearance of muscle proteins. The robust tendon in the adult and the appearance of collagen-producing cells surrounding odd extra-cellular aggregates of muscle filament protein led us to hypothesize that perhaps muscle cells could become some type of collagen-producing connective tissue fibroblast.
To definitively show that muscle cells become another cell type, or that they die by an unusual mechanism we were unable to detect by common assays, we would need to lineage label jerboa foot muscle cells and follow their existence over time. For over three years, we tried viral infection and plasmid electroporation approaches using multiple ‘muscle-specific’ promoters to lineage trace muscle cells using a Cre/lox reporter system, but none was successful. There were two technical complications hindering our ability to successfully lineage label muscle cells, discussed briefly in the response to eLife reviewers; we were unable to label a sufficient number of cells in neonatal mouse or jerboa feet, and none of the promoters specifically labeled muscle in either species. We hope that our ongoing work utilizing single-nucleus sequencing of muscle and surrounding tissue can help shed light on the fate of muscle cells by reconstructing a ‘virtual lineage’ using global similarities in the transcriptome of heterogeneous cell types3. Though not as definitive as a lineage tracing experiment, this might provide valuable insight into the most likely fate of these cells.
During those years of trying to determine the fate of the muscle, I also took a parallel approach to learn more about the developmental trajectory of jerboa foot muscles and to identify the earliest signs of impending loss. I found that muscle sarcomeres, though assembled, never fully mature to align with one another and instead became disorganized into pools of filaments. We were curious to know if the process of muscle disassembly happened all at once or in some orderly fashion. My observation that Desmin was the first protein to lose its striated pattern was our first break in this challenging project, since this was similar to what was observed in conditions of skeletal muscle atrophy. Even though atrophy is typically considered a response to disease or injury, might it be possible that the ‘normal’ developmental process of jerboa foot muscle loss is similar to atrophy? Indeed, expression of genes associated with pathological muscle atrophy, called “atrogenes,” was significantly higher in jerboa foot muscles than other ‘typical-developing’ muscles.
In 1883, Daniel John Cunningham, a Scottish physician, zoologist, and anatomist, reported the presence of intrinsic foot muscles in fetal horses, deer, and oxen that lack these muscles as adults. Cunningham observed fat and fibrotic tissue infiltration that replaced the disappearing foot muscles4. When we re-read the work of Cunningham as we prepared our own manuscript, we realized that Cunningham had left a hint we weren’t initially prepared to appreciate when he wrote “it is an instance of pathological change assisting a morphological process”4. Now aided with molecular and genetic tools, we show that the natural process of muscle loss in jerboa does indeed share molecular characteristics with pathological atrophy. If we look carefully at other ‘non-traditional species’, we may find that foot muscle atrophy in the jerboa is just one of many cellular responses associated with injury or disease in humans that is utilized in the normal development and physiology of other species.
While it is tempting to think that we might prove a cause and effect relationship by knocking down expression of the atrogenes to rescue jerboa foot muscle, it is important to consider differences between macroevolutionary phenotypes and mechanistically simpler phenotypes that are more typically studied in the lab. A comparison of the foot anatomy of living jerboas and their nearest relatives reveals that a subset of muscles was lost about 20 million years ago gradually leading to absence of all foot muscle in more recently derived jerboas. We therefore expect that multiple genetic changes contribute to the mechanism of muscle cell loss, perhaps including mutations that primarily affect integrated tissues like the skeleton and connective tissues. Although multiple genes responsible for evolutionary phenotypes can be identified by quantitative trait locus (QTL) analyses, the approach requires that species with different morphologies produce fertile hybrids. The jerboa lineage diversified too long ago for such mattings to be possible. Instead, we are currently applying interspecies differential RNA-Seq of mouse and jerboa foot and forearm muscles to identify genes with expression differences that are specifically associated with jerboa foot muscle loss. In addition to building a comprehensive molecular profile of this process, these data provide an opportunity to identify genes not previously known to influence muscle maturation and maintenance.
Thinking about the big picture of my research brought me to the realization that evo devo isn’t simply the study of individual phenomena in unusual critters. All deviations from ‘typical development’, both adaptive and maladaptive, provide insight into unifying principles. Studies of non-traditional species, while they can be technically challenging, will not only reveal the mechanisms of evolutionary malleability, but also advance our understanding of fundamental biological processes.
The development of an entire animal from a single totipotent cell – the zygote – is arguably one of the most fascinating processes in Nature. Strikingly, how this process is regulated at the lower levels of biological complexity, such as at the level of the genome and its regulation during the early phases of development, is still poorly understood. Moreover, our understanding relies on what is known for only a handful of species, such as mammals and flies. To solve this major knowledge gap, my lab is establishing annelid embryos as experimental systems to understand the genomic regulation of early animal development. Annelid embryos are unique in that they exhibit interspecific variation in the way the zygote gives rises to the major progenitor cells during early cleavage. What are the epigenetic mechanisms controlling annelid development? How do these mechanisms generate variability among species? How can annelid development inform us of the fundamental principles of animal embryogenesis?
In this project you will rigorously answer these questions combining state-of-the-art experimental and computation approaches.
You will have access to large genomic databases, and in-house live organisms to fuel your investigation.
You will gain experience of developmental biology and molecular techniques (gene expression analyses, epigenomics), bioinformatics (pipelines to analyse ATAC-seq, ChIP-seq and HiC), and statistics.
You will be encouraged to develop your own ideas and hypotheses.
This is an ERC-Starting Grant PhD funded position. The student will become part of Queen Mary’s Doctoral College, which provides training and development opportunities and financial support for research. The student will also have access to a Researcher Development Programme designed to help recognise and develop key skills and attributes needed to effectively manage research, and to prepare and plan for the next stages of their career.
Skills preferred
In a multidisciplinary project like this, candidates are unlikely to have a background in all disciplines involved. The most important qualification is motivation, enthusiasm and that the project appeals to you. However, previous computational experience would be a plus. We can envisage strong candidates coming through a variety of routes including:
– practical molecular biology
– developmental and cell biology
– computational biology
To apply, students should have a 1st class degree or have received a MSc in a relevant field (i.e. molecular biology, genetics, developmental and cell biology, bioinformatics) or are about to finish their MSc.
A new year is a good opportunity to look back on the one just passed, and whether you’re a Node regular or new to us, this selection of our 2019 highlights will hopefully be of interest, particularly if you’re interested in contributing something and looking for inspiration. We are a community site for and by developmental biologists – it’s free and easy to post, just sign up here. And if you don’t already, be sure to follow us on Twitter and Facebook. Happy new year!
Top 5 most-read posts published in 2019
Congratulations to Joachim Goedhart (twice, and for the second year in a row) and Shinichi Morita for making our top 5 most-read articles this year.
We love to hear the stories behind the latest papers – these might be personal recollections or more data-focused, and can take advantage of our informality and lack of word limit. Here’s ten of my favourites from 2019:
We’re now up to 42 in the series (Arabidopsis, Ascidian, Axolotl, Bat, Butterfly, C. elegans, Capitella, Chicken, Cnidaria, Colonial tunicate, Coral, Cricket, Ctenophore, Drosophila, ES cells, Gar, Gecko, Honeybee, Hydractinia, Lamprey, Larval-microbe, Maize, Marchantia, Mayfly, Moss, Mouse, Oikopleura, Onychophoran, Parhyale, Planaria, Platynereis, Rhino beetle, Sea urchin, Shark, Siphonophore, Skate, Spider, Sponge, Termite, Turtle, Xenopus, Zebrafish). Is your model missing? Get in touch.
Art and science
Annabel’s cover image
In March we posted a fascinating account from an artist (Gemma Anderson, University of Exeter) and a cultural historian of science (Janina Wellmann, MECS, Leuphana University Lüneburg) who visited the EMBL. Their outsider perspective on how science is done is well worth the read.
In October we interviewed Annabel Ebbing (the Hubrecht Institute in The Netherlands), first author of a Development paper on C. elegans neuroblast migration. She designed a beautiful piece of worm art that made Development’s cover – we heard about what science and art meant to her.
Science on tour
We published many meeting, workshop and course reports this year:
JanuaryThe launch of the Node Network, a global directory of developmental and stem cell biologists. You may have filled out our survey in the summer – the site is nearly there now and we’d love as many people as possible to sign up for it.
March(ish)We will run a community survey all about the Node. What can we do better in the coming years? What type of content do you want to see more (or less) of? What are we missing? Look our for our survey early in the year.
April Cast your mind back to April 1st, 2010 – Gordon Brown was UK Prime Minister, Clash of the Titans (eh?) topped the cinema charts, atmospheric carbon dioxide was 389ppm…and the Node was launched, publishing its first post (the first of 2403 and counting). Ten years on and it’ll be a good time to reflect on the past and look forward to the future. We’ll probably have some cake in the office and metaphorical cake on Twitter!
Department/Location: Wellcome Trust-Medical Research Council Stem Cell Institute
Salary: £32,816-£40,322
Reference: PS21993
Category: Research
Published: 6th January, Closing date: 5th February 2020
A postdoctoral Research Associate position is currently available for an individual to work in the laboratory of Prof. Anna Philpott within the Cambridge Stem Cell Institute (https://www.stemcells.cam.ac.uk/research/pis/philpott). The Philpott lab has broad interests in understanding the fundamental mechanisms that determine cell fate choice and differentiation during embryonic development and in cancers, as well as how these processes are co-ordinated with cell cycle progression.
The successful candidate will undertake a project focused around transcriptional regulation of lineage fidelity during fate specification and differentiation of mouse embryonic stem cells, focusing on uncovering epigenetic and co-factor-dependent mechanisms underlying these processes. There is also an opportunity to work on parallel mechanisms of fate specification and differentiation in Xenopus embryos. Within the laboratory, we use several experimental systems including mammalian embryonic stem cells, cancer cell culture, organoid systems as well as embryos of the frog Xenopus laevis. We use many techniques including genome-wide analysis of gene expression in single and multiple cells, chromatin binding and accessibility studies and crispr genome editing, alongside diverse biochemical approaches.
The successful candidate will have a PhD, considerable experience in stem cell biology, epigenetics, molecular biology, developmental biology, or a similar field, and a proven track record in scientific publication. Prior experience in mammalian cell culture is essential. Experience of epigenetics and/or transcriptional regulation are essential, while experience of genome-wide transcriptional analysis, and in particular analysis at the single cell level, would also be an advantage. Applicants must display an ability to undertake project management, work within a multi-disciplinary team environment, have good presentation and communication skills and the ability to contribute to an environment supporting researchers at all stages of their careers.
The Wellcome – MRC Cambridge Stem Cell Institute (CSCI) is a world-leading centre for stem cell research with the mission to transform human health through a deep understanding of stem cell biology. https://www.stemcells.cam.ac.uk/ . CSCI moved to the brand new, state of the art Jeffrey Cheah Biomedical Centre building on the Cambridge Biomedical Campus in summer 2019. The University actively supports equality, diversity and inclusion and encourages applications from all sections of society.
A 3-years-PhD position is available at the Chédotal Lab at the Vision Institute in Paris.
The goal of the PhD project will be to describe the organization of the developing human eye and characterize molecular and cellular mechanisms underlying human retinogenesis. We propose to revisit eye formation combining modern and innovative single-cell molecular and 3D imaging approaches on human embryo/fetal eye samples. Data from light sheet fluorescent microscopy (LSFM), spatial transcriptomics and in situ sequencing will be incorporated into one scaffold to provide a comprehensive 3D map of human retinogenesis. The project will provide significant and detailed insights into human ophthalmology, generating the first reference map of human retina development. It will further contribute to the Human Cell Atlas initiative. The thesis will be directed by Dr Alain Chédotal.
An experience in bioinformatics and transcriptomics or 3D imaging would be a plus but is not required.
Candidates should hold a diploma or a master’s degree in Neuroscience, Developmental Biology, Microscopy or Bioinformatics
They may send their application (in a PDF-format) to Dr Alain Chédotal (alain.chedotal@inserm.fr), including a CV and contact information for two references.
 (No Ratings Yet)
(No Ratings Yet)

 (2 votes)
(2 votes)