International Institute of Molecular and Cell Biology in Warsaw
Laboratory of Protein Metabolism in Development and Aging
is seeking a talented Postdoctoral fellow
Location: Warsaw, a vibrant city with an international academic research environment. International Institute of Molecular and Cell Biology (www.iimcb.gov.pl) – one of the most dynamic and top ranked Polish research institutes.
Job description: Laboratory of Protein Metabolism in Development and Aging, which will be opening on August 2017, is seeking highly motivated and talented Postdoc to join young team investigating the protein homeostasis in development and aging. We use both genetic, molecular and biochemical approaches, primarily in the C. elegans, to study proteolytic networks. Postdoc fellowship is funded in frame of National Science Centre OPUS grant.
Summary: Organismal development or environmental stimuli challenge the homeostatic protein balance (proteostasis) of individual cells, tissues or the entire organism. The ubiquitin proteasome system (UPS) is a key determinant of proteostasis as it regulates the turnover of damaged proteins supporting cellular protein homeostasis and thereby maintains the proteome during stress and aging. Our long-term objective is to understand the mechanistic and developmental aspects of protein degradation pathways defined by combinations of particular ubiquitin ligases (E3). The identification of stress and aging-induced signals that coordinate the interplay between specific E3s will offer intriguingly new mechanistic insights how proteolytic networks are fine-tuned to maintain the cellular proteome and support development and longevity.
Qualifications:
• PhD (or be close to completion) in Molecular Biology, Cell Biology, Protein Chemistry, Genetics or a related discipline;
• experience in C. elegans or cell culture is an advantage;
• experience in Next Generation Sequencing techniques (RNA-Seq, ChIP-Seq) and genomic engineering is an advantage;
• good writing and oral communication skills in English, and competence in scientific writing.
How to apply:
Please send your application including CV, motivation letter and the list of publications to the e-mail address: wpokrzywa@iimcb.gov.pl, until 10th December 2017. Thanking all applicants for their interest, we will contact only selected candidates for an interview.
Please include in your application the following statement: “In accordance with the personal data protection act from the 29th of August 1997, I hereby agree to process and to store my personal data by the Institution for recruitment purposes”.
The recruitment procedure fulfills the National Science Centre’s regulations on granting the scholarships to young scientists.
Selected publications:
Riga T*, Pokrzywa W*, Kevei E, Akyuz M, Vishnu Balaji, Svenja Adrian, Hoehfeld J, Hoppe T. (2017). The ubiquitin ligase CHIP integrates proteostasis and aging by regulation of insulin receptor turnover. Cell. 169: 470-482.
Ackermann L., Schell M., Pokrzywa W., Gartner A., Schumacher B., Hoppe T. (2016). E4 ubiquitin ligase specific degradation hubs coordinate DNA double strand break repair and apoptosis. Nat Struct Mol Biol. 23: 995-1002.
Kaushik S, and Cuervo AM (2015). Proteostasis and aging. Nat Med. 21, 1406-15
Frumkin A, Dror S, Pokrzywa W, Bar-Lavan Y, Karady I, Hoppe T, Ben-Zvi A. (2014). Challenging muscle homeostasis uncovers novel chaperone interactions in Caenorhabditis elegans. Front Mol Biosci., doi: 10.3389
van Oosten-Hawle P, and Morimoto RI (2014). Organismal proteostasis: role of cell-nonautonomous regulation and transcellular chaperone signaling. Genes & Dev. 28: 1533-43.
Segref A, Kevei E, Pokrzywa W, Mansfeld J, Schmeisser K, Livnat-Levanon N, Ensenauer R, Glickman M.H, Ristow M, Hoppe T. (2014). Pathogenesis of human mitochondrial diseases is modulated by reduced activity of the ubiquitin/proteasome-system. Cell Metab. 4:642-652.
Pokrzywa W. and Hoppe T. (2013). Chaperoning myosin assembly in muscle formation and aging. Worm. 2:e25644.
Gazda L*, Pokrzywa W*, Hellerschmied D, Loewe T, Forné I, Mueller-Planitz F, Hoppe T, Clausen T. (2013). The myosin chaperone UNC-45 is organized in tandem modules to support myofilaments formation in C. elegans. Cell. 1, 183-195.
Kuhlbrodt K, Janiesch PC, Kevei E, Segref A, Barikbin R, and Hoppe T (2011). The Machado-Joseph disease deubiquitylase ATX-3 couples longevity and proteostasis. Nat Cell Biol. 13, 273-81.
We are looking for a highly motivated Senior Laboratory Scientist to join the quantitative cell biology laboratory headed by Dr Silvia Santos. The lab is a new addition to The Francis Crick Institute in London and focuses on understanding control principles in cell decision-making. Current areas of research include understanding control of cell division and differentiation, using human embryonic stem cells as a model system. There is a strong focus on single cell analysis and live cell imaging approaches. The team is currently composed of three PhD students and two post-doctoral fellows.
The successful post holder is expected to drive his/her own research, help with lab management and training and support on-going research projects. The ideal candidate is likely to be an energetic, focused and productive individual with a desire to work on interesting biological problems in a collegial and collaborative work environment. Excellent time management and organisation skills are essential.
PROJECTSCOPE
The decision to divide is a fundamental cellular decision and the conserved networks that trigger cell division adapt and remodel in a variety of biological contexts including developmental transitions and malignancy. We have been exploring spatio-temporal control of cell division in mammalian cells and remodelling of cell cycle networks during developmental transitions, using embryonic stem cells as a model system.
Embryonic stem cells have the propensity to differentiate into the three germ layers. The switch between pluripotency and differentiation in these cells has been our paradigm of choice to understand how protein and gene networks decode cellular signals and thereby encode irreversible commitment to different cell fates.
The molecular basis of these decisions is of fundamental biological importance and have significant clinical applications.
Informal enquires can be sent to Silvia Santos at: silvia.santos@crick.ac.uk
If interested please apply here with a cover letter stating your background and motivation, CV and names of two referees.
The story behind our recent paper: Liu Z*, Wang L*, Welch JD*, Ma H, Zhou Y, Vaseghi HR, Yu S, Wall JB, Alimohamadi S, Zheng M, Yin CY, Shen WN, Prins JF, Liu JD, Qian L (2017). Single cell transcriptomics reconstructs lineage conversion from fibroblast to cardiomyocyte. Nature, 551, 100-104 *: co-first author. https://www.nature.com/articles/nature24454
The Journey Started
In February 2014, I graduated from Indiana University with a PhD in Microbiology and Immunology. Being fascinated by regenerative medicine and how cell fate can be changed/ reversed during reprogramming, I joined Dr. Li Qian’s lab at University of North Carolina (UNC) in March 2014. Until today I still feel things just happened so fast and I am such a lucky person to be a part of this group. So I started my new journey at the beautiful small town of Chapel Hill to become a “reprogrammer.” Previously, Dr. Qian discovered that introduction of three cardiac transcription factors Mef2c, Gata4, and Tbx5 into mouse heart after myocardial infarction could significantly improve the heart’s function by converting the local fibroblasts in the heart to beating cardiomyocytes – which we termed as the process of “direct cardiac reprogramming” or “induced cardiomyocytes (iCM) reprogramming.” In addition to iCM, other researchers have reported the conversion of fibroblasts into induced neurons, hepatocytes, β cells, and so on. Direct lineage conversion not only offers a new strategy for tissue regeneration and disease modeling, but also provides a unique platform for understanding cell fate.
The Cell, the Single Cell
The cell is the most basic functional unit of an organism. The human body contains more than 40 trillion cells that are from different origins, form different organs, and carry out different functions. Each cell has its unique transcriptome, proteome, and functional role even though all cells originate from the same zygote and in theory share the same genome. Due to technical limitations, researchers were only able to check either the expression of limited number of genes at the single cell level (like immunofluorescence staining) or the transcriptome but at the price of averaging signals from a population of cells (like bulk RNA-sequencing). As the dawn of the era of single cell OMICs, we realized that we could do more now to understand the mechanisms of cell fate determination.
Direct Cardiac Reprogramming
Direct cardiac reprogramming shows promise as an approach to replenish lost cardiomyocytes in diseased hearts and its utilization of local scar-forming fibroblasts adds to its chance of potential clinical application. Considerable efforts have been made to improve the efficiency and unravel the underlying mechanism. However, it still remains unknown how the conversion of fibroblast into cardiomyocyte is achieved without following the conventional cardiomyocyte specification and differentiation. The reprogramming process is inherently heterogeneous in that the starting cells (primary fibroblasts) exhibit uncharacterized molecular heterogeneity and they don’t reprogram at the same pace, rendering it difficult to study using conventional bulk RNA-seq. Therefore, we decided that we were going to leverage the power of single-cell transcriptomics to really dissect the cellular and molecular mechanisms of iCM reprogramming.
Figure 1. In vitro conversion of neonatal mouse cardiac fibroblasts into iCMs. Fluorescent images of the cardiac reporter αMHC-GFP and two cardiac structural proteins cTnT and α-actinin were shown. Nuclei were stained with DAPI. Scale bar = 200 µm. The middle panels are enlarged from left panels showing striated sarcomere structure. Figure adapted from paper: Wang L*, Liu Z*, Yin C, Asfour H, Chen O, Li Y, Bursac N, Liu J, Qian L (2014). Stoichiometry of Gata4, Mef2c, and Tbx5 influences the efficiency and quality of induced cardiac myocyte reprogramming. Circulation Research, 116 (2), 237-244. *: co-first author. http://circres.ahajournals.org/content/116/2/237.full
Collaborator Hunting
Me and Li Wang at old well, UNC
Everything was perfect and exciting except that we had never done single cell RNA-seq at that time. Not many people around have either. So now, it’s time for the wonderful collaborators to come to the stage! First is the co-first author of the paper, my dear colleague and friend Dr. Li Wang, also from the Qian lab. This wonderful lady joined the lab earlier than me; she set up the experimental platform for everyone who joined after her and literally taught everyone hand-by-hand all the technical details to become a “reprogrammer”. This was our second project to co-first author. We designed and performed all the single cell experiments together. We discussed the results along the way and encouraged each other when things were not working. With the help of the UNC Advanced Analytics Core, we managed to get the experiments done with high quality. Nevertheless, throughout the time span of this project including the revision, we still went through difficulties such as changing the type and amount of control RNA spike-in due to evolvement of the technology and standard in the field, changing the version of the microfluidic chip because of a new release from Fluidigm, and revising our experimental design to take into account the difference in starting material (mRNA abundance) between different treatments.
Now we have the data! But how should we analyze it? In order to do data analysis, I took a great two-week workshop organized by UNC High Throughput Sequencing Facility that jumpstarted my bioinformatics and enabled some basic analyses by myself. But as we all understand, a two-week workshop and some self-learning are not sufficient for a high-quality paper and we needed a real expert at that time, desperately. So here is the magic how we found our wonderful collaborator, the computer modeling expert and the other co-first author of the paper, Dr. Josh D. Welch . To tell the story, I had to mention Haley Ruth Vaseghi first. Haley is a graduate student in the Qian lab. She knew that we were looking for help with single cell RNA-seq data analysis and her best girlfriend’s husband happened to be an computer modeling expert with a strong interest in analyzing single cell omics data. So she basically connected us by email and said:” Hey, maybe you will want to talk to each other~” Then we found our amazing collaborator Josh, who was still a graduate student in Dr. Jan Prins’s lab in the Computer Science Department at UNC at that time, now a postdoc at the Broad Institute and Assistant Professor-to be at University of Michigan. I knew immediately after our first meeting that Josh was not those graduate students that you say “em…” or ”fine.” He was one of those few graduate students that you say “wow!” His passion, expertise, and sparkling ideas turned out to tremendously help the project. From experimental design to data processing and normalization, then to data modeling and analysis and manuscript writing, his input to this project ensured the quality of data analysis in the paper.
Our wonderful collaborator Josh! & graduate student in the lab, the “matchmaker” Haley!
Back to the Science
With everything ready, we explored the mechanisms of iCM reprogramming using single cell RNA-seq. We analyzed the transcriptome of 513 single mouse neonatal cardiac fibroblasts that undergo reprogramming for 3 days. Combined with computational modeling and experimental validation, here are what we found:
Using unsupervised dimensionality reduction and clustering algorithms, we identified molecularly distinct subpopulations of cells during reprogramming, including an novel intermediate cell population pre-iCM that express both fibroblast (the start cell) and cardiomyocyte (the target cell) genes. Our findings here suggest that iCM reprogramming is different from the induced pluripotent stem cell reprogramming, which requires early down-regulation of fibroblast markers for reprogramming to proceed.
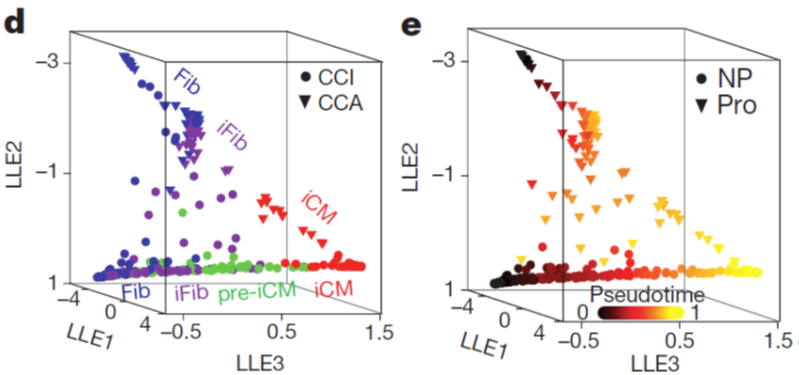
We constructed the route of iCM formation using SLICER, an algorithm for inferring nonlinear cellular trajectories, and calculated pseudotime of reprogramming progress for each cell based on the trajectory. The trajectory revealed a bifurcation of reprogramming and proliferation states of cells.
Figure 2. Single-cell RNA-seq reconstructs iCM reprogramming and identifies intermediate cell populations. Three-dimensional trajectory constructed by SLICER showing hierarchical clustering/PCA cell groups (d) or pseudotime (e). Fib, fibroblast; iFib, intermediate fibroblast; CCI, cell-cycle inactive; CCA, cell-cycle active; LLE, local linear embedding; NP, non-proliferating; Pro, proliferating. Figure adapted from paper: https://www.nature.com/articles/nature24454
We followed up on the trajectory and delineated the relationship between cell proliferation and iCM induction. Our data suggest that decreased proliferation or cell cycle synchronization promote iCM reprogramming and that increased proliferation inhibits reprogramming. This finding could be important for potential clinical application of reprogramming. Because after myocardial infarction, cardiac fibroblasts become activated and proliferative but their proliferation gradually decrease along with time post injury.
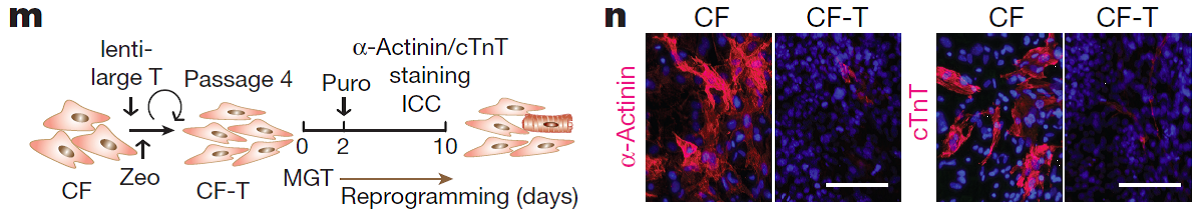
Figure 3. Impact of immortalization of cardiac fibroblasts and hence accelerated proliferation on iCM induction. Schematic for the experiment (m) and fluorescent images showing expression of two cardiac structural proteins cTnT and α-actinin (n). Nuclei were stained with Hoechst. Scale bar = 100 µm. CF, cardiac fibroblast; ICC, immunocytochemistry; puro, puromycin; zeo, zeocin. Figure adapted from paper: https://www.nature.com/articles/nature24454
We performed nonparametric regression and k-medoid clustering to identify clusters of genes that are significantly related to and show similar trends over the reprogramming process. We found immediate and continuous downregulation of genes involved in the basic cellular machineries of protein translation/ biosynthesis, modification and transportation. This is consistent with another observation during the data analysis that total mRNA abundance decreased by 40% upon reprogramming. These changes are probably to balance for increased energy requirements during the cell-fate switch and/or to transit from a protein production and ‘secretion factory’ (a fibroblast) to an energy-consuming ‘power station’ (a cardiomyocyte).
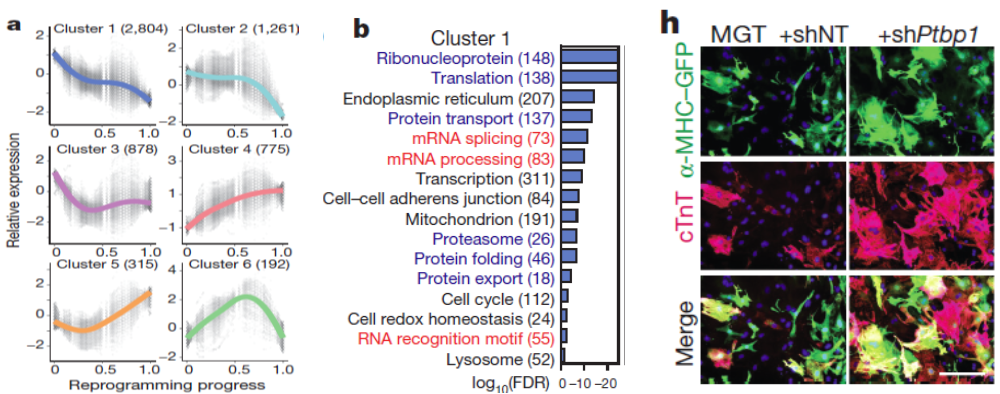
Further analysis of global gene expression changes during reprogramming revealed unexpected downregulation of factors involved in mRNA processing and splicing. We therefore performed a loss-of-function screen against a library of major splicing factors and identified Ptbp1 as the top candidate. Detailed functional analysis revealed that Ptbp1 is a critical barrier for the acquisition of cardiomyocyte-specific splicing patterns in fibroblasts. Concomitantly, Ptbp1 depletion promoted cardiac transcriptome acquisition and increased iCM reprogramming efficiency.
Figure 4. Identification of Ptbp1 as a barrier to iCM splicing repatterning. Six gene clusters were identified to change during reprogramming (a) with gene ontology analysis of cluster 1 shown (b). False discovery rate (FDR) < 0.05. The number of genes in each cluster/ GO term is shown in parentheses. (h) Fluorescent images of cTnT and α-MHC–GFP of MGT-infected cardiac fibroblasts treated with shRNA against Ptbp1 (shPtbp1) or shRNA non-targeting control (shNT). Nuclei were stained with Hoechst. Scale bar = 200 µm. Figure adapted from paper: https://www.nature.com/articles/nature24454
Additional quantitative analysis of our dataset revealed a strong correlation between the expression of each reprogramming factor and the progress of individual cells through the reprogramming process, suggesting that reprogramming is a process highly associated with/determined by the expression levels of reprogramming factors.
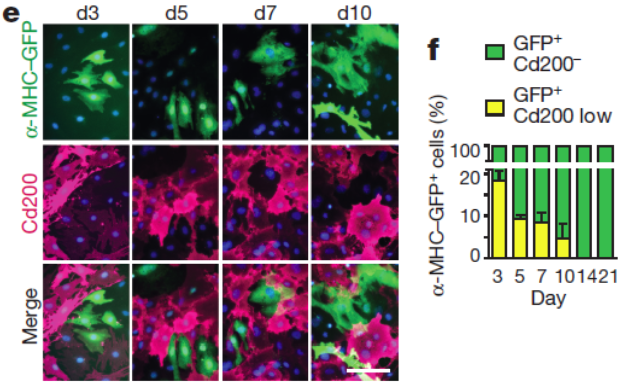
Correlation analysis led to the discovery of new surface markers for the enrichment of iCMs such as the top negative selection marker candidate Cd200. Combinatorial use of these negative selection markers with positive selection cardiac reporters might enable convenient enrichment of iCMs using e.g., FACS sorting.
Figure 5. Negative selection marker of iCM identified from single cell RNA-seq. Fluorescent images (e) and quantification (f) of Cd200 and α-MHC–GFP during reprogramming. n = 20 images. Scale bar = 100 μm. Figure adapted from paper: https://www.nature.com/articles/nature24454
In summary, we used single-cell transcriptomics analysis to gain insights into the heterogeneity of cells within an unsynchronized cardiac reprogramming system. The findings show promise for improving the efficiency and detection of iCM formation. We also anticipate that the experimental and analytical methods presented here, when applied in additional cell programming or reprogramming contexts, will yield crucial insights into cell fate determination and the nature of cell type identity.
Everyone’s Efforts Make Things Happen— the Collaborative Qian Lab
Throughout this project especially during the revision process, everyone in the Qian lab and our collaborators all help a lot. In addition to the other two first authors mentioned above, Dr. Yang Zhou, another postdoc in the lab helped with a lot of the proliferation assays; Dr. Hong Ma helped with most of the Ptbp1-related experiments; Haley, as mentioned above, helped with the shRNA screen; our lab manager Dr. Chaoying Yin helped with cloning and a bunch of other things throughout the project; Dr. Weining Shen in University of California-Irvine, expert in stats and my college classmate helped with the statistical analysis; several other students and volunteers Blake, Sahar, Michael, and Shuo helped with different experiments. The two most important people behind this project are surely Dr. Jiandong Liu and Dr. Li Qian. They are not only a couple of passionate and talented scientists, but also caring mentors always supporting and guiding us on our way of scientific adventure. The most amazing part of my experience in the lab besides the quality and productivity of research is actually the unbelievable collaborative, encouraging, and happy environment Li and Jiandong have created. I think that is the exact reason why things can happen this way and everyone is enjoying both life and research in this lab. I feel blessed for being a part of the lab.
The Qian and Liu lab at the award ceremony of 2017 UNC Postdoc Mentoring Award! All five postdoc in the two labs nominated Dr. Qian together and she became the youngest winner of the award, showing her effort and excellence in mentoring trainees in the lab! Upper row left to right: Sahar, Michael, Blake, Chaoying, and Hong. Lower row left to right: Yang, Li Wang, Ziqing, Li Qian, Jiandong, Haley, Tiffany and Peisen. Here is the lab website: http://uncliqian.web.unc.edu/
Our other wonderful collaborators Dr. Jan Prins at UNC and Dr. Weining Shen at UCI
What Is Next?
Decades of accumulation of bioinformatics and computational modeling algorithms and improvement on sensitivity of each step in the sequencing pipeline finally led to the quantum leap that resulted in the advent of single cell OMICs. It is like the invention of smart phone – a completely new era of biological discovery has dawned. Single cell OMICs provide access to new biological questions or revisits of old biological questions but at an unprecedented resolution. Examples of questions to re-ask include “what defines a cell?”, “ultimately how is the fate of a cell determined?” and “what are molecular cascades to establish the cell fate once determined?”
In the world of big data, changes in technology and computer modeling algorithms are rapidly re-shaping the world around us and the way we do research. Interdisciplinary projects adopting combinatorial approaches of both biological experiments and computational analyses will open new windows of biological discoveries and inspire new ideas on both sides. In the future, we expect to continue this strategy of interdisciplinary collaboration and further explore cell fate control in development, regeneration, and disease.
Interested in exploring cell adhesion, cytoskeletal regulation, or Wnt signaling & their roles in development and oncogenesis, as a graduate student or postdoc? The Peifer lab at UNC-Chapel Hill is a great place to explore these. You can learn more about our work at: http://peiferlab.web.unc.edu and a graphic summary is at: http://peiferlab.web.unc.edu/files/2017/11/PeiferLabPoster2017sm.pdf
I’ll be at the ASCB/EMBO Meeting in December-drop me a line to meet or email at peifer@unc.edu
On August 2013, I took my first one-way trip departing from Puerto Rico. Although I have always been passionate about travelling to as many places (the cheapest way) possible, embracing the PhD-journey in a new city I would have to call “Home” was very intimidating. Close to the start of my journey at the Georgia Institute of Technology I joined the Garcia Lab due to my interest in the engineering of materials to study tissue development and regeneration. A couple more months later and I have already found myself very engaged in a new project in collaboration with the Nusrat Lab. The project involved the engineering of a synthetic hydrogel to direct mouse intestinal stem cell differentiation in vitro and serve as an in vivo cell-delivery vehicle that promotes intestinal wound repair, overcoming the translational limitations associated with natural matrices. This project aimed to provide potential therapeutic options by facilitating intestinal mucosal wound repair which is central in many pathologic states, such as inflammatory bowel disease.
At the time I was very much enjoying the work and, considering my heavy fundamental engineering background, it was all very novel to me as I faced a steep learning curve of very exciting biological concepts. In addition, having such amazing collaboration with cell biologists and clinicians placed me in the forefront of multiple fields that collide for a single goal.
It was all fun and games until I found myself two years later stuck in the same first-stage of my project because the mouse intestinal tissue did not survive within (or basically, hated) my hydrogels. As I continued to assimilate that we weren’t going to reach our desired experimental results (or in colloquial words, that “the project wasn’t working”) we met Jason Spence. He introduced us to the novel technology of in vitro generation of human intestinal organoids (HIOs) from human pluripotent stem cells (hPSCs). We grew to what is now a three-lab interdisciplinary collaboration involving bioengineers, clinicians, cell biologists and developmental biologists. And that’s where this journey took a different turn.
With experiments happening mostly at the University of Michigan I now had the opportunity to merge into another new environment with new collaborators and, just as significant, a different weather. Must say that for a Caribbean islander (despite having such amazing hosts) spending winter in Ann Arbor made the experience quite more challenging. Furthermore, due to the complexity of the hPSC culture I used to travel to Michigan in very short (sometimes even with one-week) notice. Thus, having to figure out the logistics for the trip (from lodging to shipping reagents; from all I was leaving behind in Atlanta to the experimental planning in Michigan) was very challenging. Once I was there, there was no time to waste. The clock was ticking, Miguel and I had an experimental plan to accomplish despite any (or the many) complications we faced and, to me, returning to Atlanta saying, “I didn’t have enough time to do that” was not an option. Nevertheless, after several trips to the beautiful (and cold) Ann Arbor, lots of planning and experimenting, research started to move forward, my project kept evolving and, most importantly, “it was working”…
Ricardo Cruz-Acuña examines differentiating human intestinal organoids (HIOs) under a microscope. The research may lead to a new technique for treating injuries caused by gastrointestinal diseases. (Credit: Rob Felt, Georgia Tech)
The Science
In this project we studied the in vitro generation of human intestinal organoids (HIOs) from human pluripotent stem cells (hPSCs). This technology offers strategies for generating multi-cellular 3D structures that recapitulate important features of the human intestine. For example, HIOs can be used for the establishment of chronic intestinal disease models, such as inflammatory bowel disease (IBD), and provides a platform for functional modeling and repair of genetic defects in intestinal development. As part of the method to generate HIOs, hPSCs must be cultured and differentiated in a MatrigelTM-coated substrate, giving rise to 3D intestinal spheroids which are collected and encapsulated within MatrigelTM for expansion into HIOs. MatrigelTM suffers from lot-to-lot variability and an undefined tumor-derived nature which limits its clinical translational potential. Therefore, we implemented a synthetic hydrogel based on a four-armed, maleimide-terminated poly(ethylene glycol) macromer (PEG-4MAL), developed in the Garcia Lab, as a substitute to MatrigelTM for encapsulation of intestinal spheroids and further generation of HIOs. Our synthetic material offers significant advantages over natural materials due to its well-defined structure, cytocompatibility and minimal toxicity in vivo, overcoming the limitations of MatrigelTM.
In order to generate the HIOs using our synthetic hydrogel, floating intestinal spheroids were collected from the culture plate and encapsulated within PEG-4MAL macromers that were functionalized with adhesive and crosslinking peptide motifs for expansion into HIOs. The controlled stoichiometric incorporation of peptides to the synthetic material allowed me to independently modify the biophysical and biochemical properties of the hydrogel to ultimately identify an engineered material that supports in vitro generation of HIOs from hPSC-derived spheroids without the need of MatrigelTM. Both biophysical and biochemical properties of the synthetic matrix were important to intestinal organoid formation, and allowed me to engineer an optimal formulation that supports intestinal spheroid survival, expansion and epithelial differentiation into HIOs and differentiation into mature intestinal tissue in vivo to similar levels as MatrigelTM (Figure 2).
Figure 2: Human intestinal organoids (HIOs) generated in the hydrogel matrix differentiate into mature intestinal tissue and present main human intestinal cell types, such as enteroendocrine cells (CHGA; red), after transplantation into an animal. (Credit: Miguel Quirós, University of Michigan)
In addition (and probably my favorite part of the project), we established the use of the engineered hydrogel as a delivery vehicle for HIOs to murine intestinal mucosal wounds using a murine colonoscope. The tunable reaction time scales of this material allowed injection of hydrogel liquid precursors and HIOs to mucosal wounds resulting in an in situ polymerized hydrogel that supported localized organoid engraftment and enhanced wound repair. These findings form a basis for the development of HIO-based therapies to treat gastrointestinal diseases in humans involving intestinal epithelial wounds (e.g., IBD).
The (More) Questions
As we have established a synthetic hydrogel to generate hPSC-derived HIOs and treat intestinal injuries, many new questions arise: Can we use this material to study matrix contributions to gastrointestinal disease and repair? Can we use this material as an HIO-delivery vehicle to treat chronic gastrointestinal diseases? Can we establish this material as a platform to generate different types of human organoids? And the journey continues…
The Thanks
Now back in (the warmer weather of) Atlanta, I could go on for about three paragraphs naming everybody and everything that somehow contributed to what is, so far, my major achievement research-wise. Nevertheless, I feel is more than fair to primarily thank my homeland, Puerto Rico, and the University of Puerto Rico at Mayagüez, as they are the foundation of where I am today and to where I am going. I will forever be in debt to my island (that little piece of heaven) and will always hold it in my heart, proudly, wherever I am.
Join invited speakers and showcase your work to celebrate the 30th HGM. We particularly welcome presentations from budding scientists to foster networking and collaboration opportunities.
This 2-day meeting will end with the Peter Thorogood Memorial Lecture by Prof. William Harris.
29-30 January, 2018 – UCL Great Ormond Street Institute of Child Health, London, UK
Abstract submission for oral or poster presentation: 15th December, 2017
Conference Registration: 8th January, 2018
BackgroundtotheMeeting
Following the 1987 BSDB Meeting on “Craniofacial Development” organized by Peter Thorogood and Cheryll Tickle, the organizers and other colleagues (Mark Ferguson, Andrew Lumsden, Gudrun Moore, Alisdair Ivens, Julian Lewis and Gillian Morriss-Kay) decided to start holding annual informal meetings to increase communication between laboratories working on various aspects of the development of the head, to offer the opportunity to young scientists to present their work in a friendly and stimulating environment and promote collaborative research. The group has met annually since 1989, and the initial number of participants, about 20 people, rose very rapidly through word of mouth with more groups asking to be included in the mailing list. Over the last few years there have always been over one hundred people attending the meeting.
A postdoctoral research position is available starting in 2018 with Asst. Prof. Timothy Saunders’ group at the Mechanobiology Institute, Singapore. The Saunders lab has been active since 2013 and studies the fundamental processes shaping organs and tissues during development.
One major area of study in the lab is spatial position – how do developing embryos know where to position boundaries? One mechanism is the morphogen gradient, but, despite significant work over the past 20 years, there is still little reliable data about the dynamics of morphogen gradient formation. We are part of a major five-year grant focused on understanding the dynamics of morphogen gradient formation in Zebrafish embryogenesis. The project is a close collaboration between experimentalists, biophysicists and computational biologists. We are looking for a talented post-doctoral researcher interested in working at the interface of developmental biology and biophysics. In particular, the position will involve extensive imaging and image analysis of the early Zebrafish embryo, including light-sheet microscopy. This offers an exciting opportunity for a dedicated researcher to be part of a genuinely interdisciplinary project that will advance our fundamental understanding of development.
Candidates should have strong experience in at least one of the following, and display a willingness to learn the other: (1) advanced microscopy techniques and imaging developing organisms; and/or (2) experience with image analysis and handling large datasets.
The Saunders’ lab is a young group at the Mechanobiology Institute. This provides an exciting prospect for a motivated post-doc to be involved in developing research directions. The post-doc will also be expected to guide and help the graduate students in the laboratory. More information can be found at: http://labs.mbi.nus.edu.sg/mod/.
Qualifications:
1) A Ph.D in Biophysics, Developmental Biology, Computational Biology, or related subject.
2) At least one first-author paper in English submitted to an international peer-reviewed journal.
3) Experience in: (i) Advanced microscopy; (ii) Image analysis and handling large datasets; and (iii) Imaging developing organisms.
Salary and benefits are commensurable to educational qualifications and working experience of the candidates. Benefits include annual leave, medical and flexi-benefits, etc.
Please submit your curriculum vitae and a statement about your research interests to
Lee lab has been studying on cell specification process using human pluripotent stem cells, particularly nervous system and skeletal muscle cells to study human diseases and to develop new drugs.
We are looking for highly motivated postdoc(s) who have experience on one of the following fields, including Alzheimer’s disease and tau pathology.
Previous experience on human pluripotent stem cells is not required (but basic cell culture experience will be great), and we value more on the ‘non-stem cell’ expertise. Please send your application (CV and three reference contact info) to Gabsang Lee (leelabjob@gmail.com).
Neurodevelopmental disorders are a group of different conditions in which the development of the central nervous system is disturbed. This includes developmental brain dysfunction, which can manifest as neuropsychiatric problems (Autism Spectrum Disorders, schizophrenia, fragile-X syndrome, down syndrome), or impaired motor function, learning, language or non-verbal communication. The generation of the appropriate diversity of neural cell types, their migration to correct sites in the brain and the establishment of precise connectivity with target cells are key developmental processes that may go awry, leading to brain dysfunction and neurological diseases.
During this advanced 3-week course, we will provide participants with a comprehensive theoretical and experimental/practical training on the most advanced methodologies and models in developmental neurobiology, in health and disease. World experts (both seniors and juniors) in the field of neural stem cells, brain patterning, neuronal migration, axon guidance or the genetics of human neurodevelopmental disorders will give stimulating lectures and design mini-projects to be carried out by the students.
This will give students a unique opportunity to become familiar with cutting-edge in vivo gene manipulation methods (electroporation, Crispr/Cas9, genetics…), 3D imaging technologies (SPIM and light sheet microscopy, tissue clearing, two-photon imaging…), time-lapse microscopy and cell tracking, advanced pluripotent stem cell culture (iPSCs, cortical progenitors and neurons derived thereof, organoids, organotypics…).
Deadline: 18 December 2017 (Midnight Brussels time)
The Johnson Lab in the Department of Developmental Biology at Washington University School of Medicine (http://devbio.wustl.edu) is seeking applications for NIH funded Research Scientist positions. Our lab uses multiple model systems to uncover molecular mechanisms of muscle development and disease. We are looking for highly motivated individuals to begin new projects using mouse models and human ES cells to probe mechanisms of muscle disease.
DUTIES:
Assists in the training and development of technicians, acting as a group leader.
Independently generates mouse strains and collects and analyses histology data.
Develops human ES cell differentiation methods using published protocols.
Performs complex analysis/projects according to research protocols, explaining methods and procedures to other technicians.
QUALIFICATIONS:
Master’s degree with 2 years of experience in a laboratory setting (or equivalent combination of education and experience equaling 7 years)
Experience with mouse husbandry and mammalian cell culture
Please send a cover letter, CV, and list of 3 references to anjohnson@wustl.edu
 (No Ratings Yet)
(No Ratings Yet)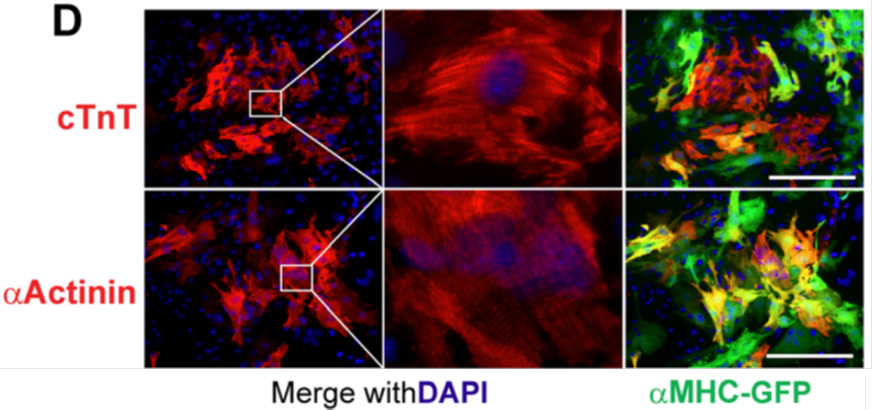




 (5 votes)
(5 votes)