December in preprints
Posted by the Node, on 3 January 2020
Welcome to our monthly trawl for developmental biology (and related) preprints.
Here’s the last cache of preprints of 2019 – happy preprinting in 2020! They were hosted on bioRxiv and arXiv. Let us know if we missed anything. Use these links to get to the section you want:
Developmental biology
| Stem cells, regeneration & disease modelling
Evo-devo & evo
Cell biology
Modelling
Tools & resources
Research practice & education
Why not…
Developmental biology
| Patterning & signalling
Interdependent regulation of stereotyped and stochastic photoreceptor fates in the fly eye
Adam C. Miller, Elizabeth Urban, Eric L. Lyons, Tory G. Herman, Robert J. Johnston Jr.
Constraints and limitations on the transcriptional response downstream of the Bicoid morphogen gradient
Huy Tran, Aleksandra M. Walczak, Nathalie Dostatni
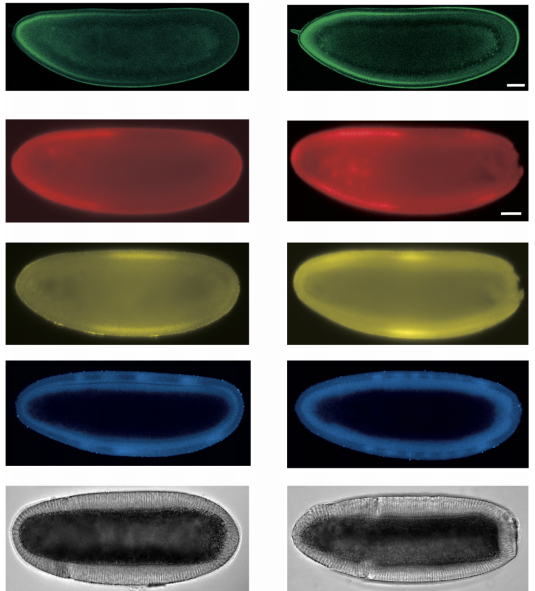
The dynamic transmission of positional information in stau- mutants during Drosophila embryogenesis
Zhe Yang, Hongcun Zhu, KaKit Kong, Jiayi Chen, Xiaxuan Wu, Peiyao Li, Jialong Jiang, Jingchao Zhao, Feng Liu
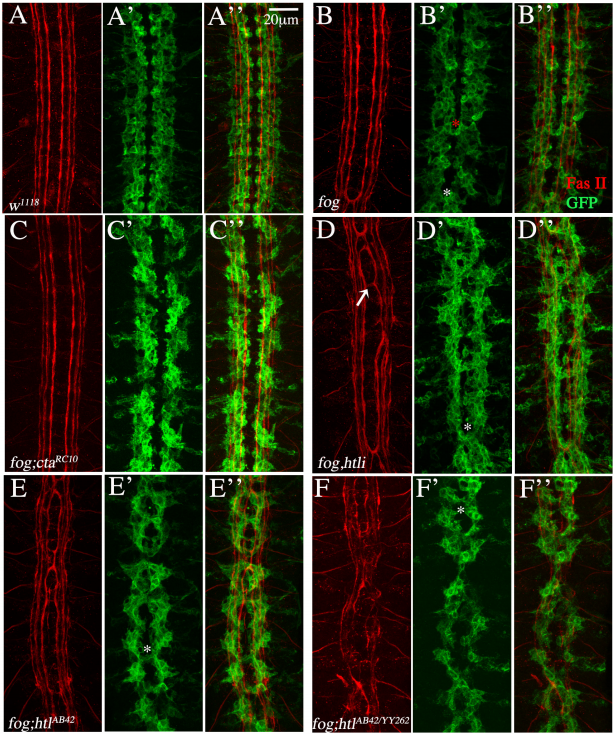
FGFR/Heartless and Smog interact synergistically to negatively regulate Fog mediated GPCR signaling
Kumari Shweta, Anagha Basargekar, Anuradha Ratnaparkhi
Modulation of Yorkie activity by alternative splicing is required for developmental stability
Diwas Srivastava, Marion de Toledo, Laurent Manchon, Jamal Tazi, François Juge
Neuropeptide F receptor acts in the Drosophila prothoracic gland to regulate growth and developmental timing
Jade R. Kannangara, Michelle A. Henstridge, Linda M. Parsons, Shu Kondo, Christen K. Mirth, Coral G. Warr
Endocycles support tissue growth and regeneration of the adult Drosophila accessory gland
Allison M. Box, Samuel Jaimian Church, David Hayes, Shyama Nandakumar, Russell S. Taichman, Laura Buttitta
Compartment and cell type-specific hypoxia responses in the developing Drosophila brain
Martin Baccino-Calace, Daniel Prieto, Rafael Cantera, Boris Egger
Presynaptic developmental plasticity allows robust sparse wiring of the Drosophila mushroom body
Najia A. Elkahlah, Jackson A. Rogow, Maria Ahmed, E. Josephine Clowney
Pegasus, a small extracellular peptide regulating the short-range diffusion of Wingless
Emile G Magny, Jose I Pueyo, Sarah A Bishop, Daniel Aguilar-Hidalgo, Juan Pablo Couso
An insulin, AMPK, and steroid hormone-mediated metabolic switch regulates the transition between growth and diapause in C. elegans
Sider Penkov, Bharath Kumar Raghuraman, Cihan Erkut, Jana Oertel, Roberta Galli, Eduardo Jacobo Miranda Ackerman, Daniela Vorkel, Jean-Marc Verbavatz, Edmund Koch, Karim Fahmy, Andrej Shevchenko, Teymuras V. Kurzchalia
The pattern of Nodal morphogen signaling is shaped by co-receptor expression
Nathan D. Lord, Adam N. Carte, Philip B. Abitua, Alexander F. Schier
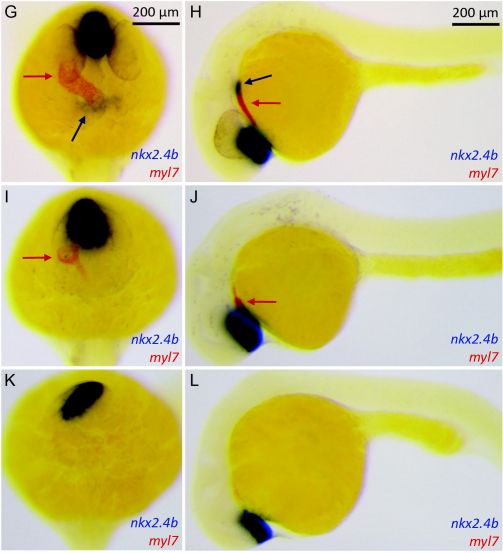
Enhanced canonical Wnt signaling during early zebrafish development perturbs the interaction of cardiac mesoderm and pharyngeal endoderm and causes thyroid specification defects
Isabelle Vandernoot, Benoît Haerlingen, Achim Trubiroha, Pierre Gillotay, Véronique Janssens, Robert Opitz, Sabine Costagliola
BACH family members regulate angiogenesis and lymphangiogenesis by modulating VEGFC expression
Batya Cohen, Hanoch Tempelhof, Tal Raz, Roni Oren, Julian Nicenboim, Filip Bochner, Ron Even, Adam Jelinski, Raya Eilam, Shifra Ben-Dor, Yoseph Adaddi, Ofra Golani, Shlomi Lazar, Karina Yaniv, Michal Neeman
Klf9 is a key feedforward regulator of the transcriptomic response to glucocorticoid receptor activity
Ian Gans, Ellen I. Hartig, Shusen Zhu, Andrea R. Tilden, Lucie Hutchins, Nathaniel Maki, Joel H. Graber, James A. Coffman
Copper Induces Zebrafish Central Neural System Myelin Defects: the Regulatory Mechanisms in Wnt/Notch-hoxb5b Signaling and Underlying DNA Methylation
Ting Zhang, PengPeng Guan, Guang Zhao, YaPing Fang, Hui Fu, Jian-Fang Gui, GuoLiang Li, Jing-Xia Liu
Cavefish Increase Red Blood Cell Development and Reprogram Metabolism as Adaptations to Environmental Hypoxia
Corine M. van der Weele, William R. Jeffery
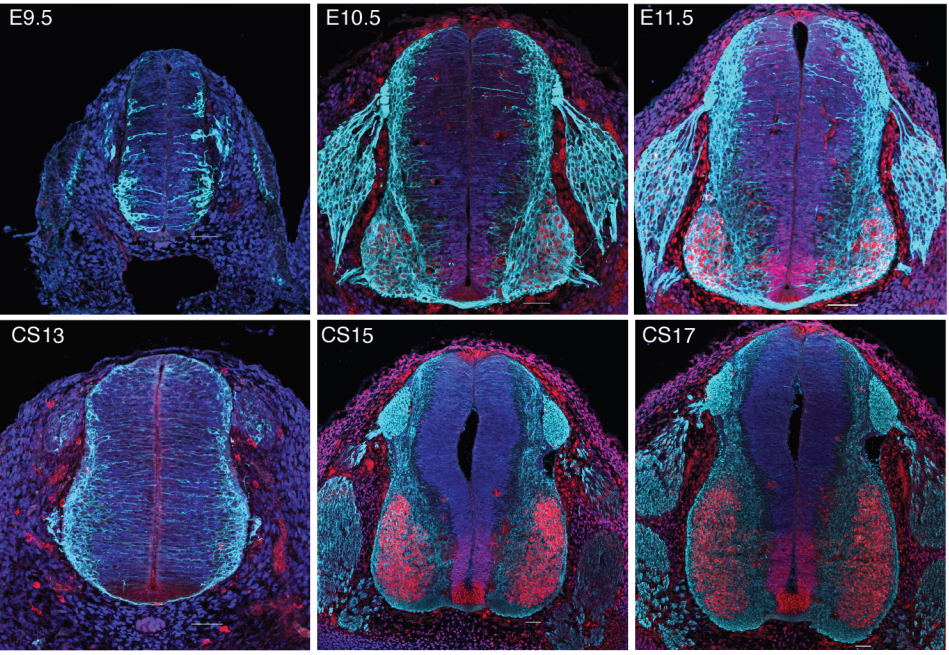
Species-specific developmental timing is associated with global differences in protein stability in mouse and human
Teresa Rayon, Despina Stamataki, Ruben Perez-Carrasco, Lorena Garcia-Perez, Christopher Barrington, Manuela Melchionda, Katherine Exelby, Victor Tybulewicz, Elizabeth M. C. Fisher, James Briscoe
The dynamics of oligodendrocyte generation: how distinct is the mouse from the human?
David G Gonsalvez, Georgina A Craig, Darragh M Walsh, Barry D Hughes, Rhiannon J Wood, Sang Won Yoo, Simon S Murray, Junhua Xiao
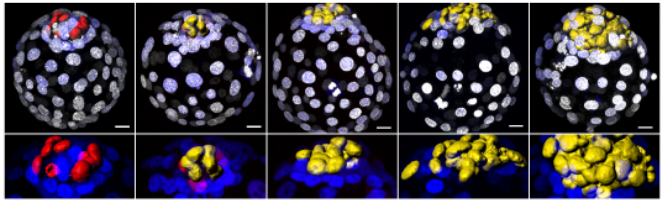
Growth factor-mediated coupling between lineage size and cell fate choice underlies robustness of mammalian development
Nestor Saiz, Laura Mora-Bitria, Shahadat Rahman, Hannah George, Jeremy P Herder, Jordi Garcia-Ojalvo, Anna-Katerina Hadjantonakis
FGF9 and FGF10 use distinct signaling pathways to direct lung epithelial specification and branching
Yongjun Yin, David M. Ornitz
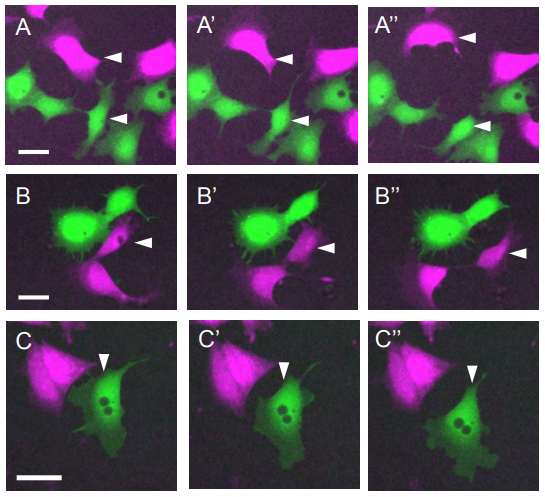
Role of SHIP2 in cell repulsion regulated by Eph receptor and ephrin signaling
Tim G. Ashlin, Zhonglin Wu, Qiling Xu, David G. Wilkinson
Nwd1 regulates neuronal differentiation and migration through purinosome formation in the developing cerebral cortex
Seiya Yamada, Ayaka Sato, Shin-ichi Sakakibara
Hominini-Specific Regulation of CBLN2 Increases Prefrontal Synaptogenesis
Mikihito Shibata, Kartik Pattabiraman, Sydney K. Muchnik, Nenad Sestan
Regulation of Prefrontal Patterning, Connectivity and Synaptogenesis by Retinoic Acid
Mikihito Shibata, Kartik Pattabiraman, Belen Lorente-Galdos, David Andrijevic, Xiaojun Xing, Andre M. M. Sousa, Gabriel Santpere, Nenad Sestan
Microglia depletion disrupts normal functional development of adult-born neurons in the olfactory bulb
Jenelle Wallace, Julia Lord, Lasse Dissing-Olesen, Beth Stevens, Venkatesh Murthy
Environmental Oxygen Regulates Astrocyte Proliferation to Guide Angiogenesis during Retinal Development
Robin M Perelli, Matthew L O’Sullivan, Samantha Zarnick, Jeremy N Kay
Circuit-specific dendritic development in the piriform cortex
Laura Moreno-Velasquez, Malte Kaehne, Hung Lo, Stephen Lenzi, Jörg Breustedt, Dietmar Schmitz, Sten Rüdiger, Friedrich W. Johenning
Oxytocin shapes spontaneous activity patterns in the developing visual cortex by activating somatostatin interneurons
Paloma P Maldonado, Alvaro Nuno-Perez, Jan Kirchner, Elizabeth Hammock, Julijana Gjorgjieva, Christian Lohmann
A SMAD1/5-YAP signaling module drives radial glial cell expansion and growth of the developing cerebral cortex
Sonia Najas, Isabel Pijuan, Anna Esteve-Codina, Susana Usieto, Juan D. Martinez, An Zwijsen, Maria L. Arbonés, Elisa Martí, Gwenvael Le Dréau
A switch in cilia-mediated Hedgehog signaling controls muscle stem cell quiescence and cell cycle progression
Sara Betania Cruz-Migoni, Kamalliawati Mohd Imran, Aysha Wahid, Oisharja Rahman, James Briscoe, Anne-Gaëlle Borycki
Association of Sonic Hedgehog with the Extracellular Matrix Requires its Putative Zinc-Peptidase Activity
Carina Jägers, Henk Roelink
Moonlighting α-PheRS connects JAK/STAT with Notch signaling for intestinal homeostasis
Manh Tin Ho, Jiongming Lu, Beat Suter
Niacin Stimulates Mammary Gland Development in Pubertal Mice through Activation of the AKT/mTOR and ERK1/2 Signaling Pathways
Yu Cao, Juxiong Liu, Lijun Ma, Qing Zhang, Jiaxin Wang, Wenjin Guo, Yanwei Li, Ji Cheng, Shoupeng Fu
Wwc2 is a novel mitotic/meiotic cell-cycle regulator and cell fate related gene, during preimplantation mouse embryo development and oogenesis
Giorgio Virnicchi, Pablo Bora, Lenka Gahurová, Andrej Šušor, Alexander W. Bruce
| Morphogenesis & mechanics
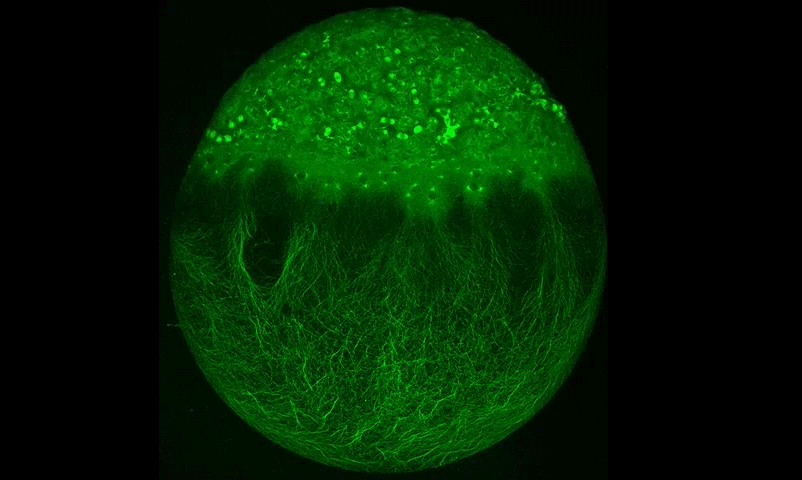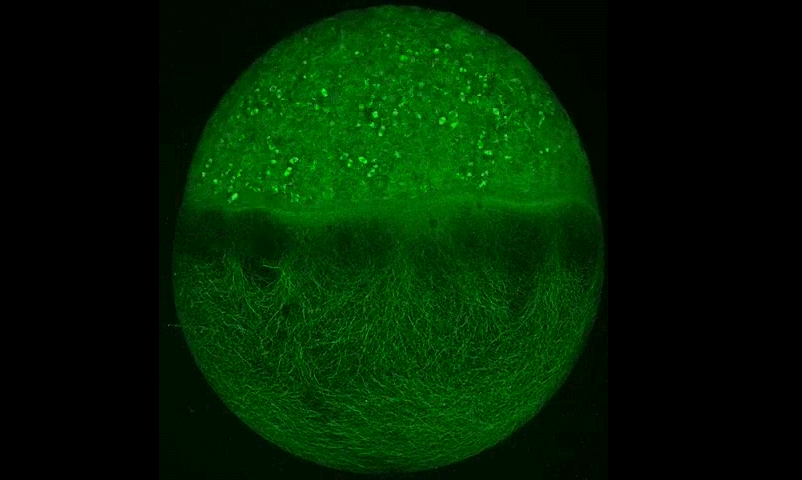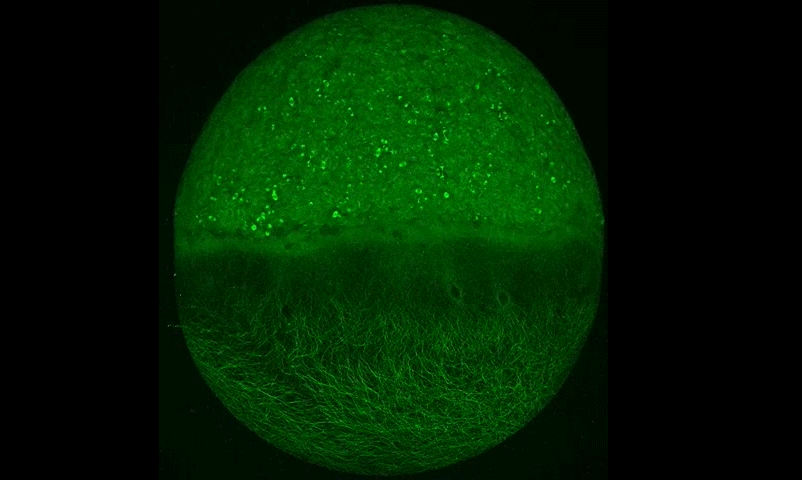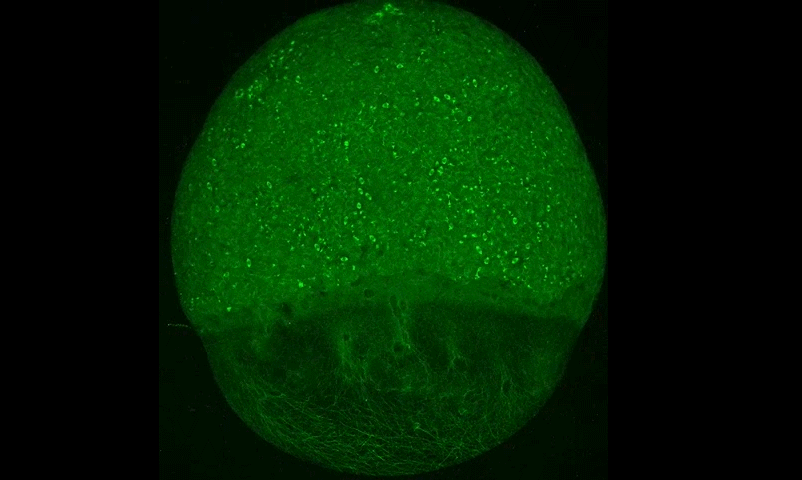
Cytoplasmic streaming drifts the polarity cue and specifies the cell polarity in Caenorhabditis elegans zygotes
Kenji Kimura, Akatsuki Kimura
Geometric cues stabilise long-axis polarisation of PAR protein patterns in C. elegans
Raphaela Geßele, Jacob Halatek, Laeschkir Würthner, Erwin Frey
RhoGAP RGA-8 supports morphogenesis in C. elegans by polarizing epithelia through CDC-42
Hamidah Raduwan, Shashikala Sasidharan, Luigy Cordova Burgos, Andre G. Wallace, Martha C. Soto
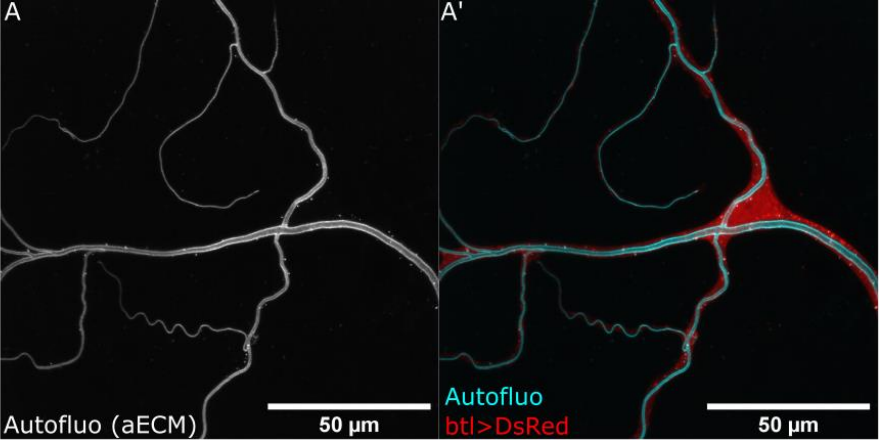
Multiple requirements for Rab GTPases in the development of Drosophila tracheal dorsal branches and terminal cells
Benedikt T. Best, Maria Leptin
broad controls leg imaginal disc morphogenesis in Drosophila via regulation of cell shape changes and remodeling of extracellular matrix
Clinton Rice, Stuart Macdonald, Xiaochen Wang, Robert E Ward IV
A polarized nucleus-cytoskeleton-ECM connection controls collective migration and cardioblasts number in Drosophila
C Dondi, B Bertin, JP Da Ponte, I Wojtowicz, K Jagla, G Junion
Apical Constriction Reversal upon Mitotic Entry Underlies Different Morphogenetic Outcomes of Cell Division
Clint S. Ko, Prateek Kalakuntla, Adam C. Martin
Lgl cortical dynamics are independent of binding to the Scrib-Dlg complex but require Dlg-dependent restriction of aPKC
Guilherme Ventura, Sofia Moreira, André Barros-Carvalho, Mariana Osswald, Eurico Morais-de-Sá
Distinct activities of Scrib module proteins organize epithelial polarity
Mark J. Khoury, David Bilder
Competition between kinesin-1 and myosin-V define Drosophila posterior determination
Wen Lu, Margot Lakonishok, Rong Liu, Neil Billington, Ashley Rich, Michael Glotzer, James R. Sellers, Vladimir I. Gelfand
The emergent Yo-yo movement of nuclei driven by collective cytoskeletal remodeling in pseudo-synchronous mitotic cycles
Zhiyi Lv, Jan Rosenbaum, Stephan Mohr, Xiaozhu Zhang, Deqing Kong, Helen Preiß, Sebastian Kruss, Karen Alim, Timo Aspelmeier, Jörg Großhans
Fibronectin-dependent tissue mechanics regulate the translation of segmentation clock oscillations into periodic somite formation
Patrícia Gomes de Almeida, Pedro Rifes, Ana Patrícia Martins-Jesus, Gonçalo G. Pinheiro, Raquel P. Andrade, Sólveig Thorsteinsdóttir
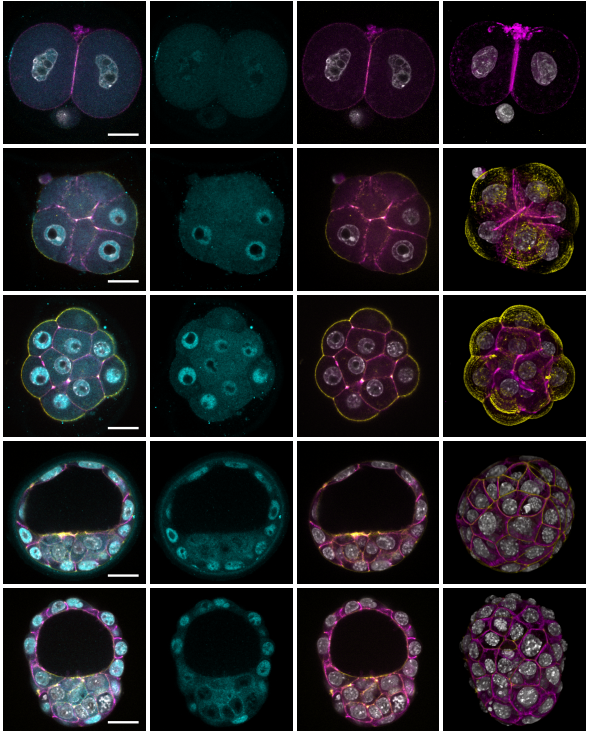
Position-sensing established during compaction dictates cell fate in the mammalian embryo
Christophe Royer, Karolis Leonavicius, Annemarie Kip, Deborah Fortin, Kirtirupa Nandi, Anna Vincent, Celine Jones, Tim Child, Kevin Coward, Chris Graham, Shankar Srinivas
Ectoderm to mesoderm transition by downregulation of actomyosin contractility
Leily Kashkooli, David Rozema, Lina Espejo-Ramirez, Paul Lasko, François Fagotto
Effects of nectin-3 misexpression on dendritic spine density during the postnatal development of layer 2/3 cortical neurons
Johanna Tomorsky, Philip R. L. Parker, Chris Q. Doe, Cristopher M. Niell
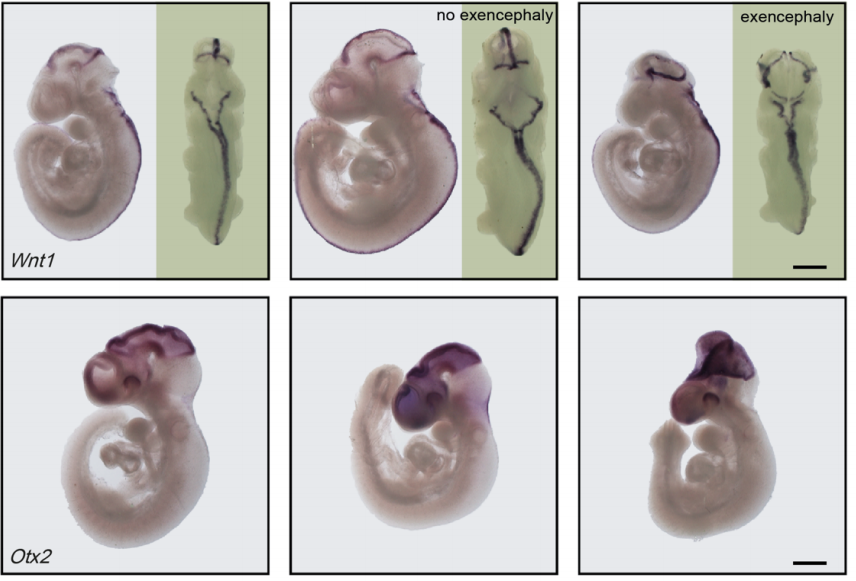
MLL4 is required for the first embryonic collective cell migration whereas MLL3 is not required until birth
Deepthi Ashokkumar, Qinyu Zhang, Christian Much, Anita S. Bledau, Jun Fu, Konstantinos Anastassiadis, A. Francis Stewart, Andrea Kranz
Thrombospondin-1 Promotes Circuit-Specific Synapse Formation via β1-Integrin
Sehwon Koh, Suva Roy, Oznur Eroglu, Samuel Strader, William J. Chen, Jeremy N. Kay, Greg D. Field, Cagla Eroglu
Laminin alpha 5 is Necessary for Mammary Epithelial Growth and Function by Maintaining Luminal Epithelial Cell Identity
Johanna I Englund, Hanne Cojoc, Leander Blaas, Alexandra Ritchie, Nalle Pentinmikko, Julia Döhla, Pauliina Munne, Manuel Patarroyo, Juha Klefström, Johanna Ivaska, Pekka Katajisto
IRSp53 shapes the plasma membrane and controls polarized transport at the nascent lumen during epithelial morphogenesis
Sara Bisi, Syed Abrar Rizvi, Stefano Marchesi, Davide Carra, Galina V. Beznoussenko, Ines Ferrara, Gianluca Deflorian, Alexander Mironov, Giovanni Bertalot, Federica Pisati, Amanda Oldani, Angela Cattaneo, Salvatore Pece, Giuseppe Viale, Angela Bachi, Claudio Tripodo, Giorgio Scita, Andrea Disanza
Polar pattern formation induced by contact following locomotion in a multicellular system
Masayuki Hayakawa, Tetsuya Hiraiwa, Yuko Wada, Hidekazu Kuwayama, Tatsuo Shibata
A Nodal/Eph signalling relay drives the transition from apical constriction to apico-basal shortening in ascidian endoderm invagination
Ulla-Maj Fiuza, Takefumi Negishi, Alice Rouan, Hitoyoshi Yasuo, Patrick Lemaire
| Genes & genomes

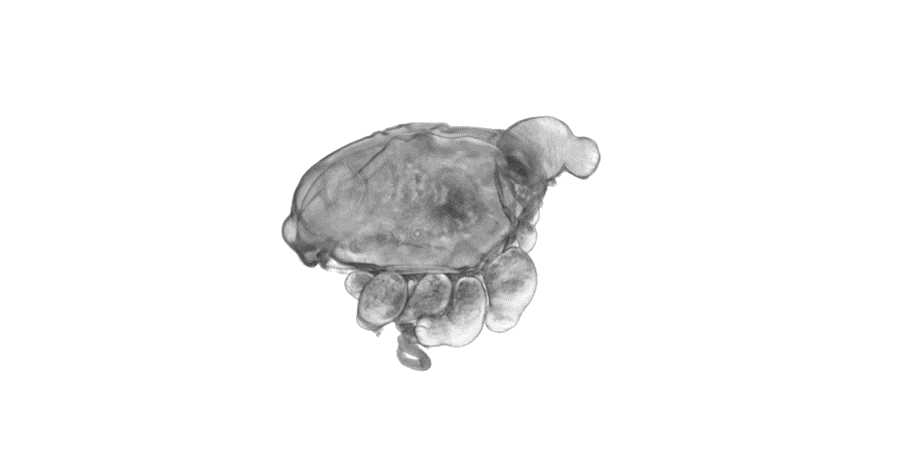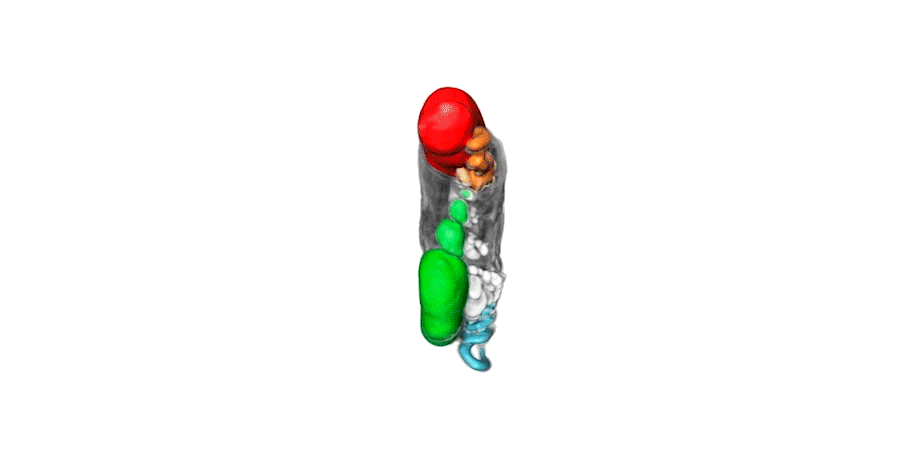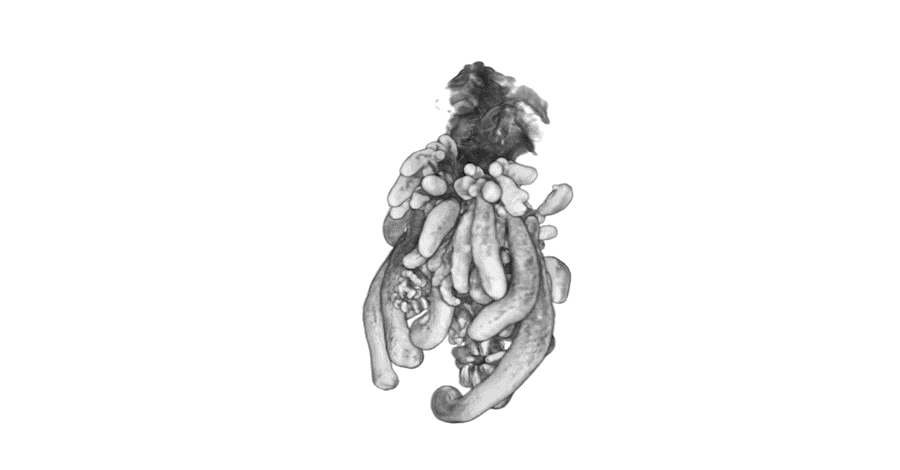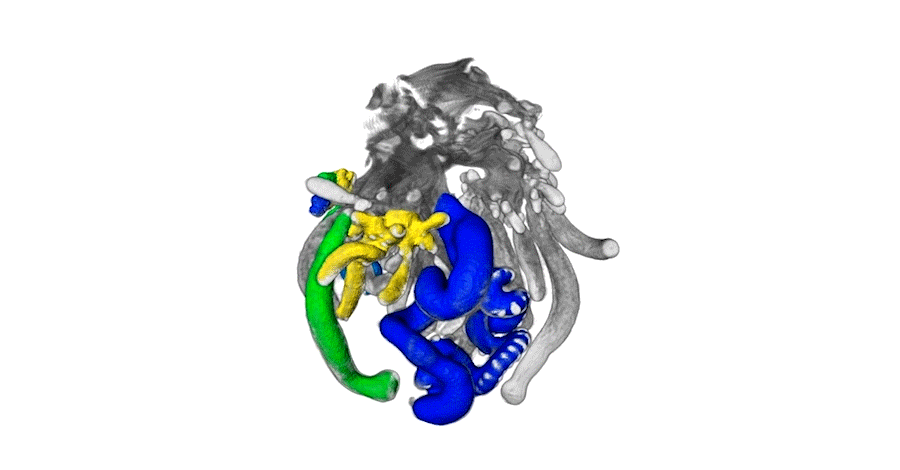
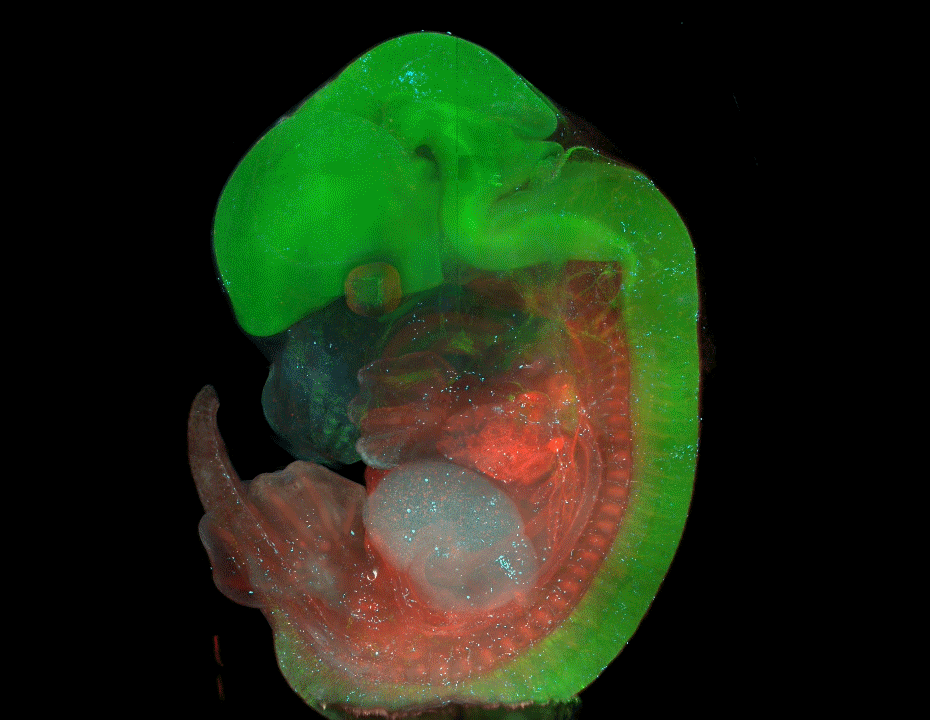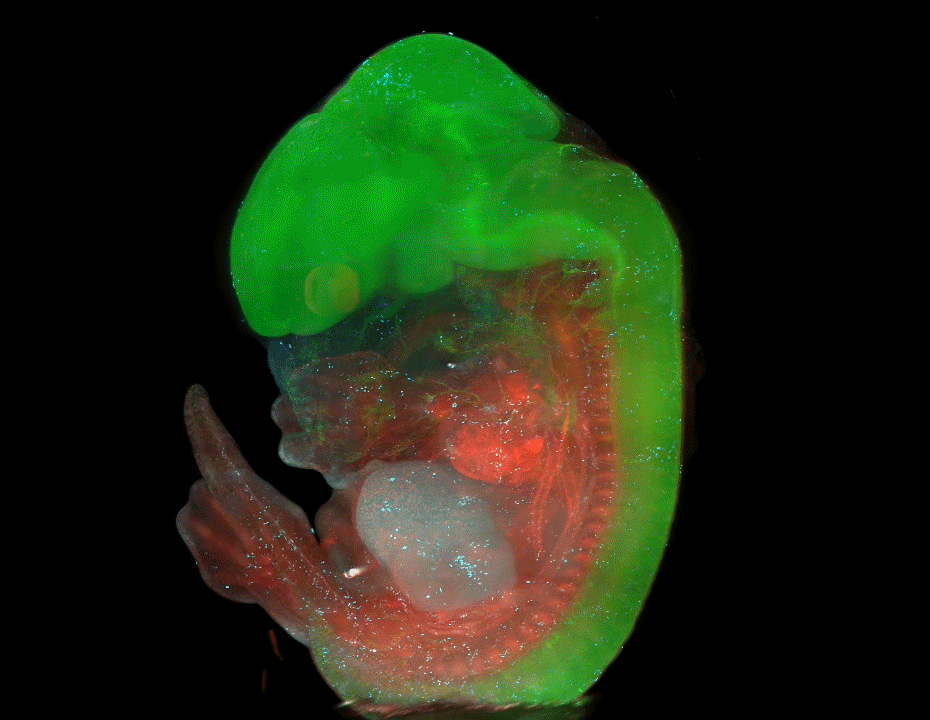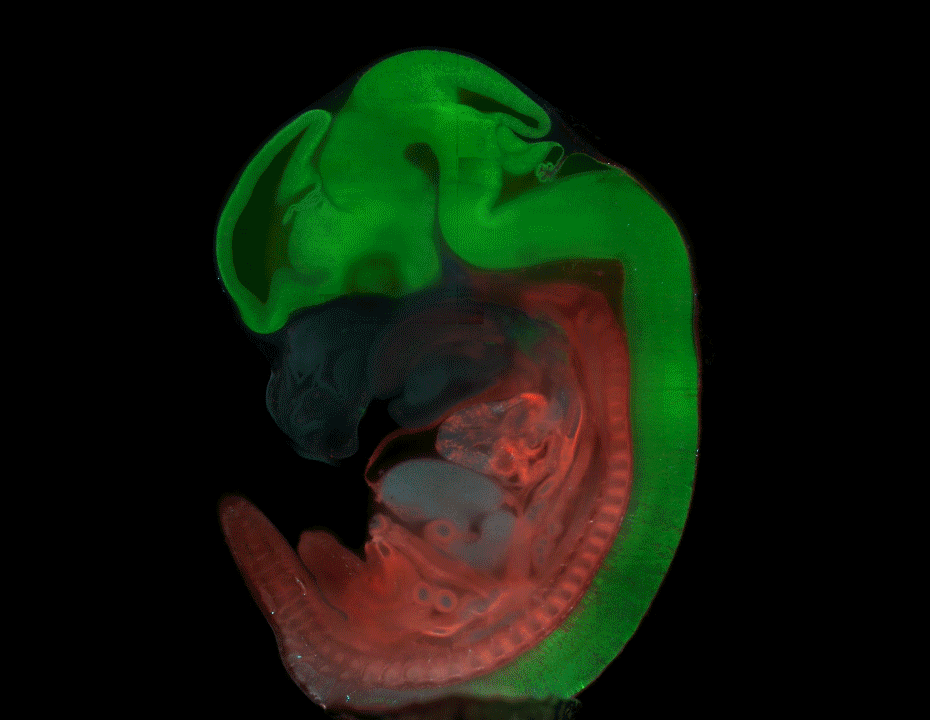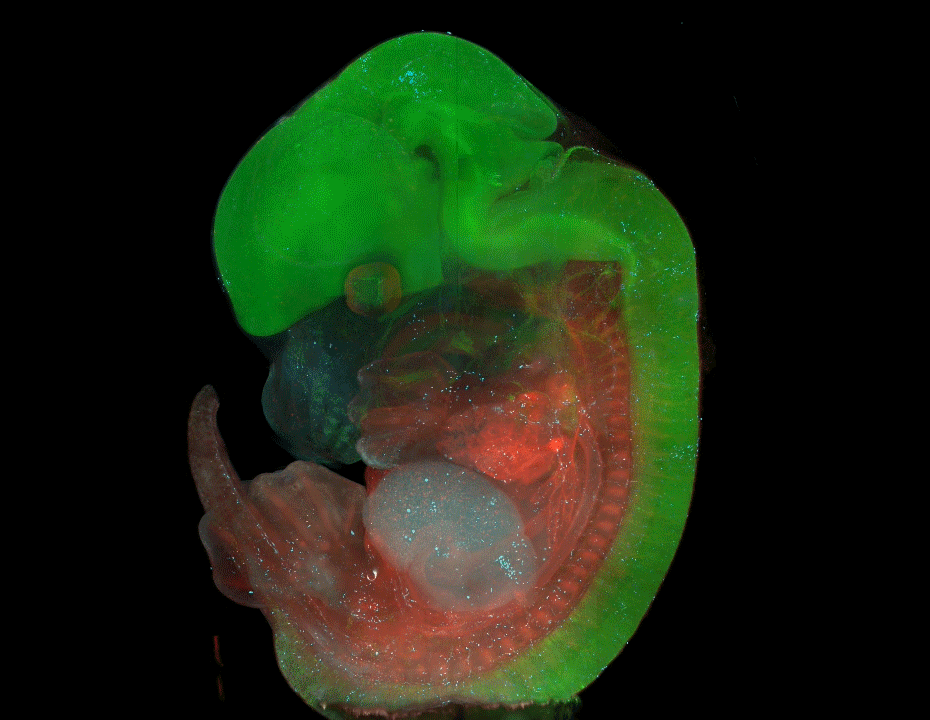
Nine-banded armadillo quadruplets from Balloiuz, et al.
The transcriptional legacy of developmental stochasticity
Sara Ballouz, Maria T. Pena, Frank M. Knight, Linda B. Adams, Jesse A. Gillis
Changing the Waddington landscape to control mesendoderm competence
James R. Valcourt, Roya Huang, Sharmistha Kundu, Divya Venkatasubramanian, Robert E. Kingston, Sharad Ramanathan
Zebrafish embryonic tissue differentiation is marked by concurrent cell cycle dynamic and gene promoter regulatory changes
Joseph W Wragg, Leonie Roos, Dunja Vucenovic, Nevena Cvetesic, Boris Lenhard, Ferenc Müller
HOX paralogs selectively convert binding of ubiquitous transcription factors into tissue-specific patterns of enhancer activation
Laure Bridoux, Peyman Zarrineh, Joshua Mallen, Mike Phuycharoen, Victor Latorre, Frank Ladam, Marta Losa, Charles Sagerstrom, Kimberley A. Mace, Magnus Rattray, Nicoletta Bobola
Hox binding specificity is directed by DNA sequence preferences and differential abilities to engage inaccessible chromatin
Milica Bulajić, Divyanshi Srivastava, Jeremy S Dasen, Hynek Wichterle, Shaun Mahony, Esteban O Mazzoni
Discovery of genes required for body axis and limb formation by global identification of conserved retinoic acid regulated enhancers and silencers
Marie Berenguer, Karolin F. Meyer, Jun Yin, Gregg Duester
Keystone genes of mammalian tooth patterning and quantification of their expression
Outi Hallikas, Rishi Das Roy, Mona M. Christensen, Elodie Renvoisé, Ana-Marija Sulic, Jukka Jernvall
Identification of Biomarkers Driving Blood Cell Development
Maryam Nazarieh, Volkhard Helms
The Human Accelerated Region HACNS1 modifies developmental gene expression in humanized mice
Emily V. Dutrow, Deena Emera, Kristina Yim, Severin Uebbing, Acadia A. Kocher, Martina Krenzer, Timothy Nottoli, Daniel B. Burkhardt, Smita Krishnaswamy, Angeliki Louvi, James P. Noonan
Cell type- and stage-specific expression of Otx2 is coordinated by a cohort of transcription factors and multiple cis-regulatory modules in the retina
Candace Chan, Nicolas Lonfat, Rong Zhao, Alexander Davis, Liang Li, Man-Ru Wu, Cheng-Hui Lin, Zhe Ji, Constance L. Cepko, Sui Wang
Mitf-family transcription factor function is required within cranial neural crest cells to promote choroid fissure closure
Katie L. Sinagoga, Alessandra M. Larimer-Picciani, Stephanie M. George, Samantha A. Spencer, James A. Lister, Jeffrey M. Gross
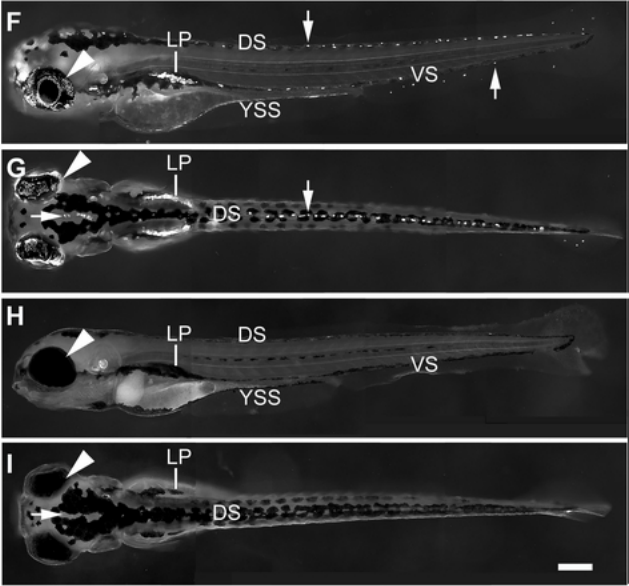
The MITF paralog tfec is required in neural crest development for fate specification of the iridophore lineage from a multipotent pigment cell progenitor
K. Petratou, S. A. Spencer, R. N. Kelsh, J. A. Lister
Sox8 and Sox9 act redundantly for ovarian-to-testicular fate reprogramming in the absence of R-spondin1 in mouse sex reversals
Nainoa Richardson, Isabelle Gillot, Elodie P. Gregoire, Sameh A. Youssef, Dirk G. de Rooij, Alain de Bruin, Marie-Cécile De Cian, Marie-Christine Chaboissier
Zebrafish dazl regulates cystogenesis upstream of the meiotic transition and germline stem cell specification and independent of meiotic checkpoints
Sylvain Bertho, Mara Clapp, Torsten U. Banisch, Jan Bandemer, Erez Raz, Florence L. Marlow
TWIST1 homodimers and heterodimers orchestrate lineage-specific differentiation
Xiaochen Fan, Ashley J. Waardenberg, Madeleine Demuth, Pierre Osteil, Jane Sun, David A.F. Loebel, Mark Graham, Patrick P.L. Tam, Nicolas Fossat
Deletion of a conserved Gata2 enhancer impairs haemogenic endothelium programming and adult haematopoiesis
Tomasz Dobrzycki, Christopher B. Mahony, Monika Krecsmarik, Cansu Koyunlar, Rossella Rispoli, Joke Peulen-Zink, Kirsten Gussinklo, Bakhta Fedlaoui, Emma de Pater, Roger Patient, Rui Monteiro
New observations on non-coding RNAs involved in the dual translation system in zebrafish development
Timo M. Breit, Johanna F. B. Pagano, Pjotr L. van der Jagt, Ellis Mittring, Wim A. Ensink, Marina van Olst, Selina van Leeuwen, Wim de Leeuw, Ulrike Nehrdich, Herman P. Spaink, Han Rauwerda, Rob J. Dekker
microRNA profiling of mouse cortical progenitors and neurons reveals miR-486-5p as a novel regulator of neurogenesis
Martina Dori, Daniel Cavalli, Mathias Lesche, Simone Massalini, Leila Haj Abdullah Alieh, Beatriz Cardoso de Toledo, Sharof Khudayberdiev, Gerhard Schratt, Andreas Dahl, Federico Calegari
Longitudinal epi-transcriptome profiling reveals the crucial role of m6A in prenatal skeletal muscle development of pigs
Xinxin Zhang, Yilong Yao, Jinghua Han, Yalan Yang, Yun Chen, Zhonglin Tang, Fei Gao
Massively parallel disruption of enhancers active during human corticogenesis
Evan Geller, Jake Gockley, Deena Emera, Severin Uebbing, Justin Cotney, James P. Noonan
Single cell epigenomic atlas of the developing human brain and organoids
Ryan S. Ziffra, Chang N. Kim, Amy Wilfert, Maximilian Haeussler, Alex M. Casella, Pawel F. Przytycki, Anat Kreimer, Katherine S. Pollard, Seth A. Ament, Evan E. Eichler, Nadav Ahituv, Tomasz J. Nowakowski
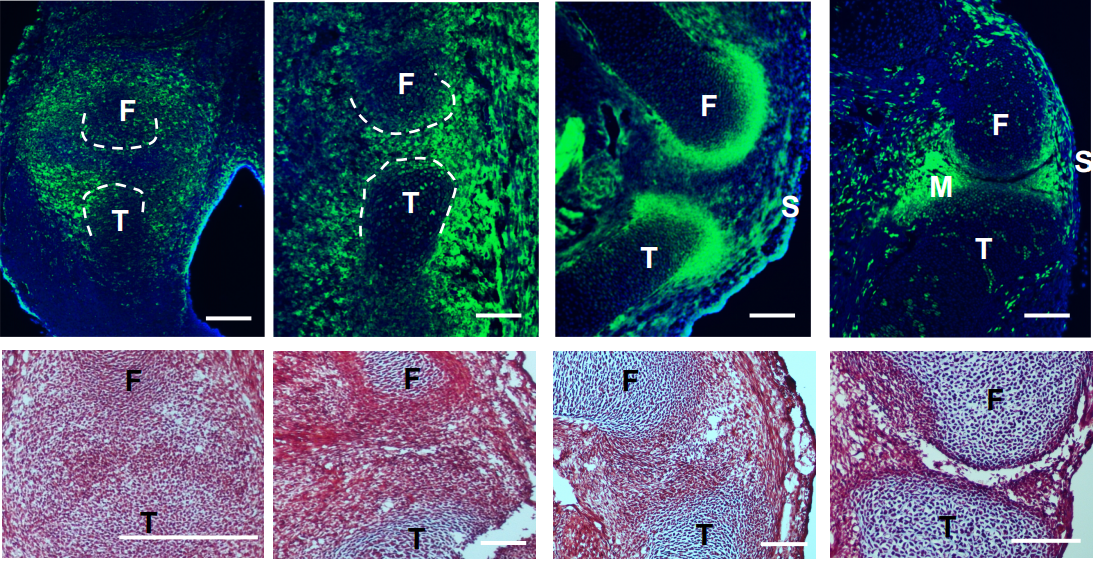
A single cell transcriptional atlas of early synovial joint development
Qin Bian, Yu-Hao Cheng, Jordan P Wilson, Dong Won Kim, Hong Wang, Seth Blackshaw, Patrick Cahan
Multiplexed single-cell transcriptomic analysis of normal and impaired lung development in the mouse
K. M. Hurskainen, I. Mižíková, D. P. Cook, C. Cyr-Depauw, F. Lesage, N. Andersson, E. Helle, L. Renesme, R.P. Jankov, M. Heikinheimo, B. C. Vanderhyden, B Thébaud
Chromatin regulatory dynamics of early development and regional specification in a directed differentiation model of the human small intestine
Yu-Han Hung, Sha Huang, Michael K. Dame, Jason R. Spence, Praveen Sethupathy
Polycomb-mediated repression compensates for loss of postnatal DNA methylation in excitatory neurons
Junhao Li, Antonio Pinto-Duarte, Mark Zander, Chi-Yu Lai, Julia Osteen, Linjing Fang, Chongyuan Luo, Jacinta D. Lucero, Rosa Gomez-Castanon, Joseph R. Nery, Isai Silva-Garcia, Yan Pang, Terrence J. Sejnowski, Susan B. Powell, Joseph R. Ecker, Eran A. Mukamel, M. Margarita Behrens
Conserved epigenetic regulatory logic infers genes governing cell identity
Woo Jun Shim, Enakshi Sinniah, Jun Xu, Burcu Vitrinel, Michael Alexanian, Gaia Andreoletti, Sophie Shen, Brad Balderson, Guangdun Peng, Naihe Jing, Yuliangzi Sun, Yash Chhabra, Yuliang Wang, Patrick P L Tam, Aaron Smith, Michael Piper, Lionel Christiaen, Quan Nguyen, Mikael Bodén, Nathan J. Palpant
poly(UG)-tailed RNAs in Genome Protection and Epigenetic Inheritance
Aditi Shukla, Jenny Yan, Daniel J. Pagano, Anne E. Dodson, Yuhan Fei, Josh Gorham, J.G. Seidman, Marvin Wickens, Scott Kennedy
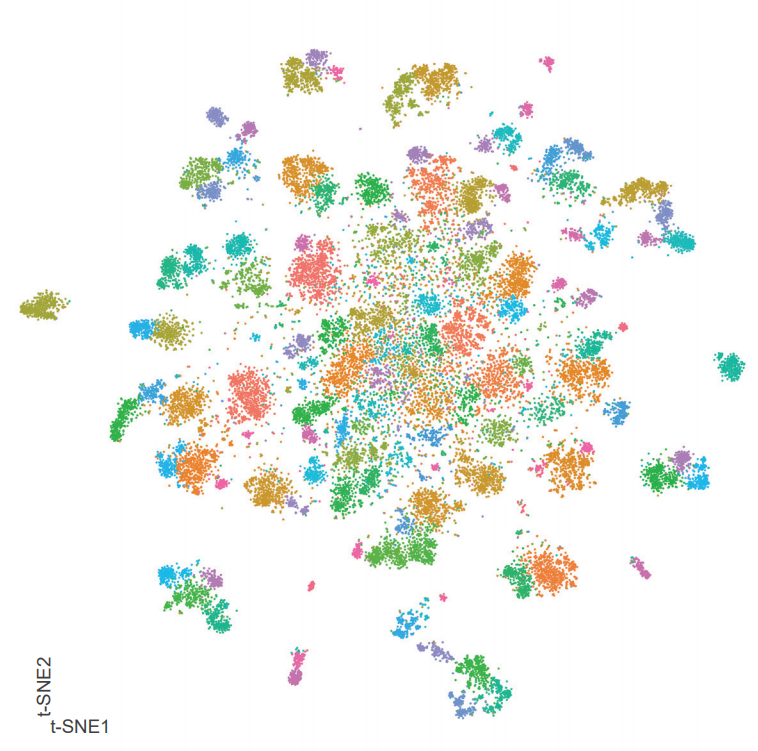
A single-cell transcriptomic atlas of the adult Drosophila ventral nerve cord
Aaron M. Allen, Megan C. Neville, Sebastian Birtles, Vincent Croset, Christoph D. Treiber, Scott Waddell, Stephen F. Goodwin
A single-cell survey of Drosophila blood
Sudhir Gopal Tattikota, Yanhui Hu, Yifang Liu, Bumsik Cho, Victor Barrera, Michael Steinbaugh, Sang-Ho Yoon, Aram Comjean, Fangge Li, Franz Dervis, Ruei-Jiun Hung, Jin-Wu Nam, Shannan Ho Sui, Jiwon Shim, Norbert Perrimon
Temporal specificity and heterogeneity of the fly immune cells’ transcriptional landscape
Pierre B. Cattenoz, Rosy Sakr, Alexia Pavlidaki, Claude Delaporte, Andrea Riba, Nacho Molina, Nivedita Hariharan, Tina Mukherjee, Angela Giangrande
sisterless A is required for the activation of Sex lethal in the germline
Raghav Goyal, Ellen Baxter, Mark Van Doren
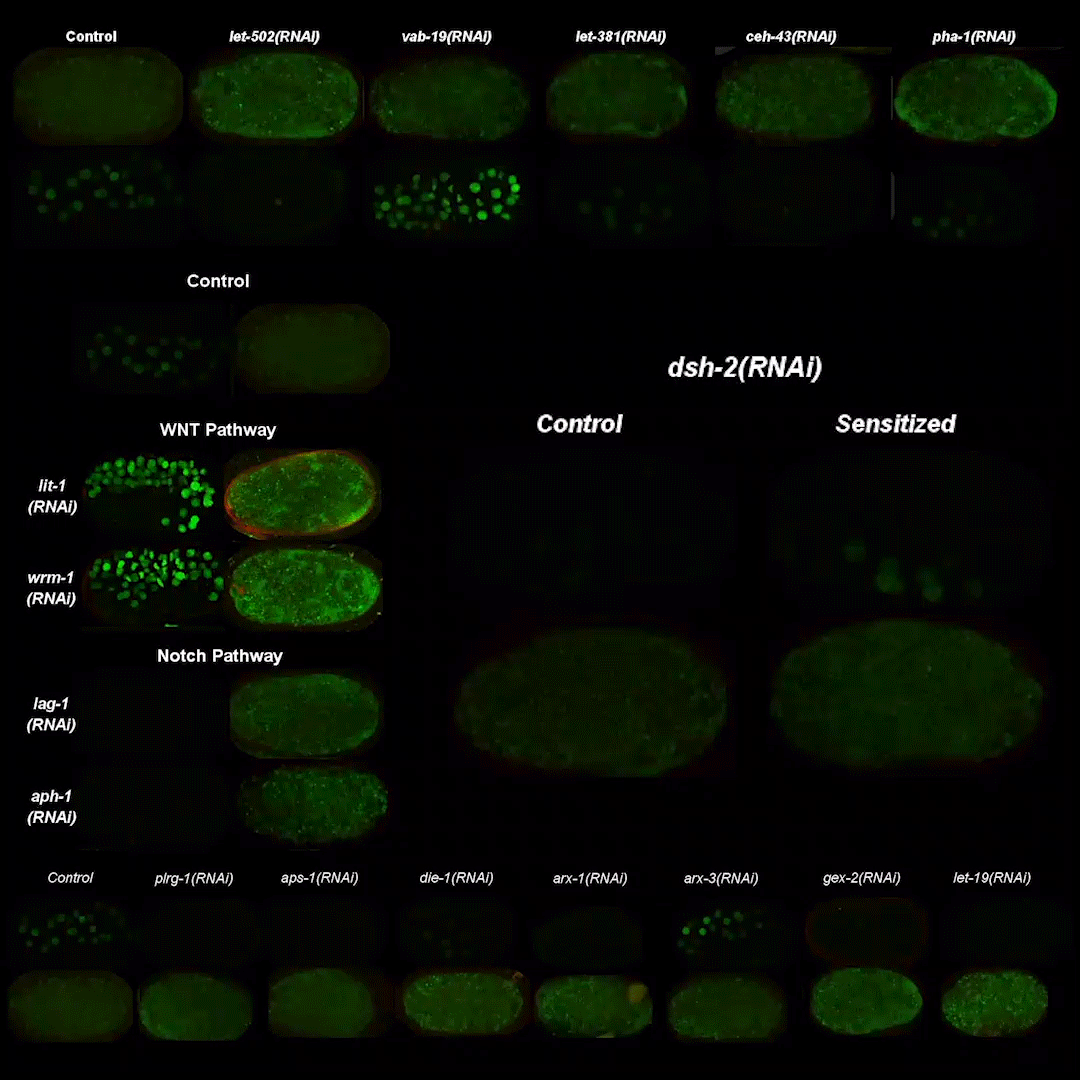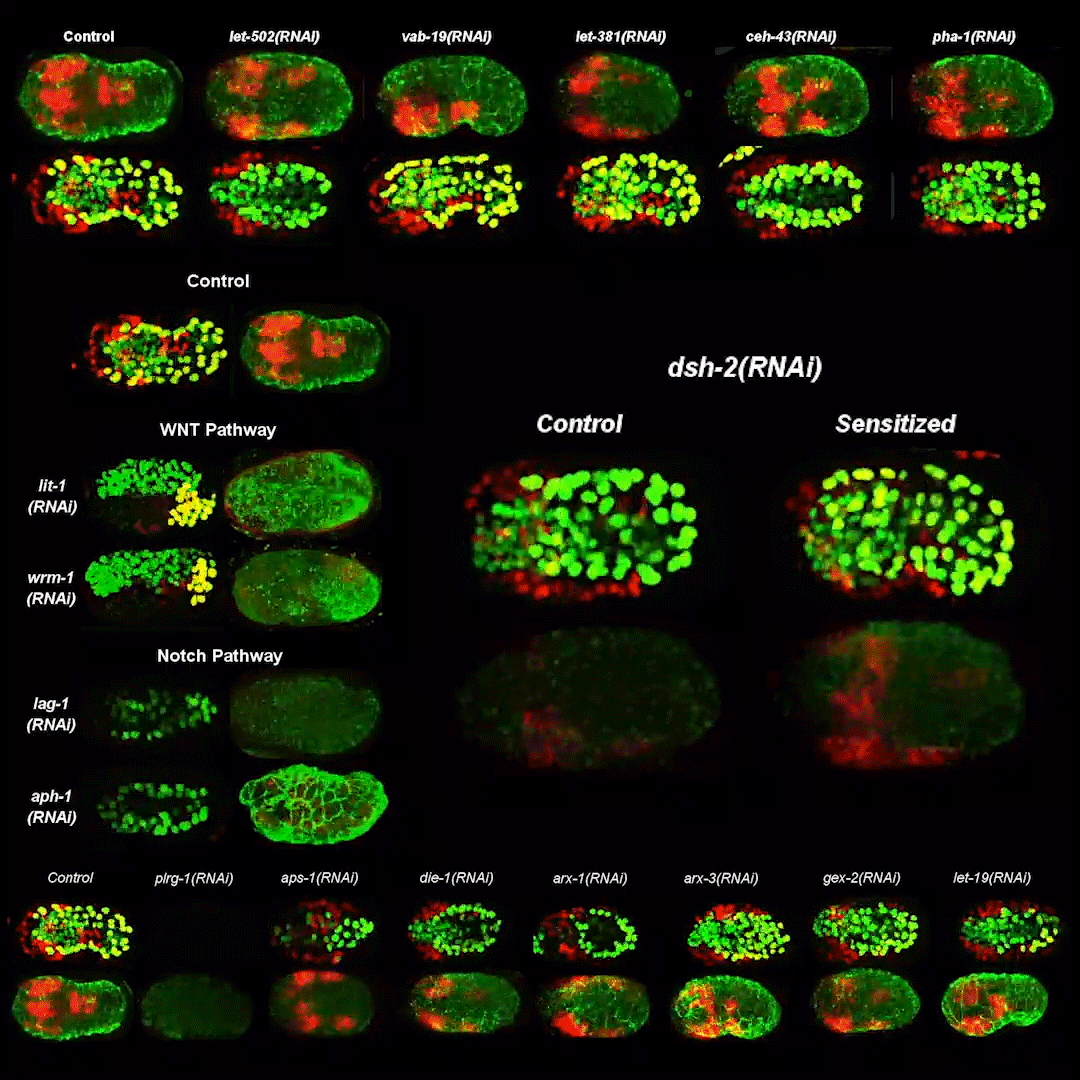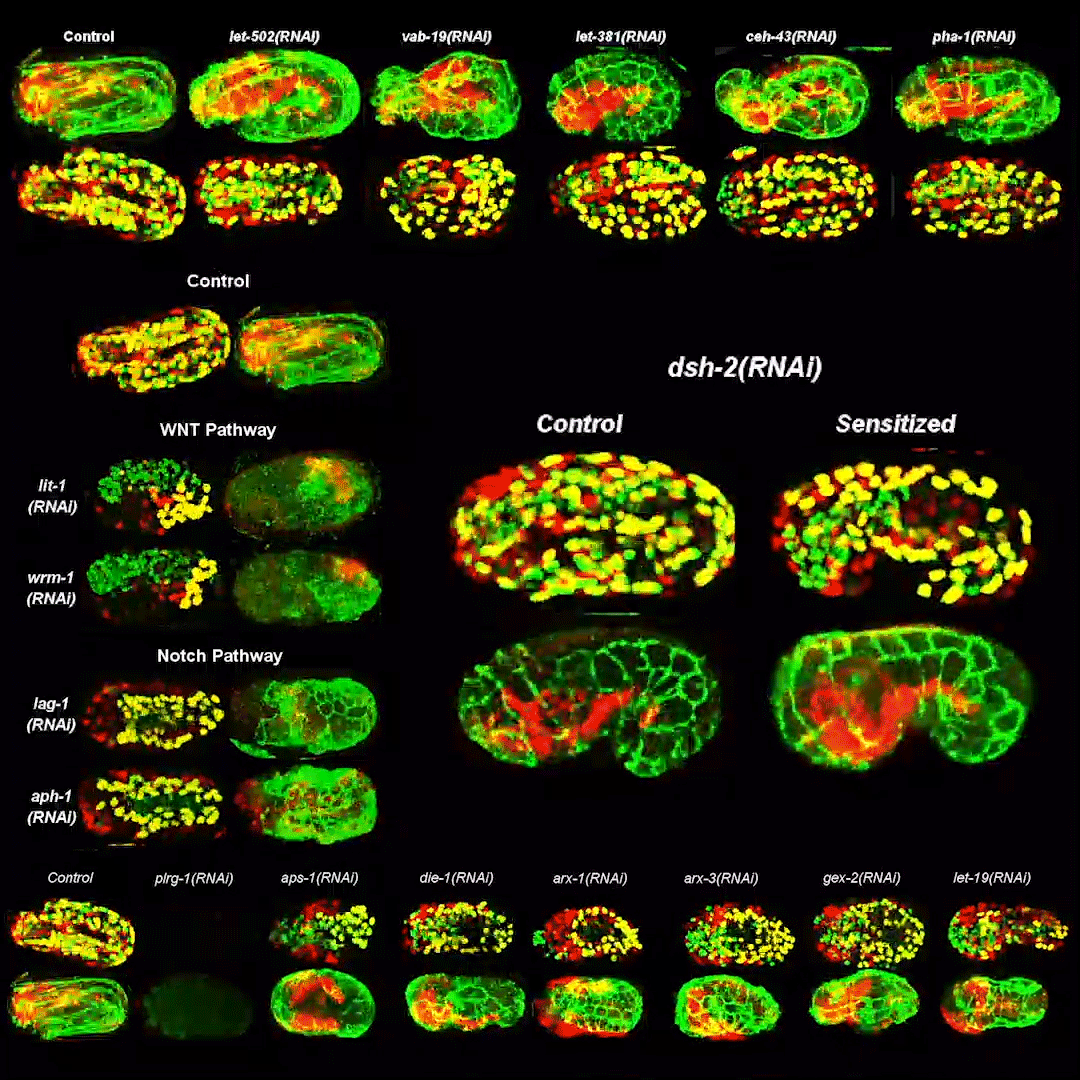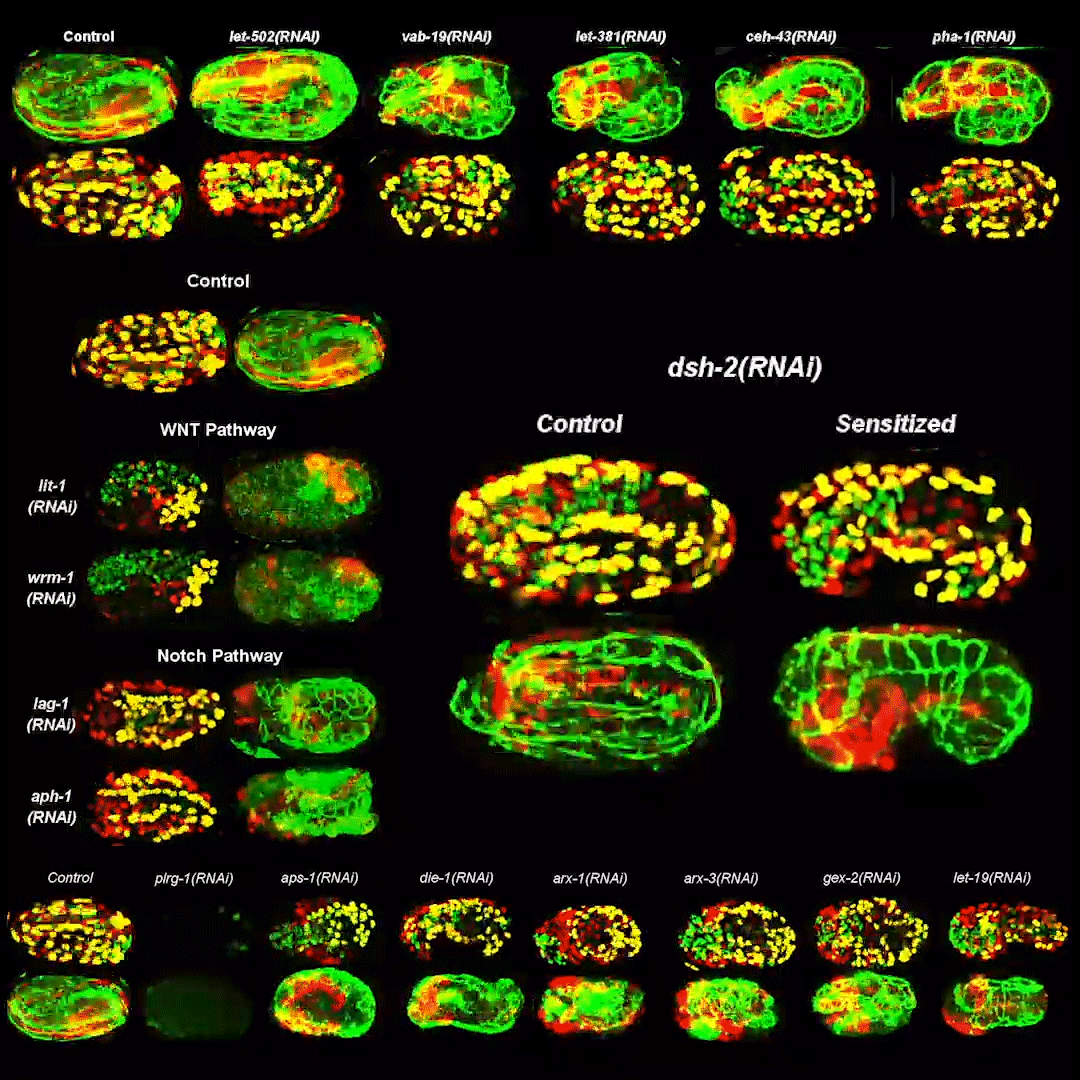
Precise temporal regulation of post-transcriptional repressors is required for an orderly Drosophila maternal-to-zygotic transition
Wen Xi Cao, Sarah Kabelitz, Meera Gupta, Eyan Yeung, Sichun Lin, Christiane Rammelt, Christian Ihling, Filip Pekovic, Timothy C. H. Low, Najeeb U. Siddiqui, Matthew H. K. Cheng, Stephane Angers, Craig A. Smibert, Martin Wühr, Elmar Wahle, Howard D. Lipshitz
Unc-4 acts to promote neuronal identity and development of the take-off circuit in the Drosophila CNS
Haluk Lacin, W. Ryan Williamson, Gwyneth M. Card, James B. Skeath, James W. Truman
Topology-driven analysis of protein-protein interaction networks detects functional genetic modules regulating reproductive capacity
Tarun Kumar, Leo Blondel, Cassandra G. Extavour
The Drosophila MLR COMPASS-like complex regulates bantam miRNA expression differentially in the context of cell fate
David J. Ford, Claudia B. Zraly, John Hertenstein Perez, Andrew K. Dingwall
| Stem cells, regeneration & disease modelling
The R2TP chaperone assembles cellular machineries in intestinal CBC stem cells and progenitors
Chloé Maurizy, Claire Abeza, Valérie Pinet, Marina Ferrand, Conception Paul, Julie Bremond, Francina Langa, François Gerbe, Philippe Jay, Céline Verheggen, Nicola Tinari, Dominique Helmlinger, Rossano Lattanzio, Edouard Bertrand, Michael Hahne, Bérengère Pradet-Balade
Notch ligand Dll4 impairs cell recruitment into aortic clusters and limits hematopoietic stem cells
Cristina Porcheri, Ohad Golan, Fernando J. Calero-Nieto, Roshana Thambyrajah, Cristina Ruiz-Herguido, Xiaonan Wang, Francesca Catto, Yolanda Guillen, Roshani Sinha, Jessica González, Sarah J. Kinston, Samanta A. Mariani, Antonio Maglitto, Chris Vink, Elaine Dzierzak, Pierre Charbord, Bertie Göttgens, Lluis Espinosa, David Sprinzak, Anna Bigas
The 9aaTAD activation domains in the four Yamanaka Oct4, Sox2, Myc, and Klf4 transcription factors essential during the stem cell development
Martin Piskacek, Kristina Jendruchova, Martina Rezacova, Marek Havelka, Norbert Gasparik, Alena Hofrova, Andrea Knight
TGFβ superfamily signaling regulates the state of human stem cell pluripotency and competency to create telencephalic organoids
Momoko Watanabe, Jillian R. Haney, Neda Vishlaghi, Felix Turcios, Jessie E. Buth, Wen Gu, Amanda J. Collier, Osvaldo A. Miranda, Di Chen, Shan Sabri, Amander T. Clark, Kathrin Plath, Heather R. Christofk, Michael J. Gandal, Bennett G. Novitch
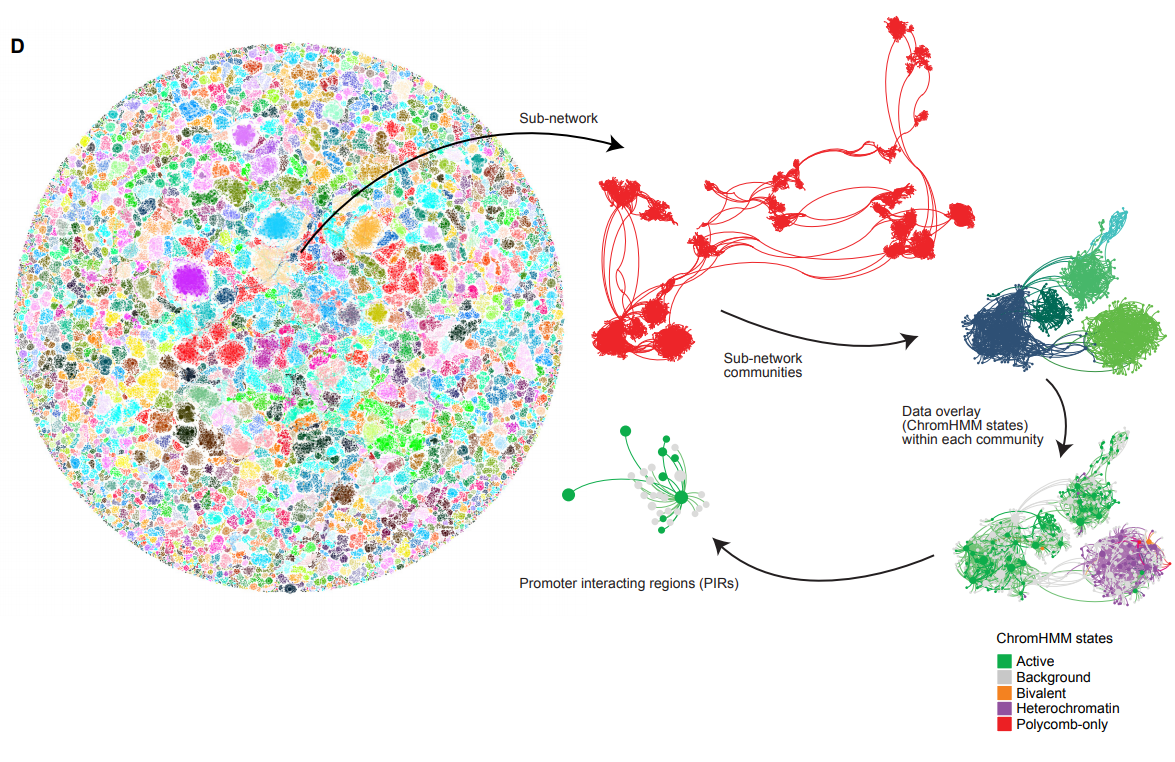
Network analysis of promoter interactions reveals the hierarchical differences in genome organisation between human pluripotent states
Peter Chovanec, Amanda J. Collier, Christel Krueger, Csilla Várnai, Stefan Schoenfelder, Anne Corcoran, Peter J. Rugg-Gunn
A Holistic Analysis of the Intestinal Stem Cell Niche Network
Darrick M. Hansen, Paloma Ivon Meneses Giles, Xi C. He, Shiyuan Chen, Ariel Paulson, Christopher M. Dekaney, Jennifer Wang, Deqing Hu, Aparna Venkatraman, Woosook Kim, John Kaddis, Barbara J. Olack, James C.Y. Dunn, Calvin Kuo, Susan Henning, Alan M. Hanash, Courtney W. Houchen, John Lynch, Martin G. Martin, Joyce C. Niland, Matthias Stelzner, Melissa Wong, Timothy C. Wang, Jian Yu, Kelley Yan, Linheng Li
Functional Mature Human Microglia Developed in Human iPSC Microglial Chimeric Mouse Brain
Ranjie Xu, Andrew J. Boreland, Xiaoxi Li, Anthony Posyton, Kelvin Kwan, Ronald P. Hart, Peng Jiang
Characterization of human-iPSCs derived spinal motor neurons by single-cell RNA sequencing
Louise Thiry, Regan Hamel, Stefano Pluchino, Thomas Durcan, Stefano Stifani
Transcriptome and Proteome analysis of Hemidactylus frenatus during initial stages of tail regeneration
Sai Pawan, Sarena Banu, Mohammed M Idris
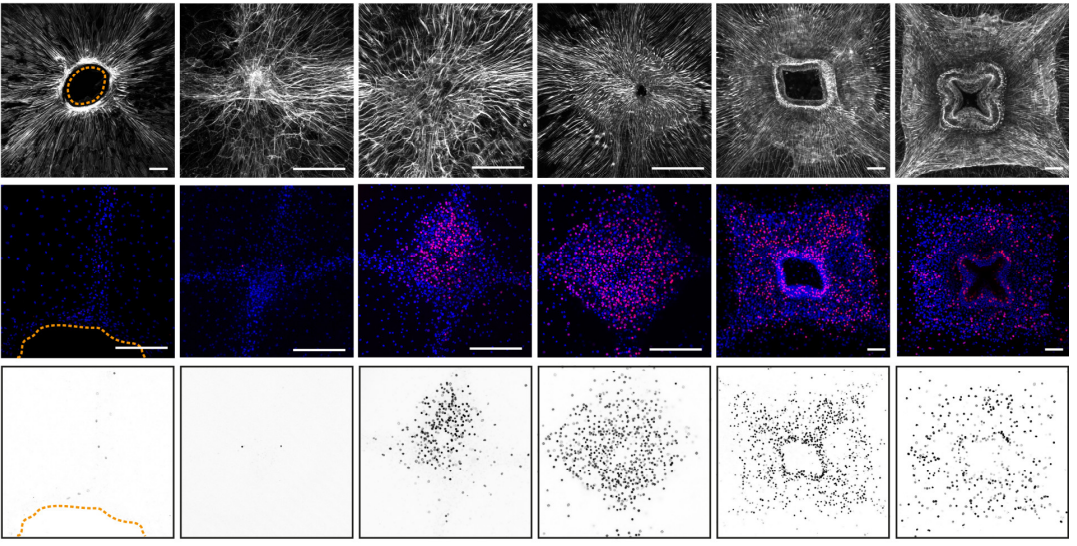
Pattern regulation in a regenerating jellyfish
Chiara Sinigaglia, Sophie Peron, Julia Steger, Evelyn Houliston, Lucas Leclère
Active Notch Signaling is Required for Arm Regeneration in a Brittle Star
Vladimir Mashanov, Jennifer Akiona, Maleana Khoury, Jacob Ferrier, Robert Reid, Denis Jacob Machado, Olga Zueva, Daniel Janies
Integrin-alpha-6+ Stem Cells (ISCs) are responsible for whole body regeneration in an invertebrate chordate
Susannah H. Kassmer, Adam Langenbacher, Anthony W. De Tomaso
Glucose metabolism promotes neonatal heart regeneration
Viviana M Fajardo Martinez, Iris Feng, Bao Ying Chen, Cesar A Perez, Baochen Shi, Peter Clark, Rong Tian, Ching-Ling Lien, Matteo Pellegrini, Heather Christofk, Haruko Nakano, Atsushi Nakano
Synergic coordination of stem cells is required to induce a regenerative response in anthozoan cnidarians
Aldine R. Amiel, Kevin Foucher, Solène Ferreira, Eric Röttinger
Citrullination regulates wound responses and tissue regeneration in zebrafish
Netta Golenberg, Jayne M. Squirrell, David A. Bennin, Julie Rindy, Paige E. Pistono, Kevin W. Eliceiri, Miriam A. Shelef, Junsu Kang, Anna Huttenlocher
Skip segment Hirschsprung disease: modelling the trans-mesenteric origin of the enteric nervous system in the human colon
Donald F Newgreen, James M Osborne, Dongcheng Zhang
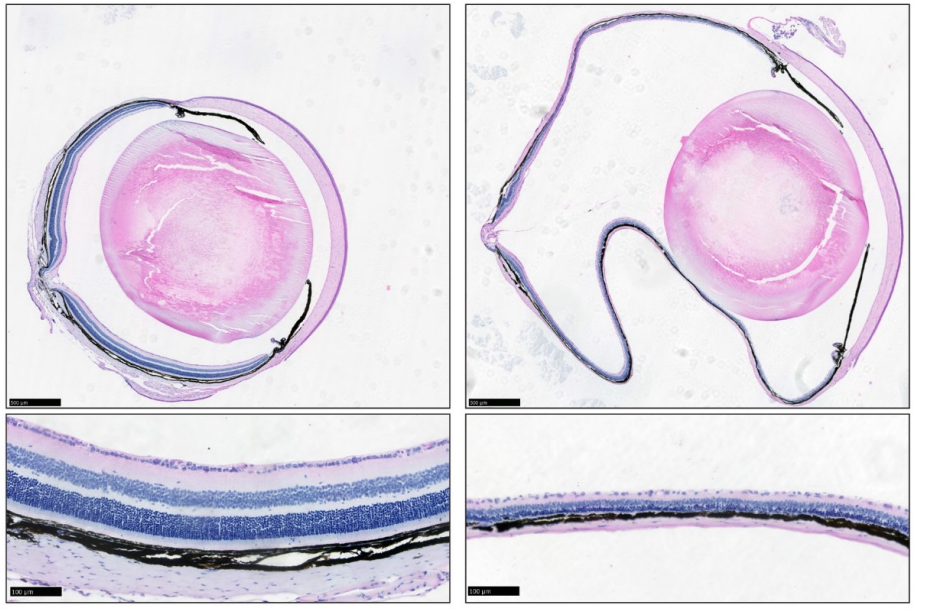
The nanophthalmos protein TMEM98 inhibits MYRF self-cleavage and is required for eye size specification
Sally H. Cross, Lisa Mckie, Toby W. Hurd, Sam Riley, Jimi Wills, Alun R. Barnard, Fiona Young, Robert E. MacLaren, Ian J. Jackson
A Genetic Screen Links the Disease-Associated Nab2 RNA-Binding Protein to the Planar Cell Polarity Pathway in Drosophila melanogaster
Wei-Hsuan Lee, Edwin Corgiat, J. Christopher Rounds, Zenyth Shepherd, Anita H. Corbett, Kenneth H. Moberg
Loss of the Reissner Fiber and increased URP neuropeptide signaling underlie scoliosis in a zebrafish ciliopathy mutant
Christine Vesque, Isabelle Anselme, Guillaume Pezeron, Yasmine Cantaut-Belarif, Alexis Eschstruth, Morgane Djebar, Diego López Santos, Hélène Le Ribeuz, Arnim Jenett, Hanane Khoury, Joëlle Véziers, Caroline Parmentier, Sylvie Schneider-Maunoury
A translational kidney organoid system bolsters human relevance of clinical development candidate
Amy Duyen Westerling-Bui, Thomas W. Soare, Srininivasan Venkatachalan, Michael DeRan, Eva Maria Fast, Alyssa B. Fanelli, Sergii Kyrychenko, Hien Hoang, Grinal M. Corriea, Wei Zhang, Maolin Yu, Matthew Daniels, Goran Malojcic, Xin-Ru Pan-Zhou, Mark W. Ledeboer, Jean-Christophe Harmange, Maheswarareddy Emani, Thomas T. Tibbitts, John F. Reilly, Peter Mundel
Characterization of human iPSC-derived astrocytes with potential for disease modeling and drug discovery
Vincent Soubannier, Gilles Maussion, Mathilde Chaineau, Veronika Sigutova, Guy Rouleau, Thomas Durcan, Stefano Stifani
mtor Haploinsufficiency Ameliorates Renal Cyst Formation in Adult Zebrafish tmem67 Mutants
Ping Zhu, Qi Qiu, Peter C. Harris, Xiaolei Xu, Xueying Lin
Characterization of SETD1A haploinsufficiency in humans and Drosophila defines a novel neurodevelopmental syndrome
Joost Kummeling, Diante E Stremmelaar, Nicholas Raun, Margot RF Reijnders, Marjolein H Willemsen, Martina Ruiterkamp-Versteeg, Marga Schepens, Calvin CO Man, Christian Gilissen, Megan T Cho, Kirsty McWalter, Margje Sinnema, James W Wheless, Marleen EH Simon, Casie A Genetti, Alicia M Casey, Paulien A Terhal, Jasper J van der Smagt, Koen L van Gassen, Pascal Joset, Angela Bahr, Katharina Steindl, Anita Rauch, Elmar Keller, Annick Raas-Rothschild, David A Koolen, Pankaj B Agrawal, Trevor L Hoffman, Nina N Powell-Hamilton, Isabelle Thiffault, Kendra Engleman, Dihong Zhou, Olaf Bodamer, Julia Hoefele, Korbinian M Riedhammer, Eva MC Schwaibold, Velibor Tasic, Dirk Schubert, Deniz Top, Rolph Pfundt, Martin R Higgs, Jamie M Kramer, Tjitske Kleefstra

Cleft lip and cleft palate (CL/P) in Esrp1 KO mice is associated with alterations in Wnt signaling and epithelial-mesenchymal crosstalk
SungKyoung Lee, Matthew J. Sears, Zijun Zhang, Hong Li, Imad Salhab, Philippe Krebs, Yi Xing, Hyun-Duck Nah, Trevor Williams, Russ P. Carstens
NCBP2 modulates neurodevelopmental defects of the 3q29 deletion in Drosophila and X. laevis models
Mayanglambam Dhruba Singh, Matthew Jensen, Micaela Lasser, Emily Huber, Tanzeen Yusuff, Lucilla Pizzo, Brian Lifschutz, Inshya Desai, Alexis Kubina, Sneha Yennawar, Sydney Kim, Janani Iyer, Diego E. Rincon-Limas, Laura Anne Lowery, Santhosh Girirajan
Transcriptomic changes due to early, chronic alcohol exposure during cortical development implicate regionalization, cell-type specification, synaptogenesis and WNT signaling as primary determinants of fetal alcohol Spectrum Disorders
Máté Fischer, Praveen Chander, Huining Kang, Jason P. Weick
| Plant development
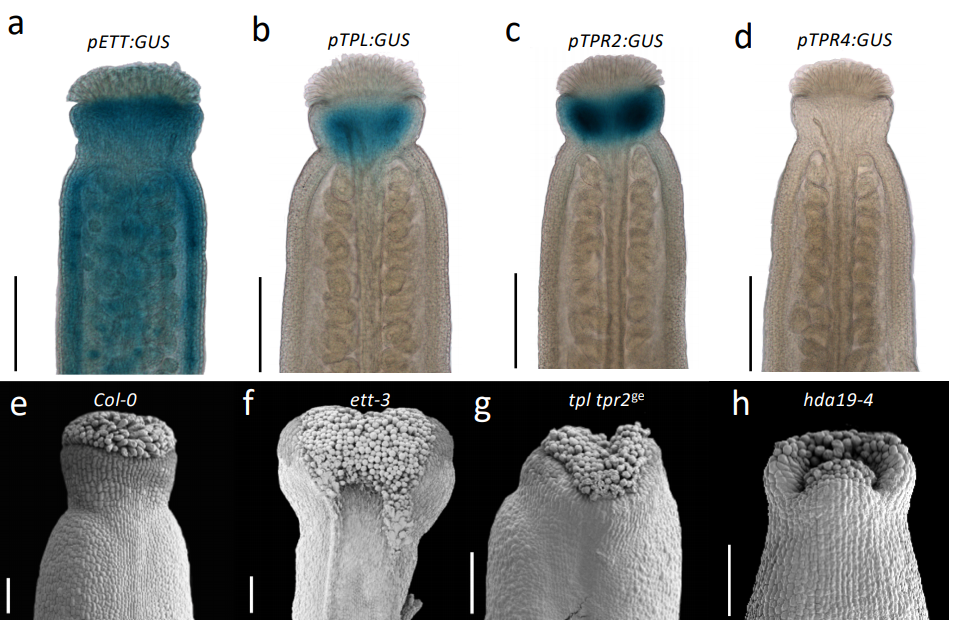
Direct ETTIN-auxin interaction controls chromatin state in gynoecium development
André Kuhn, Sigurd Ramans Harborough, Heather M. McLaughlin, Stefan Kepinski, Lars Østergaard
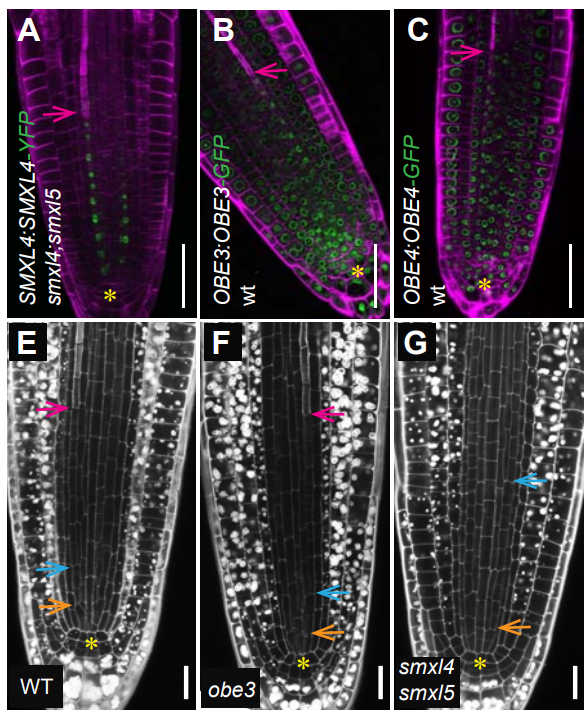
OBERON3 and SUPPRESSOR OF MAX2 1-LIKE proteins form a regulatory module specifying phloem identity
Eva-Sophie Wallner, Nina Tonn, Friederike Wanke, Vadir Lopéz-Salmerón, Michael Gebert, Christian Wenzl, Jan U. Lohmann, Klaus Harter, Thomas Greb
Genetic analysis of the Arabidopsis TIR1/AFB auxin receptors reveals both overlapping and specialized functions
Michael J. Prigge, Matthieu Platre, Nikita Kadakia, Yi Zhang, Kathleen Greenham, Whitnie Szutu, Bipin K. Pandey, Rahul Bhosale, Malcolm J. Bennett, Wolfgang Busch, Mark Estelle
Abundant expression of maternal siRNAs is a conserved feature of seed development
Jeffrey W. Grover, Diane Burgess, Timmy Kendall, Abdul Baten, Suresh Pokhrel, Graham J. King, Blake C. Meyers, Michael Freeling, Rebecca A. Mosher
Vein Patterning by Tissue-Specific Auxin Transport
Priyanka Govindaraju, Carla Verna, Tongbo Zhu, Enrico Scarpella
Impact of small RNAs in retrograde signalling pathways in Arabidopsis thaliana
Kristin Habermann, Bhavika Tiwari, Maria Krantz, Stephan O. Adler, Edda Klipp, M. Asif Arif, Wolfgang Frank
Arabidopsis ROOT PHOTOTROPISM2 is a Light-Dependent Dynamic Modulator of Phototropin1
Taro Kimura, Tomoko Tsuchida-Mayama, Hirotatsu Imai, Koji Okajima, Kosuke Ito, Tatsuya Sakai
Arabidopsis XTH4 and XTH9 contribute to wood cell expansion and secondary wall formation
Sunita Kushwah, Alicja Banasiak, Nobuyuki Nishikubo, Marta Derba-Maceluch, Mateusz Majda, Satoshi Endo, Vikash Kumar, Leonardo Gomez, Andras Gorzsas, Simon McQueen-Mason, Janet Braam, Björn Sundberg, Ewa J. Mellerowicz
ROOT PATTERNING AND REGENERATION ARE MEDIATED BY THE QUIESCENT CENTER AND INVOLVE BLUEJAY, JACKDAW AND SCARECROW REGULATION OF VASCULATURE FACTORS
Alvaro Sanchez-Corrionero, Pablo Perez-Garcia, Javier Cabrera, Javier Silva-Navas, Juan Perianez-Rodriguez, Inmaculada Gude, Juan Carlos del Pozo, Miguel A. Moreno-Risueno
Boron deficiency-induced root growth inhibition is mediated by brassinosteroid signalling regulation in Arabidopsis
Cheng Zhang, Mingliang He, Sheliang Wang, Liuyang Chu, Chuang Wang, Ningmei Yang, Guangda Ding, Hongmei Cai, Lei Shi, Fangsen Xu
Live-cell imaging of early events following pollen perception in self-incompatible Arabidopsis thaliana
Frédérique Rozier, Lucie Riglet, Chie Kodera, Vincent Bayle, Eléonore Durand, Jonathan Schnabel, Thierry Gaude, Isabelle Fobis-Loisy
MicroRNA function transitions from regulating developmental genes to transposable elements during the maturation of pollen
Cecilia Oliver, Maria Luz Annacondia, Zhenxing Wang, R Keith Slotkin, Claudia Köhler, German Martinez
Cytokinin induction by the jasmonate-induced AP2/ERF115 represses adventitious rooting in Arabidopsis
Abdellah Lakehal, Asma Dob, Zahra Rahneshan, Ondřej Novák, Sacha Escamez, Sanaria Alallaq, Miroslav Strnad, Hannele Tuominen, Catherine Bellini
In vivo mRNA structure regulates miRNA cleavage in Arabidopsis
Minglei Yang, Hugh C. Woolfenden, Yueying Zhang, Xiaofeng Fang, Qi Liu, Maria Louisa Vigh, Jitender Cheema, Xiaofei Yang, Matthew Norris, Sha Yu, Alberto Carbonell, Peter Brodersen, Jiawei Wang, Yiliang Ding
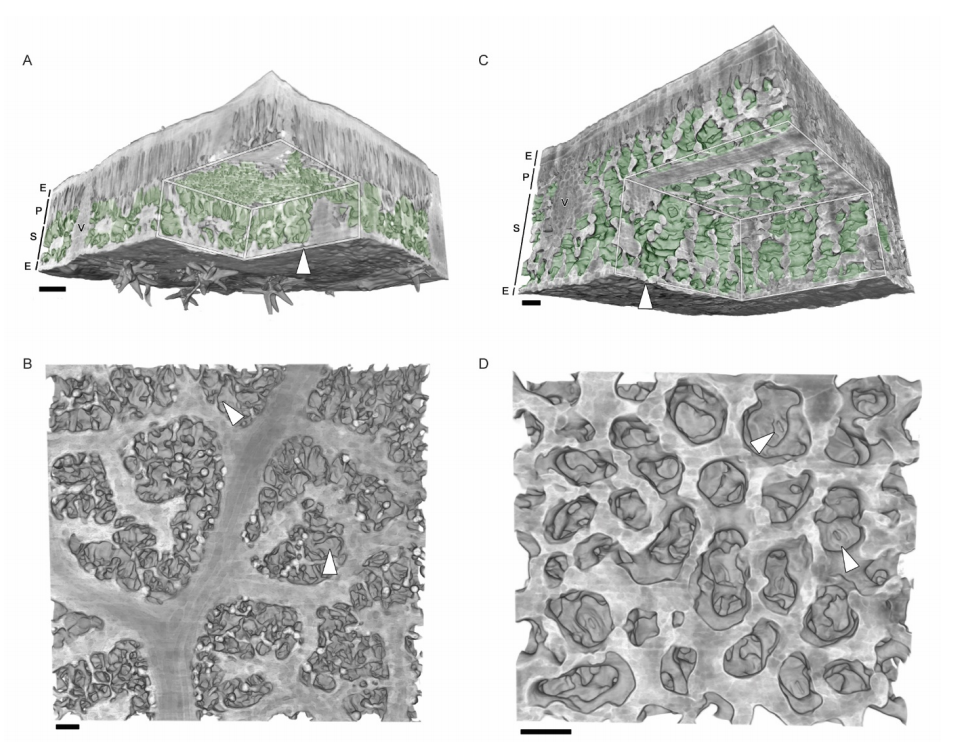
Emergent honeycomb topology of the leaf spongy mesophyll
Aleca M. Borsuk, Adam B. Roddy, Guillaume Théroux-Rancourt, Craig R. Brodersen
Auxin transport network underlies xylem bridge formation between the hemi-parasitic plant Phtheirospermum japonicum and host Arabidopsis
Takanori Wakatake, Satoko Yoshida, Ken Shirasu
Functional role of Polymerase IV during pollen development in Capsella
Zhenxing Wang, Nicolas Butel, Juan Santos-González, Filipe Borges, Jun Yi, Robert A Martienssen, German Martinez, Claudia Köhler
Compensatory guaiacyl lignin biosynthesis at the expense of syringyl lignin in 4CL1-knockout poplar
Chung-Jui Tsai, Peng Xu, Liang-Jiao Xue, Hao Hu, Batbayar Nyamdari, Radnaa Naran, Xiaohong Zhou, Geert Goeminne, Ruili Gao, Erica Gjersing, Joseph Dahlen, Sivakumar Pattathil, Michael G. Hahn, Mark F. Davis, John Ralph, Wout Boerjan, Scott A. Harding
The regulatory landscape of early maize inflorescence development
Rajiv K. Parvathaneni, Edoardo Bertolini, Md Shamimuzzaman, Daniel Vera, Pei-Yau Lung, Brian R. Rice, Patrick J. Brown, Alexander E. Lipka, Hank W. Bass, Andrea L. Eveland
LYS3 encodes a prolamin-box-binding transcription factor that controls embryo growth in barley and wheat
Beata Orman-Ligeza, Philippa Borrill, Tansy Chia, Marcella Chirico, Jaroslav Doležel, Sinead Drea, Miroslava Karafiátová, Nicole Schatlowski, Charles U. Solomon, Burkhard Steuernagel, Brande B. H. Wulff, Cristobal Uauy, Kay Trafford
Gene expression data support the hypothesis that Isoetes rootlets are true roots and not modified leaves
Alexander J. Hetherington, David M. Emms, Steven Kelly, Liam Dolan
Gene regulatory networks associated with lateral root and nodule development in soybean
Shuchi Smita, Jason Kiehne, Sajag Adhikari, Erliang Zeng, Qin Ma, Senthil Subramanian
Evo-devo & evo
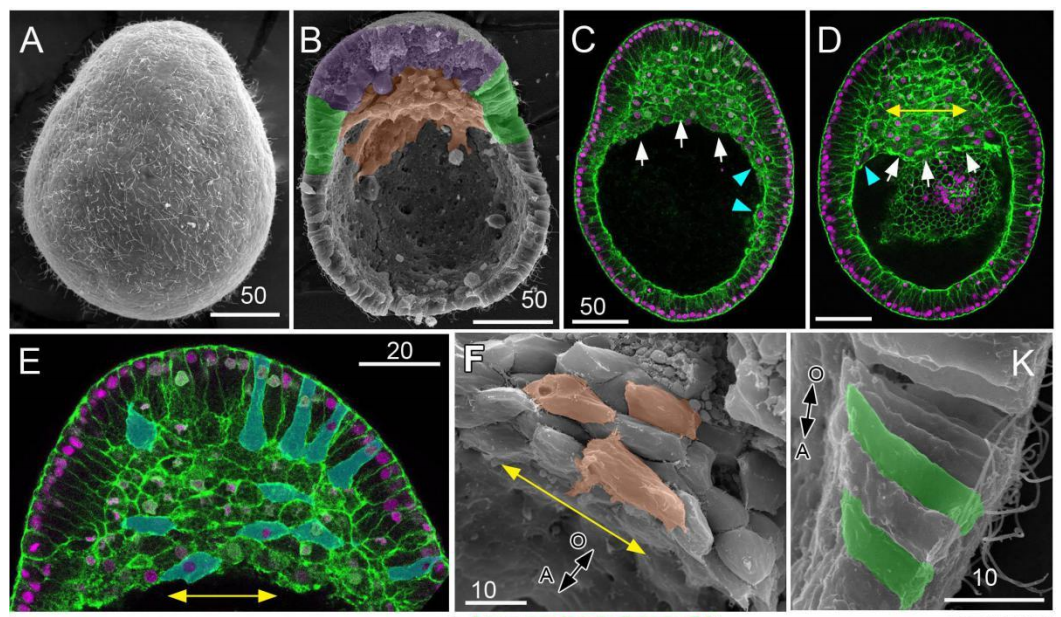
Cell shape changes during larval body plan development in Clytia hemisphaerica
Yulia Kraus, Sandra Chevalier, Evelyn Houliston
Evidence of multifaceted functions of codon usage in translation within the model beetle Tribolium castaneum
Carrie A. Whittle, Arpita Kulkarni, Cassandra G. Extavour

Genomic adaptations to aquatic and aerial life in mayflies and the origin of wings in insects
Isabel Almudi, Joel Vizueta, Alex de Mendoza, Chris Wyatt, Ferdinand Marletaz, Panos Firbas, Roberto Feuda, Giulio Masiero, Patricia Medina, Ana Alcaina, Fernando Cruz, Jessica Gómez-Garrido, Marta Gut, Tyler S. Alioto, Carlos Vargas-Chavez, Kristofer Davie, Bernhard Misof, Josefa González, Stein Aerts, Ryan Lister, Jordi Paps, Julio Rozas, Alejandro Sánchez-Gracia, Manuel Irimia, Ignacio Maeso, Fernando Casares
Co-evolving wing spots and mating displays are genetically separable traits in Drosophila
Jonathan H. Massey, Gavin R. Rice, Anggun Firdaus, Chi-Yang Chen, Shu-Dan Yeh, David L. Stern, Patricia J. Wittkopp
Dscam homophilic specificity is generated by high order cis-multimers coupled with trans self-binding of variable Ig1 in Chelicerata
Fengyan Zhou, Guozheng Cao, Songjun Dai, Guo Li, Hao Li, Zhu Ding, Shouqing Hou, Bingbing Xu, Wendong You, Feng Shi, Xiaofeng Yang, Yongfeng Jin
H3K27me3 natural variation selectively marks genes predicted to be important for differentiation in unicellular algae
Xue Zhao, Achal Rastogi, Anne Flore Deton Cabanillas, Ouardia Ait Mohamed, Catherine Cantrel, Berangère Lombard, Omer Murik, Auguste Genovesio, Chris Bowler, Daniel Bouyer, Damarys Loew, Xin Lin, Alaguraj Veluchamy, Fabio Rocha Jimenez Vieira, Leila Tirichine
Two distinct bacterial biofilm components trigger metamorphosis in the colonial hydrozoan Hydractinia echinata
Huijuan Guo, Maja Rischer, Martin Westermann, Christine Beemelmanns
Cell biology
The nucleus acts as a ruler tailoring cell responses to spatial constraints
A.J. Lomakin, C.J. Cattin, D. Cuvelier, Z. Alraies, M. Molina, G. Nader, N. Srivastava, J.M. Garcia-Arcos, I.Y. Zhitnyak, A. Bhargava, M.K. Driscoll, E.S. Welf, R. Fiolka, R.J. Petrie, N. Manel, A.M. Lennon-Duménil, D.J. Müller, M. Piel
Emergence of single cell mechanical behavior and polarity within epithelial monolayers drives collective cell migration
Shreyansh Jain, Victoire M.L. Cachoux, Gautham H.N.S. Narayana, Simon de Beco, Joseph D’Alessandro, Victor Cellerin, Tianchi Chen, Mélina L. Heuzé, Philippe Marcq, René-Marc Mège, Alexandre J. Kabla, Chwee Teck Lim, Benoit Ladoux
ERK-mediated mechanochemical waves direct collective cell polarization
Naoya Hino, Leone Rossetti, Ariadna Marin-Llaurado, Kazuhiro Aoki, Xavier Trepat, Michiyuki Matsuda, Tsuyoshi Hirashima
Excess Centrosomes Disrupt Vascular Lumenization and Endothelial Cell Adherens Junctions
Danielle M Berlin, Erich J Kushner, Katy L Davis, Victoria L Bautch
Tyrosine-based Signals Regulate the assembly of Daple•PARD3 Complex at Cell-cell Junctions during Polarized Planar Cell Migration
Jason Ear, Anokhi Saklecha, Navin Rajapakse, Julie Choi, Majid Ghassemian, Irina Kufareva, Pradipta Ghosh
Tuba activates Cdc42 during neuronal polarization downstream of the small GTPase Rab8a
Pamela J. Urrutia, Felipe Bodaleo, Daniel A. Bórquez, Victoria Rozes-Salvador, Cristopher Villablanca, Cecilia Conde, Mitsunori Fukuda, Christian González-Billault
Zygote morphogenesis but not the establishment of cell polarity in Plasmodium berghei is controlled by the small GTPase, RAB11A
H. Patil, K.R. Hughes, L. Lemgruber, N. Philip, N. Dickens, A. P. Waters
Alpha synuclein aggresomes inhibit ciliogenesis and multiple functions of the centrosome
Anila Iqbal, Marta Baldrighi, Jennifer N. Murdoch, Angeleen Fleming, Christopher J. Wilkinson
Ubiquitin Links Smoothened to Intraflagellar Transport to Regulate Hedgehog Signaling
Paurav B. Desai, Michael W. Stuck, Bo Lv, Gregory J. Pazour
Aurora B and C kinases regulate prophase exit and chromosome segregation during spermatogenesis
Stephen R. Wellard, Karen Schindler, Philip Jordan
Akt1-associated actomyosin remodelling is required for nuclear lamina dispersal and nuclear shrinkage in epidermal terminal differentiation
Clare Rogerson, Duncan Wotherspoon, Ryan F L O’Shaughnessy
Modelling
Perturbation analysis of a multi-morphogen Turing Reaction-Diffusion stripe patterning system reveals key regulatory interactions
Andrew D. Economou, Nicholas A.M. Monk, Jeremy B.A. Green
A mathematical model for cell polarization in zebrafish primordial germ cells
Carolin Dirks, Paul Striewski, Benedikt Wirth, Anne Aalto, Adan Olguin-Olguin, Erez Raz
Large-Scale Survey of Cell-Differentiation Programs in a Generative Model Reveals Regeneration as an Epiphenomenon of Development
Somya Mani, Tsvi Tlusty
Homeorhesis in Waddington’s Landscape by Epigenetic Feedback Regulation
Yuuki Matsushita, Kunihiko Kaneko
Multiscale mechanical model for cell division orientation in developing biological systems
B. Leggio, J. Laussu, E. Faure, P. Lemaire, C. Godin
Collective cell migration and residual stress accumulation: modeling consideration
I. Pajic-Lijakovic, M. Milivojevic
Mechanical cell competition in heterogeneous epithelial tissues
R. J. Murphy, P. R. Buenzli, R. E. Baker, M. J. Simpson
A Stochastic Model for Actin Waves in Eukaryotic Cells
Jifeng Hu, Varunyu Khamviwath, Hans G. Othmer
Tools & resources
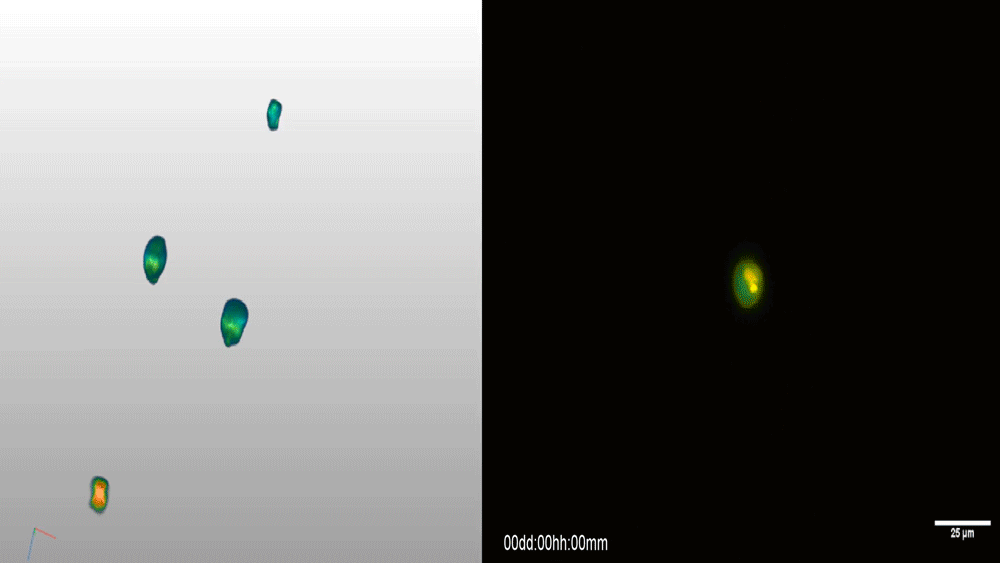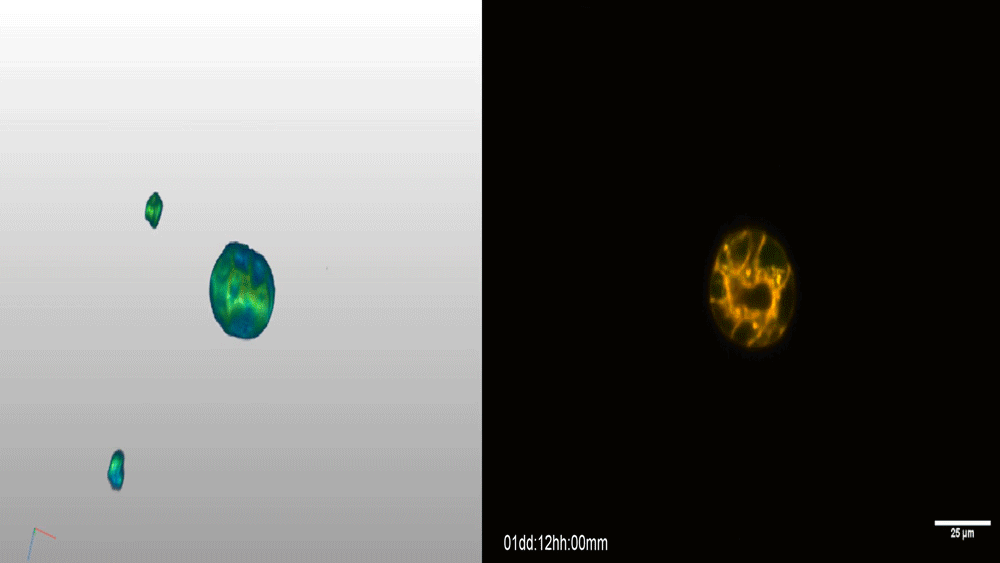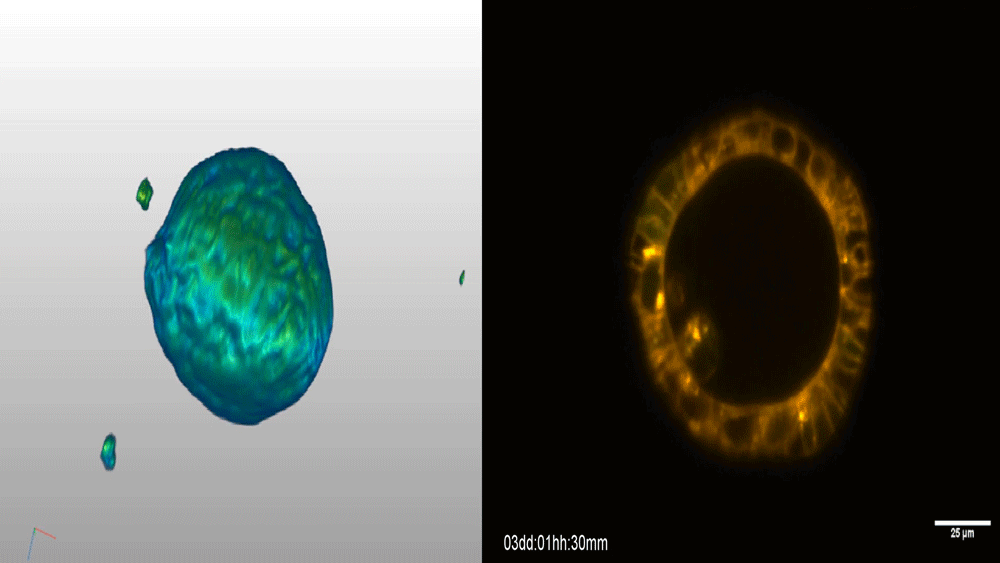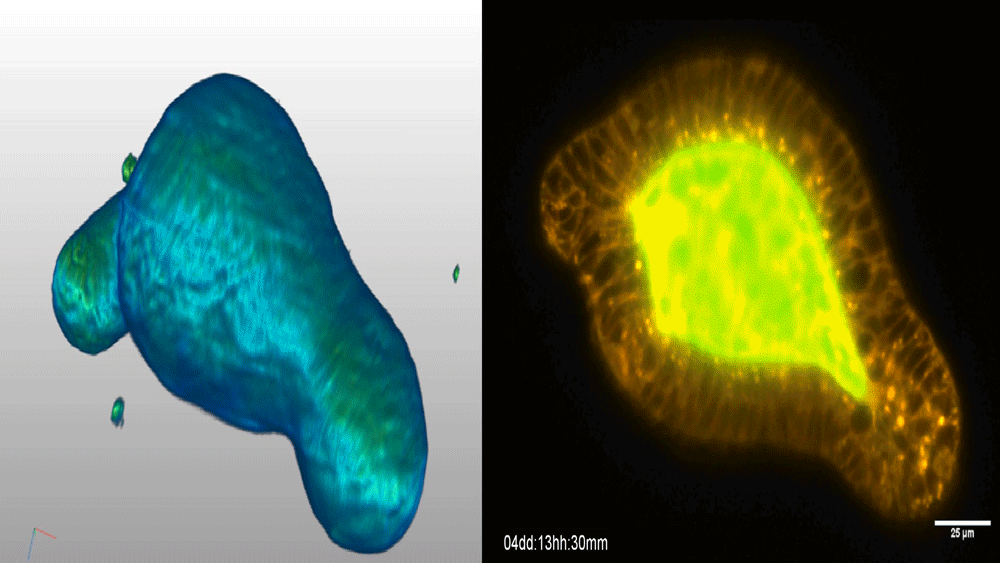
Visualizing the metazoan proliferation-differentiation decision in vivo
Abraham Q. Kohrman, Rebecca C. Adikes, Jayson J. Smith, Michael A. Q. Martinez, Taylor N. Medwig-Kinney, Nicholas J. Palmisano, Maria D. Sallee, Ononnah B. Ahmed, Nicholas Weeks, Nuri Kim, Simeiyun Liu, Wan Zhang, Ariel M. Pani, David Q. Matus
Computationally Enhanced Quantitative Phase Microscopy Reveals Autonomous Oscillations in Mammalian Cell Growth
Xili Liu, Seungeun Oh, Leonid Peshkin, Marc W. Kirschner
Multi-photon attenuation-compensated light-sheet fluorescence microscopy
Madhu Veettikazhy, Jonathan Nylk, Federico Gasparoli, Adrià Escobet-Montalbán, Anders Kragh Hansen, Dominik Marti, Peter Eskil Andersen, Kishan Dholakia
Optimization and functionalization of red-shifted rhodamine dyes
Jonathan B. Grimm, Ariana N. Tkachuk, Heejun Choi, Boaz Mohar, Natalie Falco, Ronak Patel, Jennifer Lippincott-Schwartz, Timothy A. Brown, Luke D. Lavis
SMALL ALPHAHERPESVIRUS LATENCY-ASSOCIATED PROMOTERS DRIVE EFFICIENT AND LONG-TERM TRANSGENE EXPRESSION IN THE CENTRAL NERVOUS SYSTEM
Carola J. Maturana, Jessica L. Verpeut, Thomas J. Pisano, Zahra M. Dhanerawala, Andrew Esteves, Lynn W. Enquist, Esteban A. Engel
Rapid and Inexpensive Preparation of Genome-Wide Nucleosome Footprints from Model and Non-Model Organisms
Laura E McKnight, Johnathan G Crandall, Thomas B Bailey, Orion GB Banks, Kona N Orlandi, Vi N Truong, Grace L Waddell, Elizabeth T Wiles, Drake A Donovan, Scott D Hansen, Eric U Selker, Jeffrey N McKnight
A large-scale resource for tissue-specific CRISPR mutagenesis in Drosophila
Fillip Port, Claudia Strein, Mona Stricker, Benedikt Rauscher, Florian Heigwer, Jun Zhou, Celine Beyersdörffer, Jana Frei, Amy Hess, Katharina Kern, Roberta Malamud, Bojana Pavlovic, Kristin Rädecke, Lukas Schmitt, Lukas Voos, Erica Valentini, Michael Boutros
Principles for rational Cas13d guide design
Hans-Hermann Wessels, Alejandro Méndez-Mancilla, Xinyi Guo, Mateusz Legut, Zharko Daniloski, Neville E. Sanjana
Non-destructive enzymatic deamination enables single molecule long read sequencing for the determination of 5-methylcytosine and 5-hydroxymethylcytosine at single base resolution
Zhiyi Sun, Romualdas Vaisvila, Bo Yan, Chloe Baum, Lana Saleh, Mala Samaranayake, Shengxi Guan, Nan Dai, Ivan R. Corrêa Jr., Sriharsa Pradhan, Theodore B. Davis, Thomas C. Evans Jr., Laurence M. Ettwiller
Extensive Mammalian Germline Genome Engineering
Yanan Yue, Yinan Kan, Weihong Xu, Hong-Ye Zhao, Yixuan Zhou, Xiaobin Song, Jiajia Wu, Juan Xiong, Dharmendra Goswami, Meng Yang, Lydia Lamriben, Mengyuan Xu, Qi Zhang, Yu Luo, Jianxiong Guo, Shengyi Mao, Deling Jiao, Tien Dat Nguyen, Zhuo Li, Jacob V. Layer, Malin Li, Violette Paragas, Michele E. Youd, Zhongquan Sun, Yuan Ding, Weilin Wang, Hongwei Dou, Lingling Song, Xueqiong Wang, Lei Le, Xin Fang, Haydy George, Ranjith Anand, Shi Yun Wang, William F. Westlin, Marc Güell, James Markmann, Wenning Qin, Yangbin Gao, Hong-jiang Wei, George M. Church, Luhan Yang
Research practice & education
Grant reviewer perceptions of the quality and effectiveness of panel discussion
Stephen A. Gallo, Karen B. Schmaling, Lisa A. Thompson, Scott R. Glisson
Science communication in online media: influence of press releases on coverage of genetics and CRISPR
Rafał Grochala
An illustration of reproducibility in neuroscience research in the absence of selective reporting
Xiang-Zhen Kong, ENIGMA Laterality Working Group, Clyde Francks
Women and Men Were Proportionally Represented Among Speakers at Major National Neurology Conferences in 2017
Mollie McDermott, James F. Burke, Haley McCalpin, Anita V. Shelgikar, Douglas J. Gelb, Abbey Dunn, Nicholas J. Beimer, Zachary N. London
Implicit bias is strongest when assessing top candidates
Emma R Andersson, Carolina Hagberg, Sara Hägg
Is the replication crisis a problem for biologists? A geometric morphometric approach
Juan Vrdoljak, Kevin Imanol Sanchez, Roberto Arreola-Ramos, Emilce Guadalupe Diaz Huesa, Alejandro Villagra, Luciano Javier Avila, Mariana Morando
Insights from a survey-based analysis of the academic job market
Jason D. Fernandes, Sarvenaz Sarabipour, Christopher T. Smith, Natalie M. Niemi, Nafisa M. Jadavji, Ariangela J. Kozik, Alex S. Holehouse, Vikas Pejaver, Orsolya Symmons, Alexandre W. Bisson Filho, Amanda Haage
Efficient data management infrastructure for the integration of imaging and omics data in life science research
Luis Kuhn Cuellar, Andreas Friedrich, Gisela Gabernet, Luis de la Garza, Sven Fillinger, Adrian Seyboldt, Sven zur Oven-Krockhaus, Friederike Wanke, Sandra Richter, Wolfgang M. Thaiss, Marius Horger, Nisar Malek, Klaus Harter, Michael Bitzer, Sven Nahnsen
Tracking self-citations in academic publishing
Ameni Kacem, Justin W. Flatt, Philipp Mayr
 (No Ratings Yet)
(No Ratings Yet)
 (1 votes)
(1 votes)



 Genetics Unzipped is delighted to host a short series of podcasts recorded at the 2019 Galton Institute symposium – New Light on Old Britons – which took place at the Royal Society in London at the end of October. Reporter Georgia Mills talks to some of the leading researchers uncovering the hidden stories of the people of the British Isles.
Genetics Unzipped is delighted to host a short series of podcasts recorded at the 2019 Galton Institute symposium – New Light on Old Britons – which took place at the Royal Society in London at the end of October. Reporter Georgia Mills talks to some of the leading researchers uncovering the hidden stories of the people of the British Isles.






