In this episode from our series exploring 100 ideas in genetics, we’re taking a trip to London with William Bateson and discovering that the famous story about him reading Mendel’s paper on the train might not be all that it seems.
Plus, we seek the secrets of snapdragons, and learn how to build an army of MinIONs.
If you enjoy the show, please do rate and review and spread the word. And you can always send feedback and suggestions for future episodes and guests to podcast@geneticsunzipped.com
Vascular development critically involves pruning, which helps to remodel an immature network containing excess microvessels into a mature and functioning one. The mechanisms of vascular remodelling and the relationship between the endothelial cells and the other cell types with which they are closely associated are, however, currently poorly understood. A new Development paper now demonstrates a crucial role for oxygen sensing by astrocytes in vascular remodelling of the mouse retina. We caught up with the two-author team behind the paper: research associate Li-Juan Duan and her supervisor Guo-Hua Fong, Professor of Cell Biology at the University of Connecticut Health Center, to hear more about the story.
Li-Juan (L) and Guo-Hua (R)
Guo-Hua, can you give us your scientific biography and the questions your lab is trying to answer?
GHF After graduating from Hangzhou University in 1982, I attended the PhD Program in Biochemistry at the University of Illinois at Urbana-Champaign. My interest in angiogenesis began during my postdoctoral training with Martin Breitman and Janet Rossant in Toronto. After reporting the finding that Flt-1 (VEGFR1) was a negative regulator of vasculogenesis, I set up my first lab at the University of Western Ontario before relocating to where I am now (UConn School of Medicine).
Early in my career as a developmental biologist, I used mouse embryos to study vasculogenesis and angiogenesis, but since about a decade ago I have been primarily focused on mouse retinas. At the molecular level, my current focus is on oxygen sensing and hypoxia signalling because growth of blood vessels is mostly (although not exclusively) an adaptive response to tissue hypoxia. Molecules of interest include hypoxia inducible factor (HIF) 2α (also called EPAS1), a transcription factor stabilized by hypoxia that activates the expression of angiogenic factors, and prolyl hydroxylase domain (PHD) proteins (EGLNs), which sense oxygen and destabilize HIFα proteins by oxygen-dependent prolyl hydroxylation reactions.
Li-Juan: how did you come to join the Fong lab and what drives your research?
LJD With a few exceptions, almost all tissues are vascularized. Mechanisms ensuring that blood vessels are formed at the right place, right time and with right configurations are very fascinating to me. Dr Fong’s lab investigates the molecular and cellular mechanisms regulating blood vessel growth, and has contributed significantly to this rapidly advancing field. So the opportunity to be a part of team and contribute to the exciting discoveries is a major driving force for my research. Also, the potential that knowledge generated from these studies might improve angiogenesis therapies is quite energizing.
What was known about the link between astrocytes, endothelial cells and oxygen in retinal angiogenesis prior to your work?
LJD & GHF The retinal astrocytic network is situated just beneath the vascular network on the inner retinal surface, with foot processes from astrocytes wrapping around vascular endothelial cells (ECs) and supporting their survival. It was thought that the astrocytic network forms before retinal angiogenesis and serves as a template for the subsequent vascular growth. The newly formed vascular network exhibits a honeycomb pattern consisting of almost uniformly sized capillaries, separated by roundish non-vascular tissues. To gain circulatory functions, the nascent vascular plexus undergoes active remodelling to form a tree-like pattern with large trunks and smaller branches. During this process, excess microvessels are pruned away. One widely held but never formally documented view is that oxygen causes pruning by activating EC apoptosis. The other theory is that T cells interact with ECs to cause pruning. In both theories, astrocytes were not suspected to play a role in remodelling.
Can you give us the key results of the paper in a paragraph?
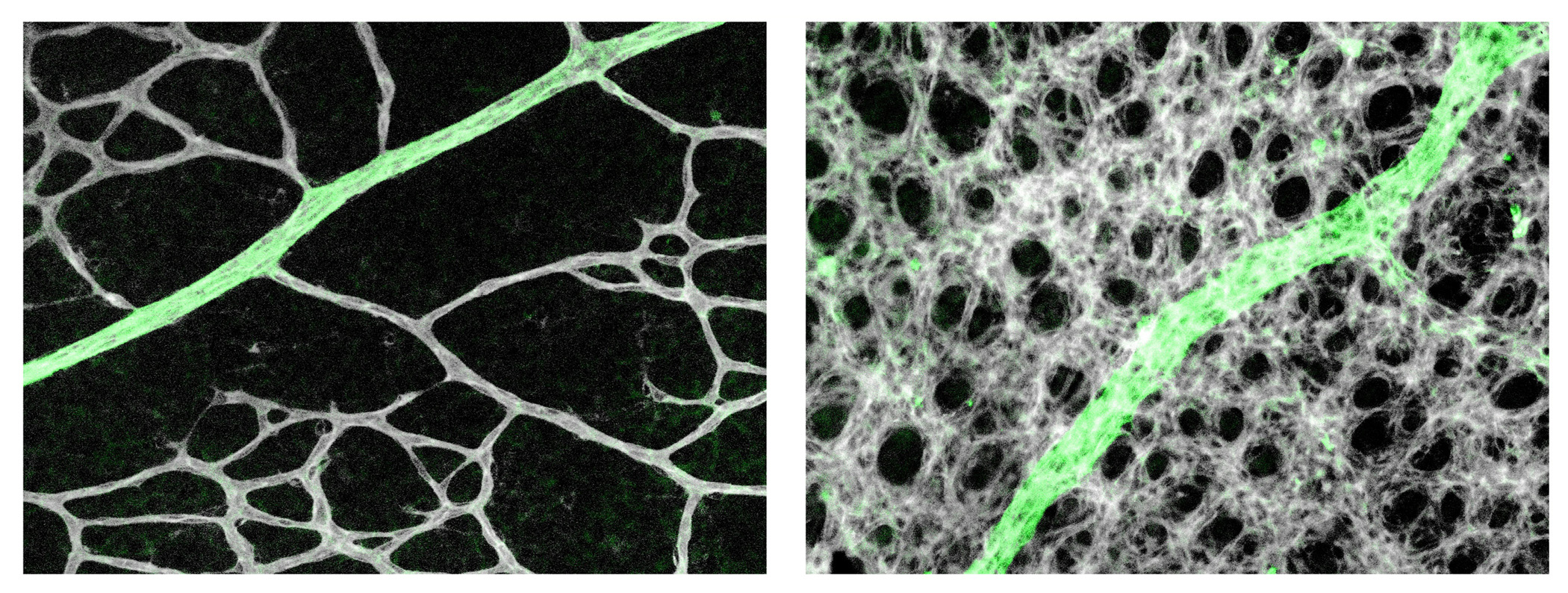
LJD & GHF We found that retinal astrocytes respond to changes in tissue oxygenation mostly through prolyl hydroxylase domain (PHD) protein 2, an enzyme known to use O2 and oxoglutarate as substrates to hydroxylate transcription factors HIF1α and HIF2α, causing them to degrade. In neonatal mouse retinas, HIF2α is essential for the expansion of the astrocyte population owing to its role in maintaining astrocytes in immature and proliferative states. Angiogenesis imposes a physiological limit to the abundance of retinal astrocytes by causing O2-dependent HIF2α degradation. As such, a proportion of the numerous capillaries generated through robust angiogenesis is unable to acquire astrocytic support and undergoes regression (pruning). In support of this theory, targeted disruption of the Phd2 gene in astrocytes led to accumulation of HIF-2α protein, expansion of retinal astrocyte population, and persistence of extra microvessels. When astrocyte growth was directly stimulated by intraocular injection of PDGFA into the eyes in wild-type mice, vascular pruning was also blocked. Based on these findings, we conclude that oxygen- and PHD2-dependent astrocyte growth arrest is a feedback mechanism to prevent runaway vascularization.
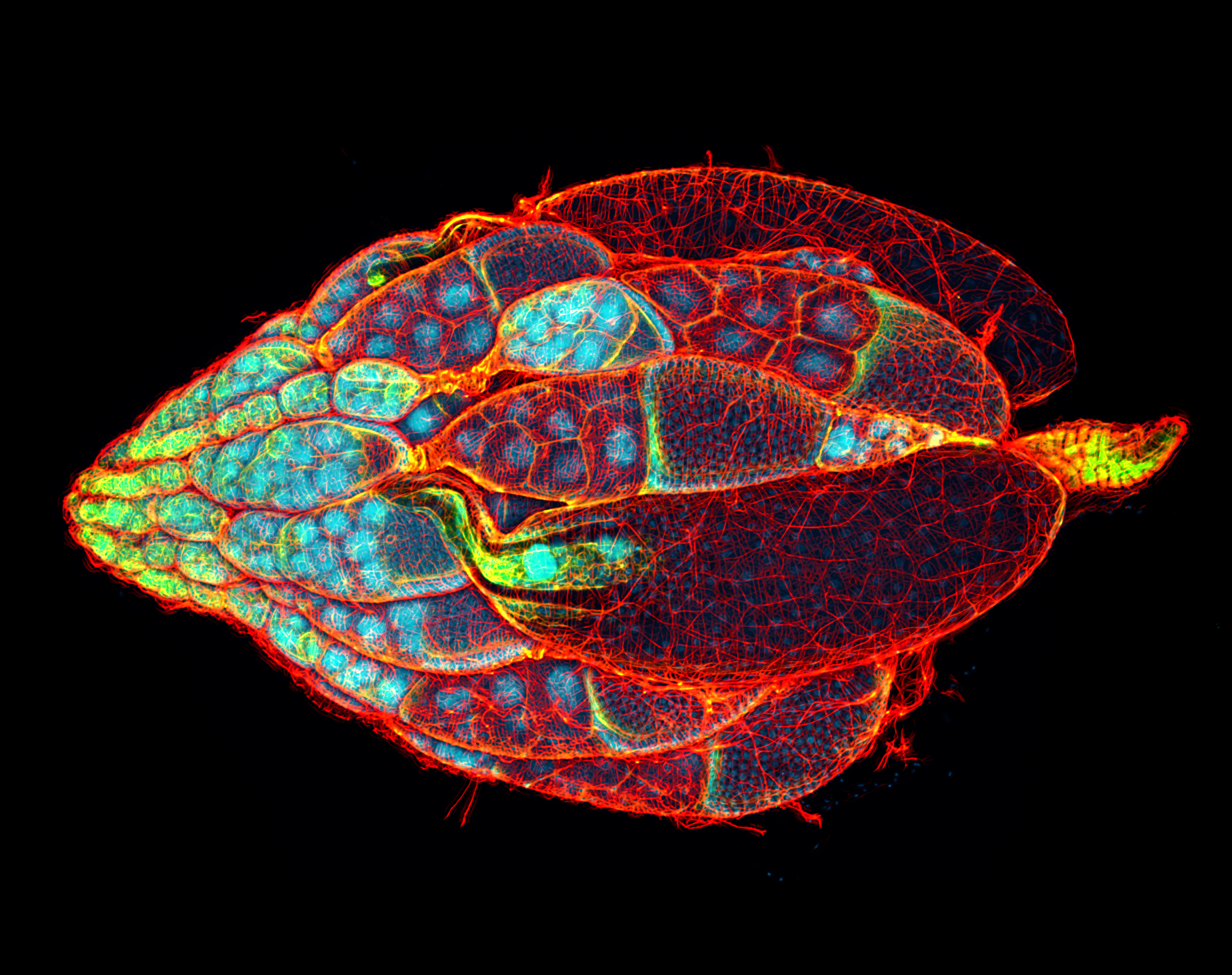
Capillaries are pruned off near arterial branches (green) in wild-type mice (left) but persist in mice deficient for PHD2 in astrocytes (right).
Do you have any ideas about what happens downstream of Phd2/HIFα to induce differentiation in astrocytes?
LJD & GHF We don’t really know yet, but there may be some candidates. HIF2α is known to promote the expression of Oct4, a transcription factor important for the maintenance of stem cell identity. While the astrocyte precursors may not be exactly stem cells, they might share certain features related to stem cells. So it seems to us that HIF2α-dependent expression of Oct4 might be a potential mechanism by which HIF2α helps maintain astrocytes at precursor and immature states. At this point, this idea remains highly speculative but it could be a reasonable starting point for the next step.
Does your work have any relevance to pathological contexts?
LJD & GHF Capillary dropout is a common feature in early stage diabetic retinopathy. The prevailing view at present is that capillary dropout starts with apoptosis of pericytes, a type of supporting cell that adheres to the endothelial cells. Our finding of a crucial role for astrocytes in capillary maintenance raises the question of whether the adverse effects of diabetes on retinal astrocytes might also contribute to capillary dropout? If so, the retinal astrocytes could be a novel therapeutic target for improving retinal vascular stability. In this regard, it may be interesting to note that there’s a publication indicating that retinal astrocyte abundance is decreased in diabetic rats (Ly et al., 2011).
Our findings are also related to retinopathy of prematurity (ROP), a disease associated with premature birth. Whereas pathological mechanisms underlying ROP are complicated and go beyond oxygen alone, sensitivity to oxygen is nonetheless an important component. Thus, targeting astrocytic PHD2 could provide a novel opportunity for minimizing the consequence of ROP.
When doing the research, did you have any particular result or eureka moment that has stuck with you?
LJD I guess the moment was when I found that simply injecting PDGFA into the eyes caused both expanded astrocyte populations and persistence of more microvessels, whereas injection of PBS into the contralateral eyes didn’t. These results indicated that increased astrocyte abundance alone was sufficient to block vascular pruning.
And what about the flipside: any moments of frustration or despair?
LJD Sure, plenty of them. This was actually a lengthy project. While I’ve been working on other projects in parallel, this project was on and off for nearly 6 years. While we observed the overcrowded vascularization at the very start of the project, it was rather challenging trying to explain why. Naturally, our initial attention was on VEGF, and lots of time was wasted in that direction. We couldn’t just publish overcrowded capillaries without being able to explain the underlying mechanism, so the project was really stuck for a long time until one day we realized perhaps it was simply the number of astrocytes itself rather than any specific molecule. We also ran into a lot of technical difficulties. One example was growing primary retinal astrocytes: because they could not be expanded too much, we had to pool the cells from a very large number of retinas. These are just two of the many examples.
Where will this work take the Fong lab?
GHF We will be pursuing several directions. We are interested in identifying the potential targets downstream of PHD2/HIF2α by RNA-seq, and then assessing their functionality in retinal vascular development by CRISPR/Cas9-mediated knockout. We will be also looking into potential interaction between HIF2α with a transcription factor called Nr2e1 or Tlx. We are interested in finding out whether the two transcription factors collaborate in retinal astrocytes in controlling astrocyte differentiation. We are also interested in investigating if PHD2 deficiency in astrocytes also confers protection to capillaries in disease models such as oxygen-induced retinopathy and diabetic retinopathy.
For whatever reasons, the vascular biology community has paid very little attention to astrocytes. The major focus is still on endothelial cells, and their interaction with pericytes, vascular smooth muscle cells and leukocytes. In fact, we also stumbled into this subject accidentally, but we increasingly appreciated the opportunity that fell upon us as we began to realize the importance of this cell type to vascular growth, stability and function. Because so little is known about astrocytes in the context of vascular biology, there are plenty of exciting findings to be made. In the coming years, we will be devoting much of our efforts trying to understand how astrocytes communicate with endothelial cells to regulate various aspects of vascular homeostasis.
Because so little is known about astrocytes in the context of vascular biology, there are plenty of exciting findings to be made
Finally, let’s move outside the lab – what do you like to do in your spare time in Connecticut?
LJD Spring is beautiful in Connecticut. I enjoy growing flowers, watching them blossom in the morning. There are also plenty of water reservoirs with hiking trails open to the public. I enjoy taking long walks around the lakes on weekends, especially in the spring and fall.
GHF Connecticut may not be a frequently talked-about place but it is not exactly in the middle of nowhere either – in fact the UConn School of Medicine campus is just about 2 h from Boston and New York, respectively. So it’s relatively convenient to spend a day in the weekend visiting museums or friends there, or even shopping. Also Connecticut is full of very long hiking trails at different levels of difficulty, so one could spend hours (or days if they like) hiking to fairly distant locations, with some leading all the way into neighbouring states.
The Poché Lab is seeking a highly motivated postdoctoral research associate/fellow with experience in mouse retinal developmental biology, regeneration, and transcriptome analysis. This position is focused on the study of the molecular mechanisms blocking mammalian Müller glial cell (MG)-mediated retinal regeneration. Our long-term goal is to determine whether the mouse retina retains latent regenerative potential, akin to other vertebrates species, and whether we can genetically “awaken” that potential to restore sight.
Special emphasis will be placed on the investigation of MG transcriptional reprogramming to a progenitor-like state. Preference will be given to candidates with a strong background in mouse genetics and in techniques to probe the retinal transcriptome and epigenome. This expertise should ideally include next gen sequencing (RNA-seq, ChIP-seq, ATAC-seq, single cell-seq, etc.) data analysis.
The Poché lab employs a multi-disciplinary approach utilizing genetic loss- and gain-of-function experiments, fate mapping, gene therapy, molecular biology, and live retinal confocal microscopy. We are housed in the Department of Molecular Physiology and Biophysics at Baylor College of Medicine (BCM). Located in the Texas Medical Center, the largest medical center on the world, BCM postdocs have a tremendous amount of technical and intellectual resources at their disposal.
In your application, please include a cover letter, current CV, and contact information for three references. Application review will begin immediately and will continue until the position is filled. Please contact Dr. Poché at poche@bcm.edu.
Two BBSRC-funded post-doctoral positions will become available as part of a collaboration between the Universities of St Andrews and Aberdeen:
evolutionary developmental biologist (three years, currently proposed start date 1 January 2020, mostly based in St Andrews)
molecular developmental biologist/RNA molecular biologist (three years, currently proposed start date 1 January 2020, mostly based in Aberdeen)
The collaboration will study evolution of Wnt signalling and alternative transcript expression, splicing and function.
A BHF-funded position will become available in Aberdeen:
theoretical biologist/bioinformatician/mathematician/physicist (three years, flexible start date, e.g. 1 August 2019)
This project will model and simulate Gene-Regulatory Networks controlling heart muscle (cardiomyocyte) cell differentiation as part of a BHF Programme Grant in collaboration with two laboratory developmental biologist already working with stem cells and embryos, respectively.
These positions have not yet been officially advertised. At this early stage, informal enquiries are encouraged to Dave Ferrier <dekf@st-andrews.ac.uk> and Stefan Hoppler <s.p.hoppler@abdn.ac.uk>. Official announcements with more details will follow as soon as possible.
With over 10,000 votes cast, almost 6,000 people viewing the galleries and a new record for daily page views on the Node, we can now announce the winners of our inaugural calendar competition. We were blown away by the quality of the entries – 62 images of all kinds of cells, tissues and embryos. Check out the original postto see all the entries – as you’ll see, so many beautiful images missed out, and we’d like to thank everyone who took part.
So, category by category, here are the 12 winners who will make the final print calendar, and below them a full vote rundown (there were quite a few close calls!). We’re aiming to take the calendars with us to two upcoming meetings: SDB in Boston in July and the European Developmental Biology Congress in Alicante in October. Come grab one if you’re going.
Mammals
1st place: Light up
By Paul Gerald Layague Sanchez
(EMBL Heidelberg)
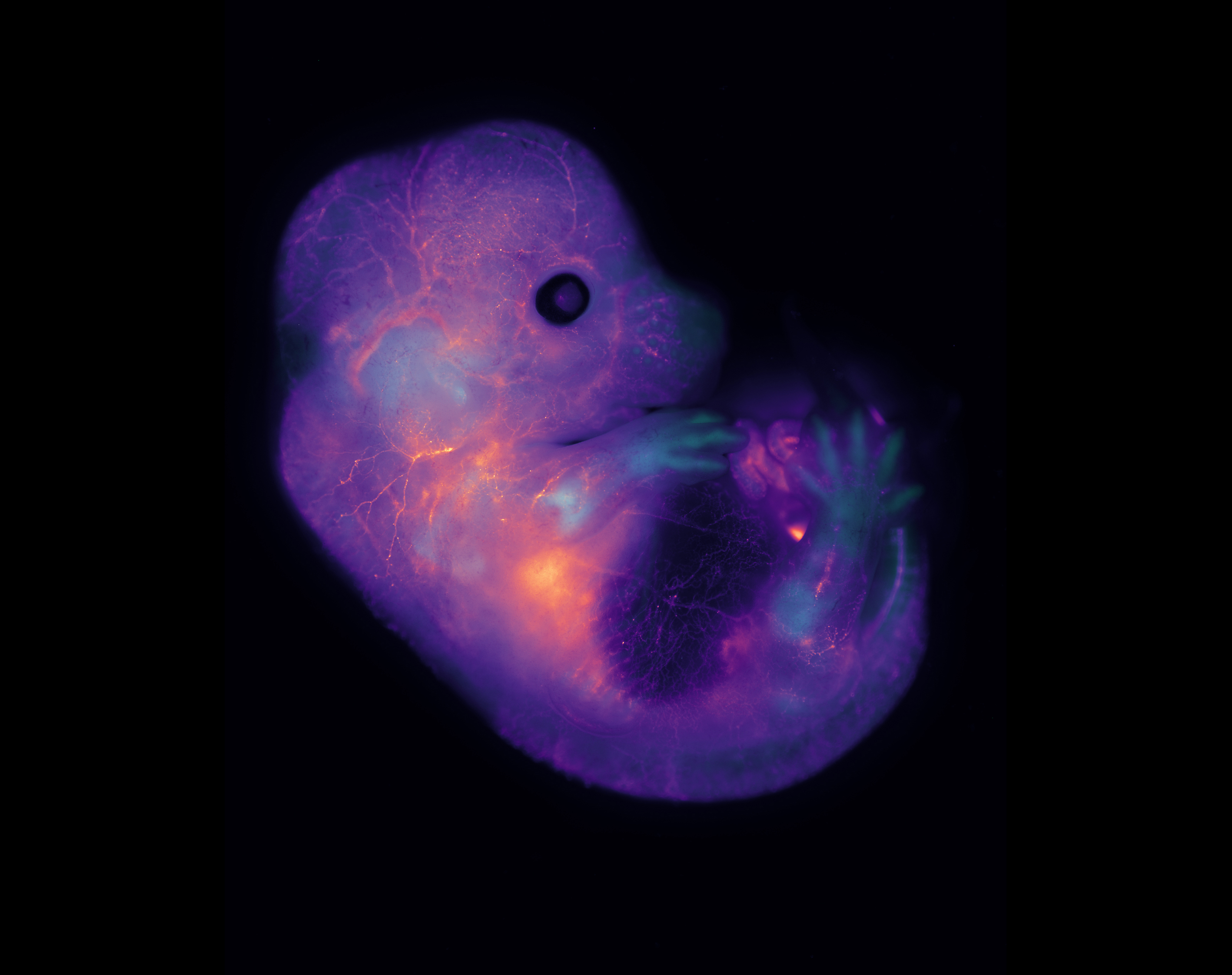
E14.5 mouse embryo labeled for cartilage (Sox9-GFP, in biop-SpringGreen) and vasculature (highlighter ink circulated by injection in a blood vessel, in mpl-magma). Vasculature “lights up” the embryo, including within the developing bones of the limbs. Image taken using a microscope kindly sponsored by Zeiss during the 2018 Embryology Course at the Marine Biological Laboratory in Woods Hole, MA.
2nd place: Human neuron
By Nicholas Gatford
(Institute of Psychiatry, Psychology and Neuroscience, King’s College London)
Super-resolution image acquired via structured illumination microscopy of a single developing human neuron ectopically expressing the autism risk gene neuroligin-4X with enlarged growth cones. F-actin is in green, HA-tagged neuroligin-4X is in magenta, doublecortin is in cyan confirming its neuronal identity, and the nuclear marker DAPI is in grey.
Zebrafish
1st place: Zebrafish head
By Oscar Ruiz
(Department of Genetics MD Anderson Cancer Center)
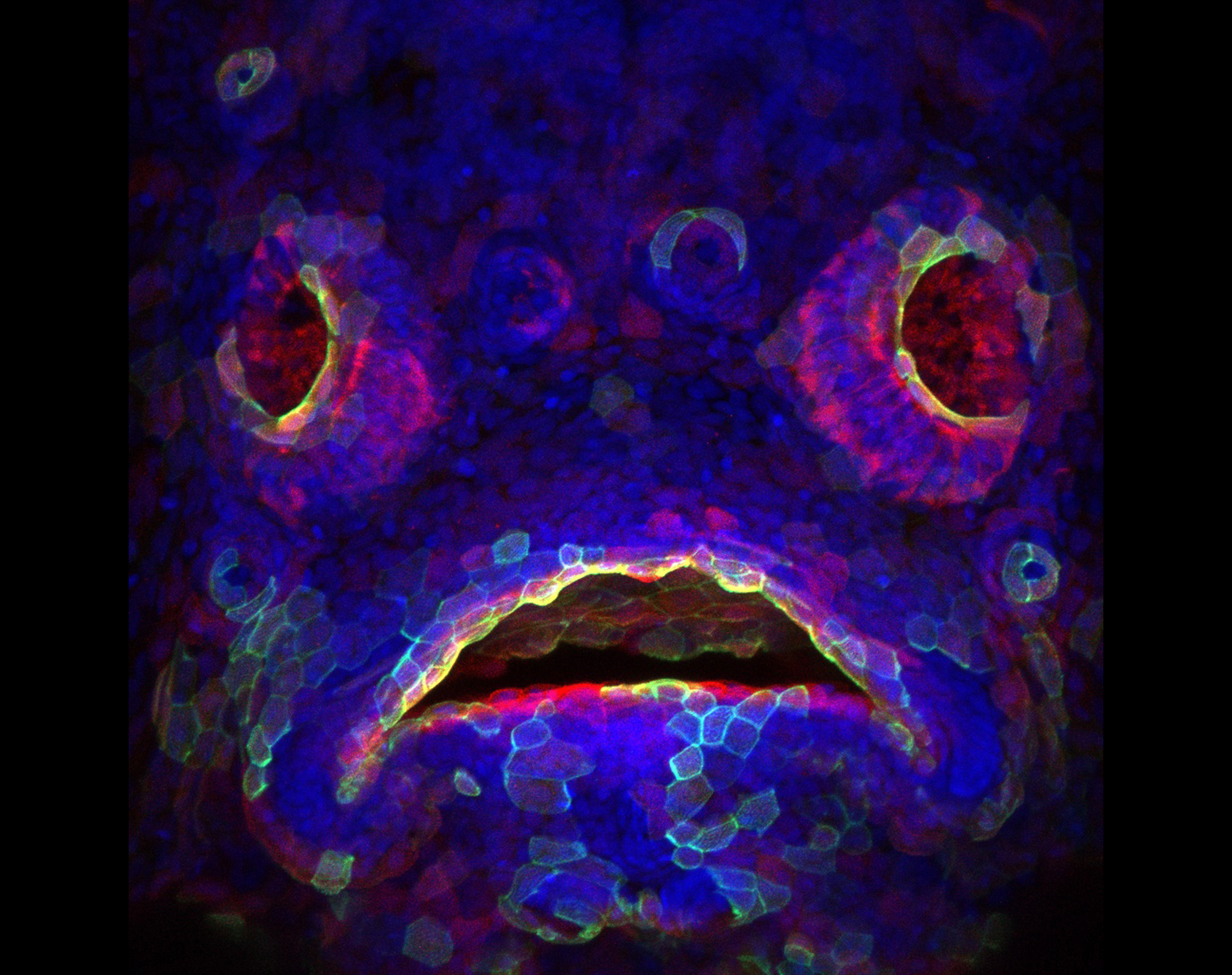
Transgenic zebrafish (Danio rerio) larva expressing red fluorescent protein in the developing mouth and olfactory epithelium. A subset of cells also express a construct that labels actin filament with green fluorescent protein. DAPI (blue) is used to label DNA in the nuclei of all cells.
2nd place: Zebrafish gills
By Philippa Carr
(Bateson Centre, University of Sheffield)
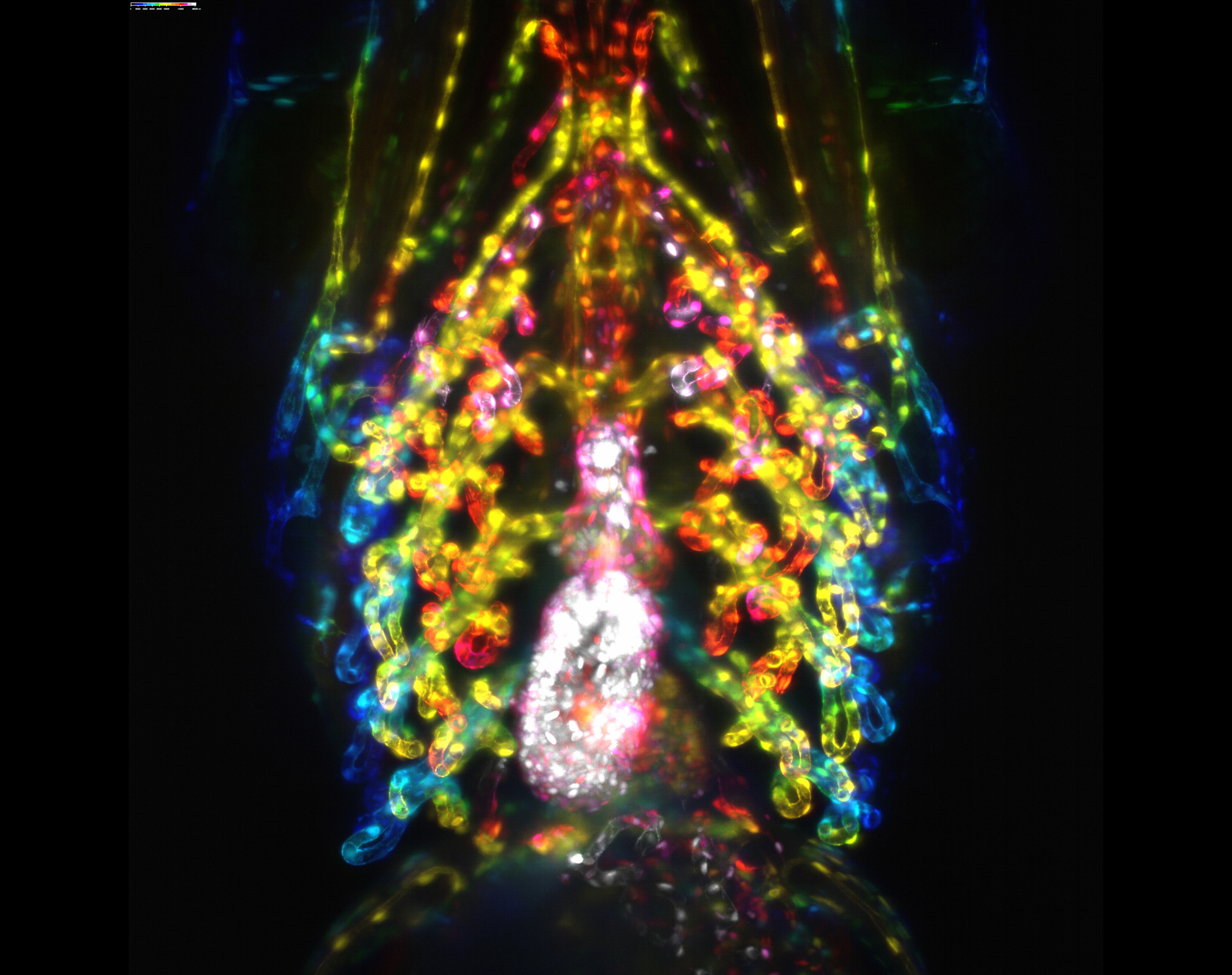
Developing gill vasculature in a 120h old zebrafish and also features the heart. This image was taken using lightsheet microscopy in two transgenic lines, one that marks the endothelial actin and the other marks the endothelial nuclei. After acquisition it was processed as a colour coded depth projection.
Vertebrate variety show
1st place: Alligator
By Daniel Smith Paredes
(Department of Geology and Geophysics, Yale University)
Alligator mississipiensis embryo at stage 13-14 immunostained against Myosin heavy chain showing the developing muscles and (red) and neurofilament labeling axons of nerves.
2nd place: Chicken embryo
By Laurel Yohe
(Department of Geology and Geophysics, Yale University)
Stage 35 chicken embryo, cleared and immunostained for DAPI (orange) and Pax3 (cyan) demonstrating the developing neural crest and spinal cord. Image was taken on the Nikon AZ-C2 macro-confocal with image analysis performed in Imaris. Image was taken in collaboration with Andrea Attardi at the Max Planck Institute of Molecular Cell Biology and Genetics during the Woods Hole 2018 Embryology course.
Drosophila
1st place: Drosophila ovary
By Yujun Chen
(Kansas State University Division of Biology).
*Yujun also wins the ‘Star of Instagram’ award for most-liked post (we posted all 62 individually from Development’s account!), and the image is the new profile pic*
Drosophila whole ovary stained for f-actin (Red), nuclei (Cyan) and actin (Green).
2nd place: Metallic flight
By Marisa Merino
(Department of Biochemistry, University of Geneva)
Drosophila mutant showing a decreased eye size compared to wild type. This line is not able to generate descendants with wild type flies.
Invertebrate variety show
1st place: Bobtail squid
By Martyna Lukoseviciute
(Weatherall Institute of Molecular Medicine, University of Oxford)
Live Hawaiian Bobtail Squid (Euprymna scolopes), stained with vital dyes (CellMask, LysoTracker and Hoechst) to understand its cellular and sub-cellular organisation during development. Blue is labelling cellular nuclei, green – cell plasma membranes and red – lysosomes that are important for cellular waste removal. This species is a candidate model organism that yet holds many answers to developmental biology questions, such as nervous system and eye development. The image was taken during the MBL 2018 Embryology Course with the confocal microscope provided by Zeiss. Animals were supplied by the cephalopod researcher Carrie Albertin.
2nd place: Hydractinia
By Indu Patwal
(Centre for Chromosome Biology, National University of Ireland Galway)
DIC and fluorescence image of Hydractinia male sexual (left) and feeding polyps (right) on a chitin bed. Chitin is shown in green. Noncycling cells probed with cyclin-dependent kinase inhibitor (CDKI) are shown in yellow, which are mainly in nematocytes, male gonophore, and gastrodermis.
Plants, Fungi and Choanoflagellates
1st place: Arabidopsis lateral root
By Robertas Ursache
(University of Lausanne, Switzerland)
The development of a lateral root in Arabidopsis thaliana. The sample has been cleared and stained with Calcofluor White to outline the cell walls and the green fluorescent nuclei represent a protein expressed specifically in the outer cell layer of developing lateral root.
Art and illustration
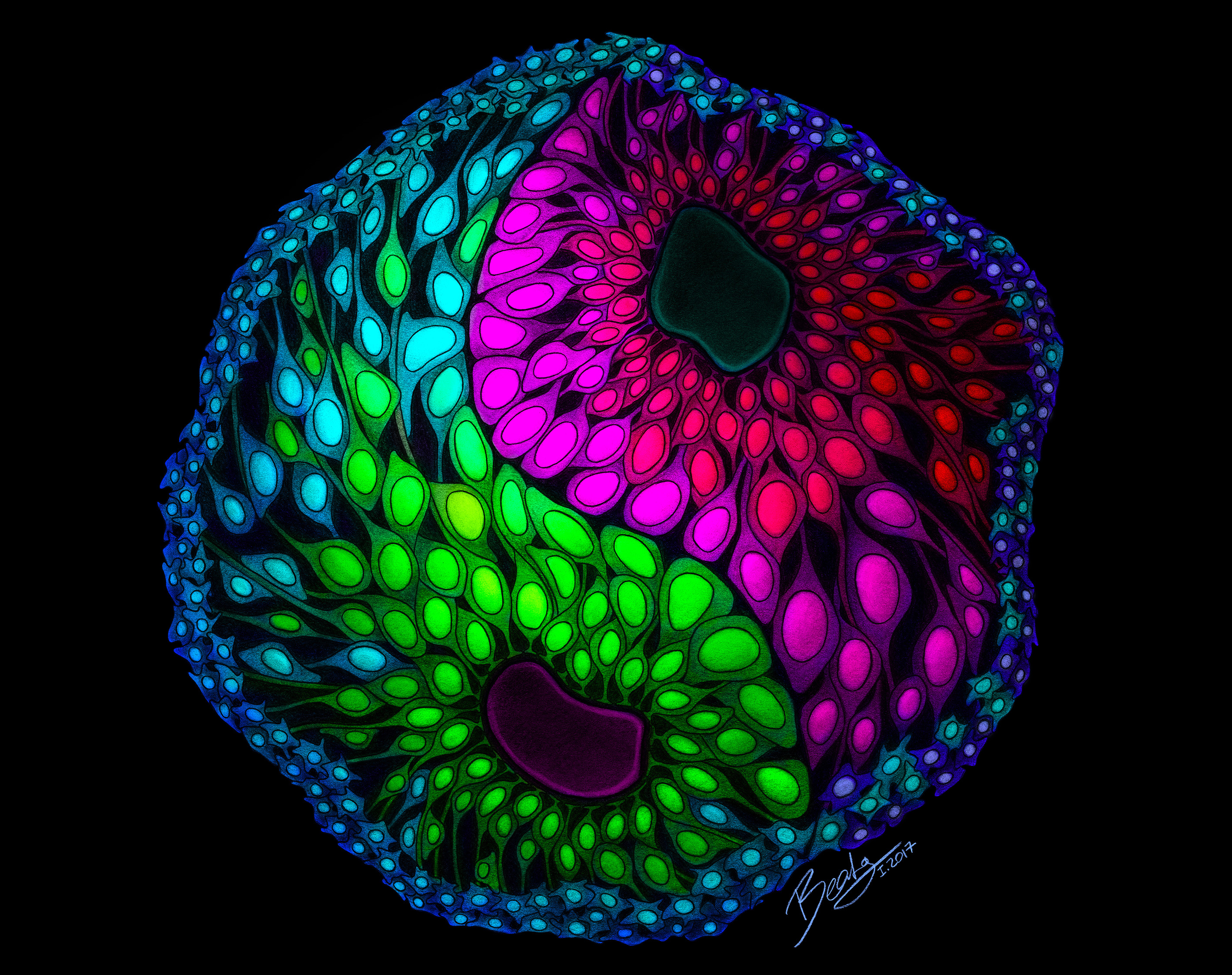
1st place: The yin and yang of developmental patterning
By Beata Edyta Mierzwa
(Ludwig Institute for Cancer Research and the University of California, San Diego, and www.beatascienceart.com)
This drawing illustrates the remarkable self-organization capacity of cerebral organoids that allows them to recapitulate human brain development in vitro. Each color represents a different type of cell, and the dorsal and ventral areas are separated by a defined boundary – like a yin and yang symbolizing the balance between distinct but complementary entities.
Brigham and Women’s Hospital, Development of the Musculo-Skeletal system / Pourquie Lab, Boston, MA, USA
Job Summary
The Research Lab Manager will assume a key role in the Pourquie lab (Brigham and Women’s Hospital, Pathology/Harvard Medical School/Genetics Departments) (15 to 18-person lab). The Lab manager works under the very general direction of Principal Investigator and participates in research involving up-to-date biological techniques in developmental biology and cell culture to investigate the mechanisms controlling the formation of the vertebrae and muscles during embryonic development. Under general direction, performs non-routine and highly specialized experimental procedures that involve substantial “hands on” work and the ability to multi-task across various projects within the lab. Has day-today supervisory responsibility of laboratory operations and personnel. Coordinates lab activities, composes and may present research reports, participates in design and modification of lab protocols and assist in grant data generation and writing.
Principal duties and responsibilities:
Works independently with general guidance from the Principal Investigator.
Independently performs routine biological experiments. Experimental techniques may include: protein biochemistry; cell fixation and basic immunohistochemistry/immunofluorescence; gene expression analysis such as quantitative real-time PCR, in-situ hybridization, molecular biology, standard mammalian cell culture techniques as well as isolation of primary stem cell cultures from animal tissues.
Independently performs non-routine, highly specialized experimental procedures including in vitro direct differentiation of pluripotent stem cells, maintenance of pluripotent cell lines, microsurgery and gene transfer technologies in chicken and mouse embryo model systems, advanced molecular cloning, in vivo cell transplantation in mouse models.
Performs confocal microscopy techniques and flow cytometry experiments, time-lapse imaging and live cell microscopy and analysis of resulting data using instrument software
Performs advanced data analysis using advanced statistical techniques.
Trains Lab staff in routine and specialized lab techniques
Oversees and coordinates scheduling of lab procedures
Oversees mouse colonies and coordinates the use of mice by other laboratory members
Oversees all lab supply and equipment, organization, purchasing and maintenance
Oversees activities related to laboratory compliance with institutional health and safety regulations, such as proper inventory, storage, use, and disposal of radioactive and toxic substances and serves as the designated laboratory Safety Officer.
Manages and supervises the day-to-day activities of the lab’s Technical Research Assistants (grades I or II)
Coordinates lab resources utilization and material requests with other labs
Qualifications:
Master of Science in a biological science required, PhD preferred
Minimum 3 years’ hands-on experience in cell culture
Previous experience as a supervisor or manager
Experience with laboratory mouse colony maintenance preferred
Experience with embryonic stem cell or iPS cell culture preferred
Skills / Abilities / Competencies required
Strong organizational and scientific skills
Sound interpersonal skills, ability to constructively interact with the research team members
Excellent oral and written communication skills
High degree of computer literacy
Working conditions
Standard biosciences laboratory environment
Exposure to laboratory reagents, chemicals, and animal /human tissues under controlled conditions. Minimal risk when following established protocols and federal, state, local, and hospital guidelines
To apply to this position please send your resume and cover letter to Olivier Pourquie (pourquie@genetics.med.harvard.edu)
An NIH-funded postdoctoral position is available in Jianbo Wang’s lab at the Univ. of Alabama at Birmingham. We study how shape forms during embryogenesis, and how aberrant shape formation leads to congenital birth defects. The process of shape formation is known as morphogenesis, and is regulated in part by the planar cell polarity (PCP) pathway. PCP is a unique signaling mechanism that coordinates cellular polarity and regulates dynamic, directional cell behavior. Our studies involve multi-disciplinary approaches including genetic, imaging, cell biology and biochemistry; and use multiple model organisms including the mouse, chick and Xenopus. Our goals are to 1) uncover novel mechanisms and logic of PCP signaling; 2) elucidate the role of PCP in cardiovascular, skeletal and neural development; and 3) investigate how human PCP gene variants contribute to congenital disorders.
We are looking for highly motivated applicants with training in Developmental Biology, Genetics, Cell Biology or a related discipline. To apply, please email your CV, a description of previous research and contact information for three references to j18wang@uab.edu.
This blog post is a reflection on the Company of Biologists-organized workshop on Chromatin-Based Regulation of Development that I recently attended at Wiston House, located in the countryside of Sussex in the UK.
Wiston House, Steyning, UK
To my mind, this workshop was the ideal of what a meeting should be. As a freshly minted assistant professor at the University of California, San Francisco, this workshop came at an excellent time for me to learn, glean inspiration, and extract advice.
A good conference is much more than the sum of its parts – the “parts” being the topics represented by the scientist attendees. By reading publications, you learn what experiments worked and hung together in a narrative. By hearing that work presented in person, you can infer the hunches or obsessions that drove the scientist to ask the question in the first place, as well as see the pieces of data that are forming the nucleus for the next phase of work.
A good conference provides an opportunity to rapidly survey ongoing work in your field. Through discussions and debates, you can learn whether your field has reached a consensus that a prevailing model consistently explains observations, or whether alternative explanations are being fomented. In this frame of mind, a new question that no one else seems to be asking might occur to you. Equally helpfully, you may discern whether a particular area is overpopulated and incorporate that knowledge to adjust your own project strategy. By assimilating all of this, you expand your knowledge base and learn by observing how other scientists think.
Mid-scientific debate in the library at Wiston House
At a good conference, you will forge connections with peers that may turn into friendships and collaborations in the future. You will interact with leaders in your field, gaining invaluable opportunity to pick their brains for career advice and to watch a well-honed scientific intuition at work. In doing all of this, you will have been doing the dreaded “networking”, too.
Taking a break for a hike “up the downs” (photo by @sudpinglay)
When it’s all over, you will return to your home lab armed with new ideas to test, with a new solution to a hurdle you have faced, or with new critiques from your peers to answer.
All of these features of the ideal conference were on display at this Company of Biologists workshop. The small size, intimate setting, and considerate pace of the meeting allowed for time to ruminate over exciting presentations and talk through ideas with other attendees. Early career researchers and established faculty were equal participants both in numbers and in the share of the conference program allotted to them. This was truly the most thoughtfully organized workshop I have ever attended.
Researchers from the Tokyo University of Science, Japan, have used CRISPR gene editing technology to make snails with shells that coil the ‘wrong’ way, providing insights into the fundamental basis of left-right asymmetry in animals. These findings were recently published in Development.
If you look at a snail’s shell, the chances are it will coil to the right. But, occasionally, you might find an unlucky one that twists in the opposite direction – as fans of Jeremy the lefty snail (https://en.wikipedia.org/wiki/Jeremy_(snail)) will remember, these snails struggle to mate with the more common rightward-coiling individuals.
This chirality (direction of coiling) of snail shells is an outward manifestation of left-right asymmetry: a phenomenon seen across animal evolution and extending to humans – your heart is (probably) on your left side, while your liver is to the right. But how does this asymmetry come about? Researchers from Japan, writing in the journal Development, think they now have a definitive answer – for one species of freshwater snail (Lymnaea stagnalis) at least.
Successfully applying CRISPR gene editing technology to molluscs for the first time, Masanori Abe and Reiko Kuroda (working at Tokyo University of Science, but recently relocated to Chubu University, Japan), have now made snails with mutations in a gene called Lsdia1, which had previously been suggested – but not conclusively proven – to be involved in snail shell coiling; snails without a functional copy of Lsdia1 produce offspring with shells that coil to the left, showing that this single gene is responsible for rightward- coiling. Surprisingly, the researchers could see signs of asymmetry at the earliest possible stage of development – when the snail embryo was just a single cell. Moreover, the mutant snails could be reared to adults, when they produced exclusively leftward-coiling offspring. According to Kuroda: “It is remarkable that these snails with reversed coiling are healthy and fertile, and that this coiling can be inherited generation after generation (we now have 5th-generation leftward-coiling snails). Further, these results may have an implication for snail evolution and speciation – given that left- and rightward-coiling snails probably wouldn’t interbreed.”
Knocking out one gene in the snail Lymnaea stagnalis reverses shell coiling. In contrast to the wild-type dextral snail (right), a CRISPR-created snail shows sinistral coiling (left). Credit: Dr Hiromi Takahashi of the Kuroda laboratory.
It’s still not clear how Lsdia1 might control left-right asymmetry: the gene encodes a formin, a protein that is involved in regulating the cell’s internal skeleton, but more work is needed to understand how this influences the cellular behaviours that control handedness – which is something Kuroda and her colleagues are actively working on. But given that genes like Lsdia1 are found throughout the animal kingdom, similar mechanisms for controlling left-right asymmetry could be at play in other species – including our own. As Kuroda says: “Although diverse mechanisms have been proposed for different animals, we think a unified mechanism, involving formins and cellular chirality, is probable”. So while it may seem a big leap from snail shell coiling to human left-right asymmetries, it’s possible that future studies on how Lsdia1 works in snails might eventually help us understand why some babies are born with their heart on the right (which is of course the wrong) side of their chest.
Our call for images to fill our 2019-20 calendar was met with an amazing response – 62 entries showcasing the diverse beauty of developmental biology. Now it’s time for you vote for the 12 that will make it into print.
Because we want a range of organisms and styles in the calendar, and because picking 12 favourites from 62 is not the easiest task, we’ve decided to split the voting up into categories. Turns out 12 into 62 doesn’t go so well, and some categories were better represented than others, so we ended up with the following:
Mammals (vote for 2 out of 10)
Zebrafish (vote for 2 out of 8)
Vertebrate variety show (vote for 2 out 10)
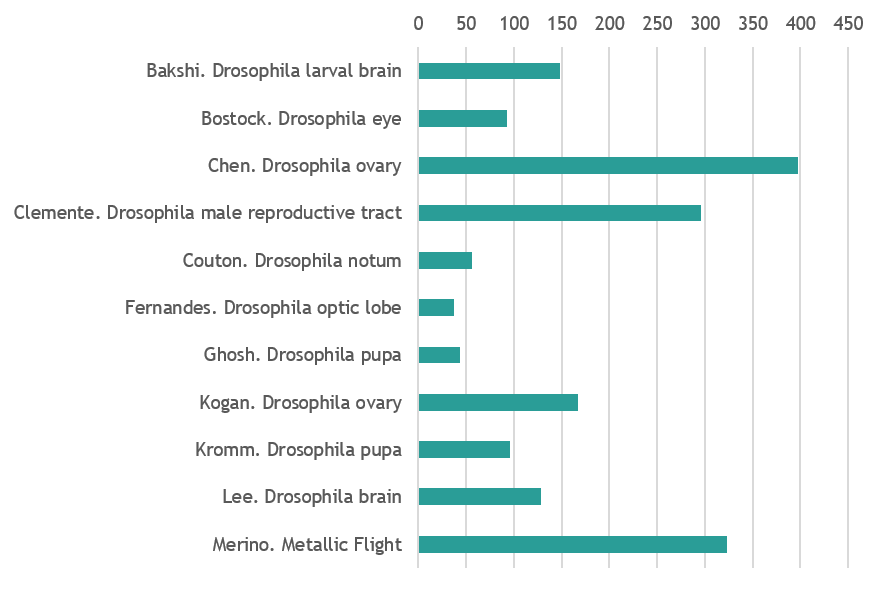
Drosophila (vote for 2 out of 11)
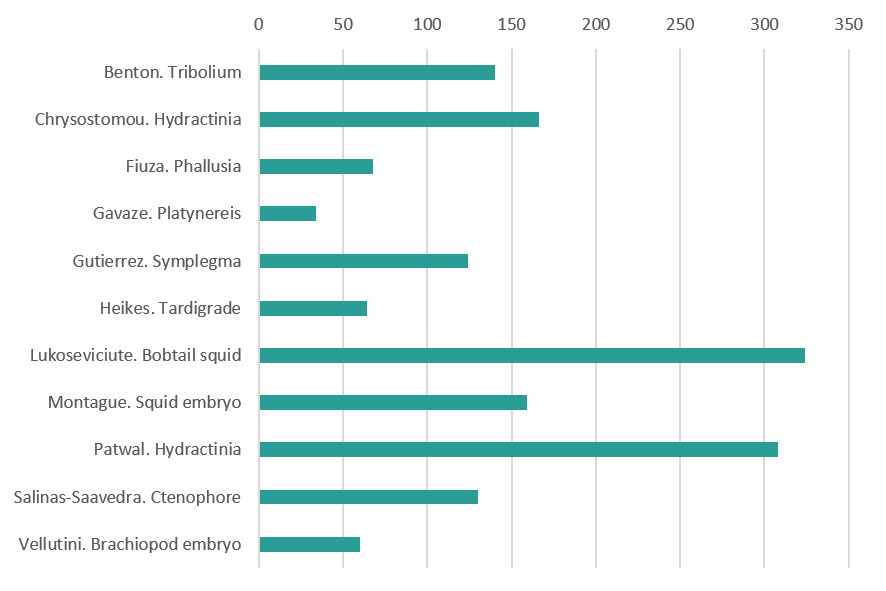
Invertebrate variety show (vote for 2 out 11)
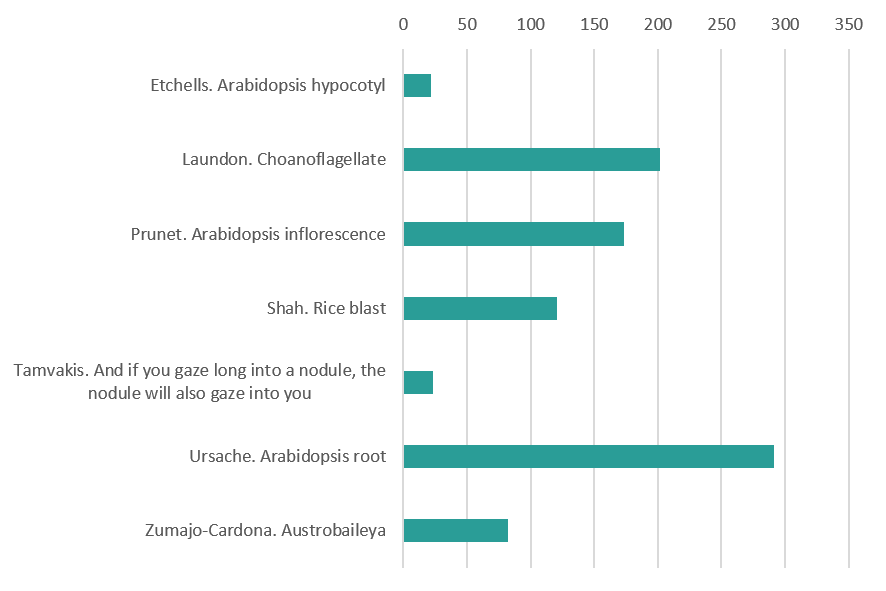
Plants, Fungi and Choanoflagellates (vote for 1 out of 7)
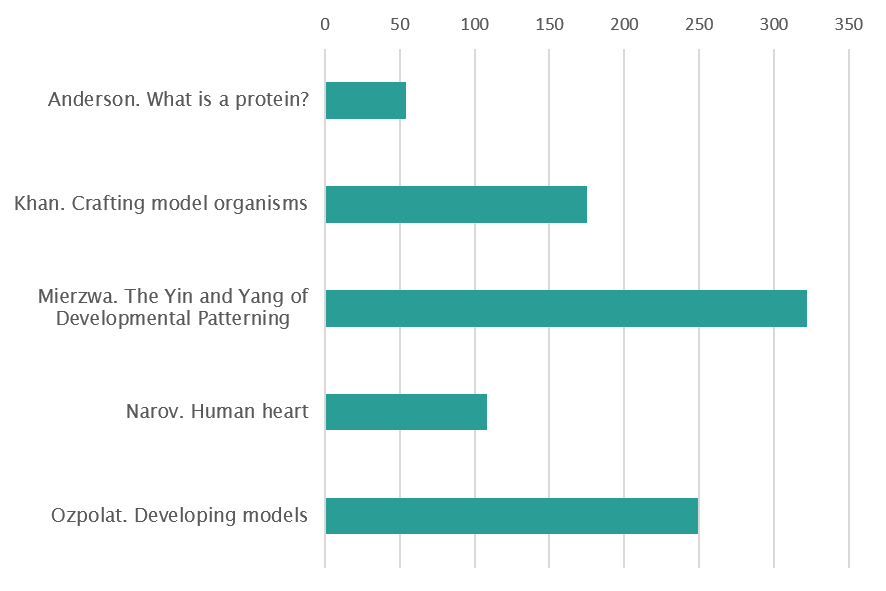
Art and Illustration (vote for 1 out of 5)
The cut offs are a little arbitrary but it’s the best scheme we could come up with. Inevitably, many beautiful images are going to miss out, but we hope the selection stands alone in showcasing the aesthetic side of research.
The pictures are arranged in galleries – click to expand the image and see the caption (there’s also a link to see the image full size). Below the galleries are independent polls to pick your favourites from each section. Both the galleries and the polls are arranged alphabetically by creator, and the poll text is the same as the file name (e.g. ‘Blin. mESC micropatterns’) which you can see below the caption. Please just vote once (well, twelve times!) – polls are set up to stop repeat voters by cookie.
Voting closes on Sunday 19 May 23:59 GMT
You can also let us know what your overall favourite is in the comments. Happy voting.
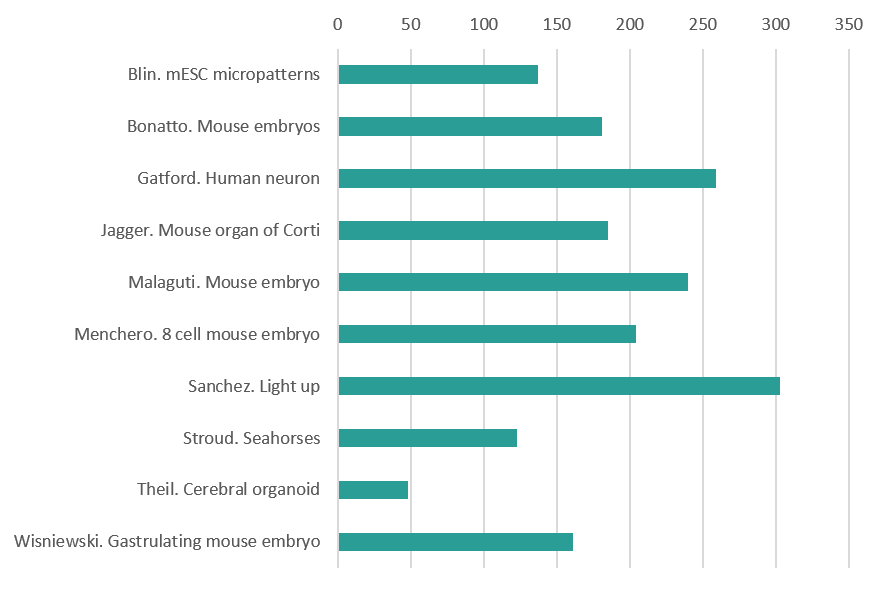
Mammals (Mus and Homo)
Epiblast Stem Cells cultured on micropatterns of various geometries. The cells were stained for LaminB1 (Green), Tbra (Blue) and GM130 (Red). By Guillaume Blin (MRC Centre for Regenerative Medicine, The University of Edinburgh)
Mouse embryos at E12.0 nuclei stained in 1) control and 2) compound mutants for AER-FGFs with a graded loss of fore and hindlimbs. By Christian Bonatto (National Cancer Institute, NIH)
Super-resolution image acquired via structured illumination microscopy of a single developing human neuron ectopically expressing the autism risk gene neuroligin-4X with enlarged growth cones. F-actin is in green, HA-tagged neuroligin-4X is in magenta, doublecortin is in cyan confirming its neuronal identity, and the nuclear marker DAPI is in grey. By Nicholas Gatford (Institute of Psychiatry, Psychology and Neuroscience, King’s College London)
The image shows a confocal section of the organ of Corti (sensory epithelium of the cochlea) in the inner ear of a neonatal (P1) mouse. Actin is stained using phalloidin (cyan), nerve fibres using anti-β3-tubulin (yellow), and nuclei using DAPI (red). This is a developmental stage before the onset of air-borne hearing (which occurs around P11 in mice), and during a period of path-finding by afferent nerve fibres, and refinement and pruning of their terminals. Many fibres temporarily contact non-sensory supporting cells, before they establish permanent synaptic connections with the mechano-sensory hair cells. By Daniel Jagger (UCL Ear Institute)
Transverse section of a chimaeric E6.5 mouse embryo generated by aggregation of Id1-Venus reporter ES cells with a wild-type morula, stained for Id1-Venus (green), Nanog (red) and T (blue). By Mattias Malaguti (MRC Centre for Regenerative Medicine The University of Edinburgh)
8-cell mouse embryo which has just aquired apical-basal polarity but we can still appreciate individual cells. E-Cadherin is labelled in red and pERM is labelled in white. By Sergio Menchero (CNIC Madrid)
E14.5 mouse embryo labeled for cartilage (Sox9-GFP, in biop-SpringGreen) and vasculature (highlighter ink circulated by injection in a blood vessel, in mpl-magma). Vasculature “lights up” the embryo, including within the developing bones of the limbs. Image taken using a microscope kindly sponsored by Zeiss during the 2018 Embryology Course at the Marine Biological Laboratory in Woods Hole, MA. By Paul Gerald Layague Sanchez (EMBL Heidelberg)
Magnified view of the aortic and mitral valves in a developing mouse heart. By Matt Stroud (BHF Centre of Excellence, King’s College London)
Cross section through a 36 days human cerebral organoid stained with Ascl1 (red) and Arl13b (green) to reveal a subset of cortical progenitors and the primary cilium projecting into the lumen of the neuroepithelial rosette, respectively. Nuclei are counterstained with DAPI. By Thomas Theil (Centre for Discovery Brain Sciences, The University of Edinburgh)
Gastrulating mouse embryo. Nuclear envelope labelled with LaminB1 (grey). Primitive-streak and ingressing mesoderm labelled with T-Brachyury (magenta). By Darren Wisniewski (MRC Centre for Regenerative Medicine The University of Edinburgh)
Please pick your favourite 2 images
(please remember to pick 2)
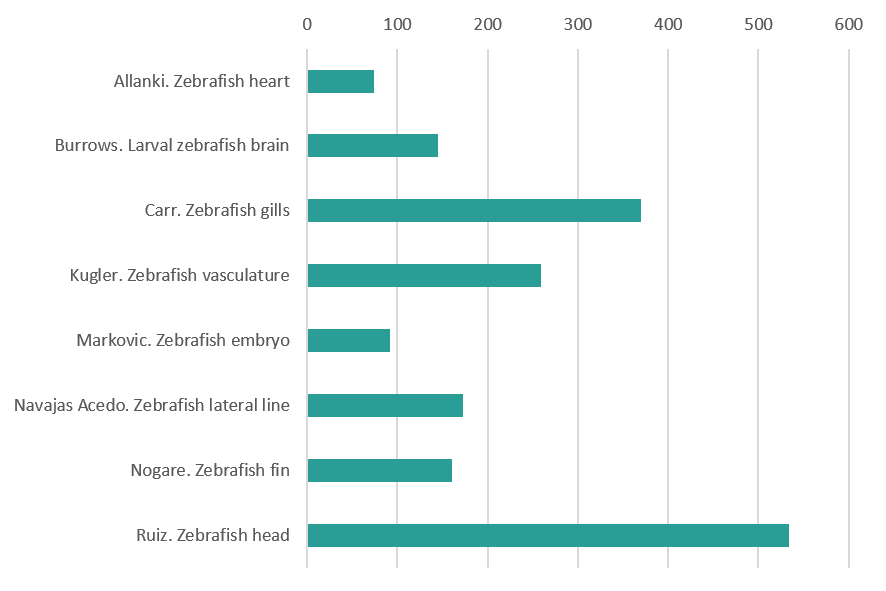
Zebrafish
Regenerating zebrafish heart after cryoinjury. By Srinivas Allanki (Max Planck Institute for Heart and Lung Research)
Larval zebrafish at 6 days post fertilisation registered to a regional neuroanatomical zebrafish atlas. By Dominic Burrows (MRC Centre for Neurodevelopmental Disorders, King’s College London)
Developing gill vasculature in a 120h old zebrafish and also features the heart. This image was taken using lightsheet microscopy in two transgenic lines, one that marks the endothelial actin and the other marks the endothelial nuclei. After acquisition it was processed as a colour coded depth projection. By Philippa Carr (Bateson Centre, University of Sheffield )
Vasculature in a double-transgenic line (Tg(kdrl:HRAS-mCherry)s916, Tg(fli1a:CAAX-eGFP)), visualized using light-sheet fluorescence microscopy.The three individual images are part of a timelapse acquisition, showing 50 hours post fertilization (hpf), 60hpf and 70hpf. Essential processes such as anastomosis (eye and fin), joining of vascular beds (head and spinal cord), and remodelling (subintenstinal vein and duct of Cuvier) can be observed.By Elisabeth Kugler (Department of Infection, Immunity & Cardiovascular Disease,University of Sheffield)
48 hpf zebrafish embryo expressing GFP in sensory neurons and expressing mCherry where the MAP Kinase Kinase Kinase LZK (map3k13) is expressed. LZK is a vertebrate homologue of dlk-1, which is essential for neuronal regeneration in C. elegans and Drosophila. By Hannah Markovic (UCLA)
The image shows the Lateral Line primordium of a zebrafish embryo labeled with a green fluorescent membrane protein. This group of cells migrates together from the head of the animal to the tip of the tail in a journey in which it will form proto-neuromasts or rosettes, which will be later deposited as clusters of cells called ‘neuromasts’. By Joaquin Navajas Acedo (Stowers Institute for Medical Research)
Spinning disc confocal image of mitochondria in the developing zebrafish fin at ~96hpf. By Damian Dalle Nogare ( Eunice Kennedy Shriver National Institute of Child Health and Human Development, NIH)
Transgenic zebrafish (Danio rerio) larva expressing red fluorescent protein in the developing mouth and olfactory epithelium. A subset of cells also express a construct that labels actin filament with green fluorescent protein. DAPI (blue) is used to label DNA in the nuclei of all cells. By Oscar Ruiz (Department of Genetics MD Anderson Cancer Center)
Please pick your favourite 2 images
(please remember to pick 2)
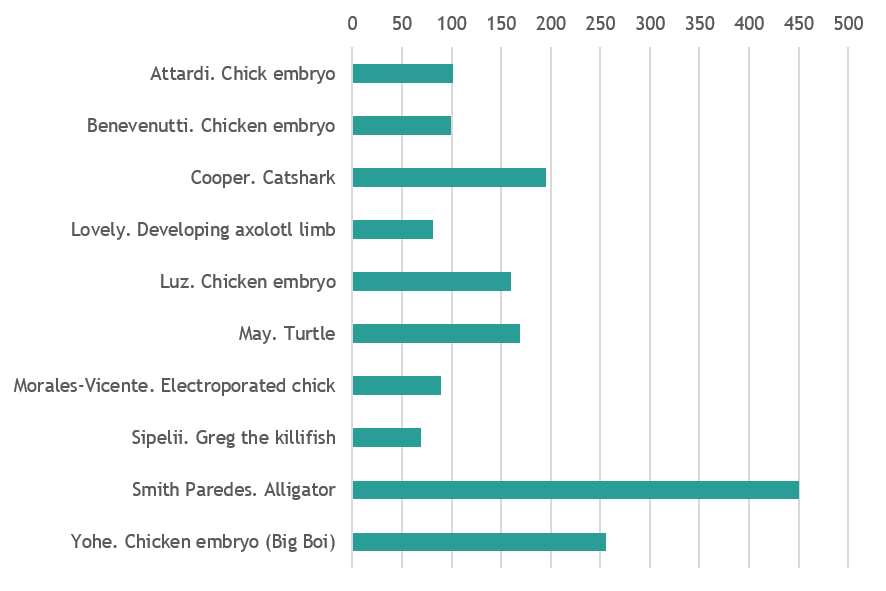
Vertebrate variety show
Chicken embryo at stage HH9. Picture was taken with an iPhone through the ocular lens of a dissecting stereomicroscope, after injecting the space between the yolk and the embryo with blue pen ink. The ink enables us to see the transparent embryo and highlights the membranes surrounding it as well as some yolk granules. The picture was taken by me at the Embryology Course 2018, Marine Biological Laboratory, Woods Hole – MA.By Andrea Attardi (Max Planck Institute for Molecular Cell Biology and Genetics, Dresden)
13-day chicken embryo. Diaphanization protocol with Alcian blue and alizarin red. By Felipe Zanghelini Benevenutti (Federal University of Santa Catarina, Brazil)
Scanning electron micrograph of hatchling catshark (Scyliorhinus canicula) dermal denticle (scale). By Rory Cooper (Animal and Plant Sciences University of Sheffield)
Our lab works to understand the regenerative ability of the axolotl salamander, and a key step in understanding how a limb can regenerate is to understand how the limb was initially developed. This image depicts a developing larval axolotl limb bud which has been stained for 3 key developmental genes. The genes shown are: FGF8 (Cyan), SHH (yellow), and PRRX1 (magenta). The method used to stain the genes in this image was fluorescent in situ hybridization, a method for directly labeling the mRNA within a cell. The image was generated using a confocal microscope. By Alexander Lovely (Department of Biology, Northeastern University, Boston)
13-day chicken embryo with diaphanization protocol with Alcian blue and alizarin red. By Daniely Ramos Luz (Federal University of Santa Catarina, Brazil)
My entry for the photo contest is an unhatched sea turtle. I took a picture of this little guy when I was working with sea turtle monitors at the Yawkey Wildlife Center in South Carolina. I helped to monitor the nests and would often find unhatched turtles which failed to resorb their yolk and emerge from the nest. By Catherine May (Boston College)
Early developed chicken embryo (HH9) electroporated in the left side. The electroporated side is detected by a GFP reporter and displayed as green dots on the beautiful chicken embryo. By David Morales Vicente (University of São Paulo, Brazil)
This is Greg. Greg is a 1 month old Austrofundulus limnaeus annual killifish who is the nicest juvenile in my undergraduate research experiment. He likes brine shrimp, blood worms, and would like to have his picture featured on a magazine if possible. Greg is a part of my experiment where I am trying to see if this particular fish species sex ratio is affected by fluctuating temperature patterns. Greg wants you to give him a chance. Trust in Greg. By Motutama Sipelii (Portland State University)
Alligator mississipiensis embryo at stage 13-14 immunostained against Myosin heavy chain showing the developing muscles and (red) and neurofilament labeling axons of nerves. By Daniel Smith Paredes (Department of Geology and Geophysics, Yale University)
Stage 35 chicken embryo, cleared and immunostained for DAPI (orange) and Pax3 (cyan) demonstrating the developing neural crest and spinal cord. Image was taken on the Nikon AZ-C2 macro-confocal with image analysis performed in Imaris. Image was taken in collaboration with Andrea Attardi at the Max Planck Institute of Molecular Cell Biology and Genetics during the Woods Hole 2018 Embryology course. By Laurel Yohe (Department of Geology and Geophysics, Yale University)
Please pick your favourite 2 images
(please remember to pick 2)
Drosophila
Micrograph shows Drosophila larval brain attached to the leg and eye imaginal discs. There is a glial migration from brain to these discs as indicated by glial cells (shown in red) in the imaginal discs, along with corresponding repo-gal4 driving GFP (glial marker). Glial cells are stained by anti-repo (red), Neural stem cells are marked by anti-Dpn (blue) and glial membranes are marked by repo-gal4::GFP (green). By Asif Ahmad Bakshi (Centre for DNA Fingerprinting and Diagnostics, Hyderabad, India)
The development of the Drosophila lamina neuropil occurs under tight spatiotemporal control that involves a signal relay between photoreceptors (gray), glia (cyan) and lamina precursor cells (magenta). Visualised in my image is a bundle of (joy) photoreceptor axons innervating and signalling to neuroepithelia located at the surface of the optic lobe. With this signal, the neuroepithelia acquire lamina precursor cell identity and form beautifully organised columns. The wrapping glia, in the optic stalk, also receives signalling molecules from the innervating photoreceptors and helps to promote differentiation in lamina precursor cells. Zeiss LSM confocal microscope. By Matthew Bostock (Department of Cell & Developmental Biology, UCL)
Drosophila whole ovary stained for f-actin (Red), nuclei (Cyan) and actin (Green). By Yujun Chen (Kansas State University Division of Biology)
Reproductive organ of a young (1-3 days old) adult Drosophila male. The male reproductive tract consists of a pair of testes, where mature sperm is produced. The sperm is temporarily stored in the seminal vesicles (sv) before being released into the ejaculatory ducts (ED). Here, the sperm is mixed with the seminal fluid produced in the accessory glands (AGs). Ejaculatory bulb (EB) is also visible. The sample expresses nuclear RFP (cyan) driven by the Gal4 driver traffic jam (tj) and it was stained for Actin (magenta) and the germline stem cell marker vasa (green). Image was acquired at the Wolfson Bioimaging Facility (University of Bristol). Scale Bar: 200 um. By Giuliana Clemente (University of Bristol)
Drosophila notum as the sensory organ precursors undergo their first division, seemingly shooting across the field. Partner of Numb ( PoN) is shown in green and microtubules (as reported by Jupiter) in Magenta. The picture is color blind friendly! By Louise Couton (Department of Biochemistry, University of Geneva)
A neuroepithelium called the ‘outer proliferation centre’ (OPC) together with the eye disc epithelium (top right) are shown in Cyan. The OPC is a broad crescent shaped structure that generates neurons of the lamina and medulla neuropils. All glia are shown in magenta. They are present in the eye disc, through the optic stalk and in the optic lobe. By The Fernandes Lab (Department of Cell & Developmental Biology, UCL)
The image is a lightsheet micrograph of a Drosophila undergoing the final stages of development(~24hrs prior to ecclosion). The anterior half of the developing fly has been dissected out from the pupal case. The posterior pupal case can still be seen in the upper part of the image. The fluorescent marker is DE-cadherin- lining apical junctions of cells. There is a surreal resemblance to a developing human foetus during its late stages- when all the major morphological development is complete. Although, the individual seems suspended in time and space, there are major internal structures still developing. By Suhrid Ghosh (Max Planck Institute of Molecular Cell Biology and Genetics, Dresden)
Drosophila melanogaster ovary 24 hours after pupal formation stained for Lamin-C and Fasciclin III (in red), VASA (green), Traffic Jam (white), and DAPI (blue). By Lena Kogan (Biological Sciences Department, Columbia University NY)
Drosophila melanogaster pupa expressing a histone marker. It was imaged on our MultiView Light-Sheet Microscope (MuVi-SPIM) at 10x magnification. By Dimitri Kromm (EMBL Heidelberg)
Combined single molecule Fluorescent in situ hybridisation and immunofluorescence of developing Drosophila larval brain. Shown here are Syncrip protein (green), syncrip RNA (red) and DNA (blue). By Jeff Lee (Dept. of Biochemistry, University of Oxford)
Drosophila mutant showing a decreased eye size compared to wild type. This line is not able to generate descendants with wild type flies. By Marisa Merino (Department of Biochemistry, University of Geneva)
Please pick your favourite 2 images
(please remember to pick 2)
Invertebrate variety show
Live embryo of the beetle Tribolium castaneum. During early development, this egg was injected with mRNA encoding a photoconvertible fluorescent protein that is localised to nuclei. Several nuclei were then irradiated with short-wavelength light to induce a conformational change in the fluorescent protein, thereby causing it to fluoresce in a different wavelength (shown here in magenta rather than the unconverted cyan). This strategy allows the targeted labeling of one or more cells and the tracing of those cells and their descendents throughout development. In this case, I labeled a group of cells came to form part of an extraembryonic tissue that covers the embryo and supports its development. By Matt Benton (Department of Zoology, University of Cambridge)
Developing oocytes (red) in the marine cnidarian Hydractinia symbiolongicarpus enclosed in sporosacs – cell boundaries are in green and nuclear staining in blue. By Eleni Chrysostomou (Centre for Chromosome Biology & Regenerative Medicine Institute, National University of Ireland Galway
Embryonic development of the ascidian Phallusia mammillata. 3D rendering of membrane imaging with light-sheet microscopy covering the zygote, cleavage, gastrulation, neurulation, tailbud and larval stages. By Ulla-Maj Fiuza (EMBL Heidelberg)
Small worm of Platynereis dumerilii (annelid) with dividing cells, in the brain and the posterior part, labelled in green (EdU incorporation and chase). By Eve Gazave (Institut Jacques Monod, Paris)
My picture is of an interesting and uncommon animal: a larva of the colonial tunicate Symplegma rubra, after metamorphosis. The five projections are the ampullae, the first structures of the blood vessels. In the center, there is the oozooid, during organogensis. Some larval organs are seen, such as the ocellus (red circular structure). I observed this larva in the marine station Cebimar (Centro de Biologia Marinha da Universidade de São Paulo). By Stefania Gutiérrez (University of São Paulo, Brazil)
Tardigrade embryo (Hypsibius exemplaris) with membranes and mitochondria labeled. By Kira Heikes (UNC Chapel Hill.)
Live Hawaiian Bobtail Squid (Euprymna scolopes), stained with vital dyes (CellMask, LysoTracker and Hoechst) to understand its cellular and sub-cellular organisation during development. Blue is labelling cellular nuclei, green – cell plasma membranes and red – lysosomes that are important for cellular waste removal. This species is a candidate model organism that yet holds many answers to developmental biology questions, such as nervous system and eye development. The image was taken during the MBL 2018 Embryology Course with the confocal microscope provided by Zeiss. Animals were supplied by the cephalopod researcher Carrie Albertin. By Martyna Lukoseviciute (Weatherall Institute of Molecular Medicine, University of Oxford)
Squid embryo (Doryteuthis pealeii) with nuclei (pink), actin (cyan) and neurons (green). By Tessa Montague (Zuckerman Institute, Columbia University NY)
DIC and fluorescence image of Hydractinia male sexual (left) and feeding polyps (right) on a chitin bed. Chitin is shown in green. Noncycling cells probed with cyclin-dependent kinase inhibitor (CDKI) are shown in yellow, which are mainly in nematocytes, male gonophore, and gastrodermis. By Indu Patwal (Centre for Chromosome Biology, National University of Ireland Galway)
Ctenophore gastrula. Cyan: DAPI; Yellow: tubulin. By Miguel Salinas-Saavedra (Centre for Chromosome Biology, National University of Ireland Galway)
Surface view of an embryo of the brachiopod Novocrania anomala during early gastrulation. Image shows the cell membrane outlines in the ectodermal surface as stained by BODIPY-FL (F-actin staining). Animal pole is top and vegetal pole is bottom where the blastoporal opening is visible. Adults were collected in 60 meter depth waters near Storingavika in Bergen, Norway, spawned and fertilized in the laboratory. Image is a maximum intensity projection of the original stack taken on a Leica SP5 confocal microscope. By Bruno Vellutini (Max Planck Institute of Molecular Cell Biology and Genetics, dresden)
Please pick your favourite 2 images
(please remember to pick 2)
Plants, Fungi and Choanoflagellates
Transverse section of an Arabidopsis hypocotyl with disrupted vascular organisation. By Peter Etchells (Durham University)
3D TEM reconstruction of the colonial choanoflagellate Salpingoeca rosetta. Choanoflagellates are the closest unicellular relatives to the animal kingdom and some species such as S. rosetta are capable of developing into multicellular colonies. This makes S. rosetta a powerful model to investigate the development, origin and evolution of animal multicellularity. Shown are apical vesicles (pink), food vacuoles (green), endocytotic vacuoles (fuschia), ER (yellow), extracellular vesicles (grey), filopodia (external, purple), flagellar basal body (light blue), flagellum (dark green), glycogen storage (white), Golgi apparatus and vesicles (purple), intercellular bridges (external, yellow; septa, red), large vesicles (brown), microvillar collar (light orange), mitochondria (red), nonflagellar basal body (dark orange), and nuclei (dark blue). By Davis Laundon (The Marine Biological Association, Plymouth UK)
An Arabidopsis inflorescence expressing a fluorescent reporter for the APETALA3 gene, which promotes petal and stamen identity (green). Cell walls were stained with rpopidium iodide (magenta). By Nathanaël Prunet (Department of Molecular, Cell and Developmental Biology, UCLA)
Development of the three-cell asexual spore of the rice blast fungus. The spore starts as a swelling of the aerial hypha, changing from a sphere, to a symmetrical oval two cell stage and finally transforming into the spindle shaped spore seen at the end of the montage. By Hiral Shah (Bharat Chattoo Genome Research Centre The Maharaja Sayajirao University of Baroda Gujarat India)
Root of Medicago truncatula treated with auxin transport inhibitors, known to induce structures similar to symbiotic nodules. By Ioannis Tamvakis (Sainsbury Laboratory University of Cambridge)
The development of a lateral root in Arabidopsis thaliana. The sample has been cleared and stained with Calcofluor White to outline the cell walls and the green fluorescent nuclei represent a protein expressed specifically in the outer cell layer of developing lateral root. By Robertas Ursache (University of Lausanne, Switzerland)
Cross section through a developing fruit (carpel) of Austrobaileya scandens. The photograph was taken with a light microscope under 10X magnification and stained with safranin and alcian blue to have a better tissue contrast. The fruit of A. scadens is formed by multiple individual carpels, here we are looking at one of those that is going to grow fleshy in order to disperse the seed. By Cecilia Zumajo-Cardona (City University of New York, New York Botanical Garden)
Please pick your favouriteimage
Art and illustration
A single amino acid, a dot ( . ), becomes a chain of amino acids – a protein, a line (——), moves in space to become a plane, then a volume. Like in drawing, a dot progresses to a line, to meander in space. So it seems that drawing might be an appropriate way to experiment, to ‘be like’ the protein. Proteins pervade life and collectively they take an infinite variety of forms. Protein’s fold, most of the time this is what a protein is trying to do. The folded protein is the relaxed protein, in its ‘natural’ state, not wasting any energy. Folding is a process, it happens through time, in an ‘energy landscape’ often imagined to be conical (cone shaped) with the folded protein resting in the very bottom – the point of least energy in the landscape. As the protein moves from the top of this landscape to the bottom (it might not make it) it embarks on an explorative journey of the space; body and environment continuously co-creating each other. There are fast and slow tracks, uphills and downhills, dead-ends and if in trouble then a ‘chaperone’ will come to help find the way together. This dynamic process is hard to imagine and the images you find in scientific textbooks don’t exactly give the game away (generally a cone, a few uphills or downhills). How else can we imagine this complexity? What other images could we see? After making countless drawings with molecular biologist JJ Phillips, we take a day to look at them all together: drawings of cones, of simple figures moving in cones, colour coding energy levels, line drawings of movement series, planes in space. It struck me that the bottom of the cone is like the centre of a maze. We like the idea. We try drawing a maze. First we need an underlying grid architecture, not a generic pattern but one that is true to the dynamic pattern of the protein journeying. We change the granularity of the grid for different stages of the process. Colour brings dimensions and guides the pathways that are created. The environment and the body co-create one another as the maze undermines the distinction between figuration and abstraction. The drawing process has its own stochasticity and the image, following its own creative process, reveals something true of the dynamic pattern of protein folding. The idiosyncrasies of this living process have informed and given rise to a new artistic process. ‘Organic development in a work of art is at least analogous to, and probably identical with, organic development in nature; in an organic-artistic scheme the essence of art is in processes rather than its products; and such artistic ‘events’ as are thrown up are significant merely in that they reflect past, present and future aspects of the dominant process’. (Thistlewood, 1981). By Gemma Anderson (University of Exeter and Falmouth School of Art, http://www.gemma-anderson.co.uk/)
These are three hand-crafted coasters, made from perler beads. They represent pictures of three classic developmental biology model organisms, the fruitfly Drosophila melanogaster, a hatching chick Gallus gallus domesticus, and the flower of Arabidopsis thaliana. Pertinent to their natural habitat, the fruitfully is shown in a blue background representing its flight in the sky, the chick is shown in a green background representing the grass it forages and the flower is shown in a brown background representing the soil it grows in. These were created by me for use by any artsy developmental biologist who can use this during coffee breaks in lab. By Sumbul Jawed Khan (Sci-Illustrate and https://www.linkedin.com/in/sumbul-jawed-khan/)
This drawing illustrates the remarkable self-organization capacity of cerebral organoids that allows them to recapitulate human brain development in vitro. Each color represents a different type of cell, and the dorsal and ventral areas are separated by a defined boundary – like a yin and yang symbolizing the balance between distinct but complementary entities. By Beata Edyta Mierzwa (Ludwig Institute for Cancer Research and the University of California, San Diego, and www.beatascienceart.com)
The image depicts different views (cranial, ventral, dorsal, right, caudal and left) of a digital 3D model of the embryonic human heart at Carnegie stage 12. The model is based on work from Antoon Moorman’s group at the Academic Medical Centre in Amsterdam. My model is an example of scientific illustration, and is not volume-reconstructed from histological sections, confocal images or micro-CT data. By Kalin Narov (Embryo Safari, https://www.embryosafari.com/)
This drawing combines embryos and structures from embryos. Featuring: Bats, Drosophila, Xenopus, Parhyale, Ascidians, Chicken. By B. Duygu Özpolat (Eugene Bell Center for Regenerative Biology and Tissue Engineering, Marine Biological Laboratory Woods Hole)
Please pick your favouriteimage
Thanks for voting – if you made an error, just email aidan.maartens@biologists.com and we’ll correct the numbers!
 In this episode from our series exploring 100 ideas in genetics, we’re taking a trip to London with William Bateson and discovering that the famous story about him reading Mendel’s paper on the train might not be all that it seems.
In this episode from our series exploring 100 ideas in genetics, we’re taking a trip to London with William Bateson and discovering that the famous story about him reading Mendel’s paper on the train might not be all that it seems. (No Ratings Yet)
(No Ratings Yet)




 (2 votes)
(2 votes)






























")


































 (20 votes)
(20 votes)