Post doctoral position available to study the genetic and epigenetic control of stem cell attributes and pluripotency, focusing on the neural crest gene regulatory network (NC-GRN). Neural crest cells are stem cell-like progenitors that migrate extensively and whose genesis was central to the evolution of vertebrates. Misregulation of components of the NC-GRN underlies numerous human diseases and congenital disorders. Studies involve post-translational regulation of known network components, and use of proteomics and next generation sequencing to identify novel components.
Highly motivated applicants with a PhD and strong background in cell and molecular biology and/or developmental biology are encourage to apply. Please send a CV, brief description of research interests, and the names of three references to:
Carole LaBonne, PhD (clabonne@northwestern.edu)
Department of Molecular Biosciences
Northwestern University, Evanston, IL 602028
The Fight for Sight charity have provided a three year studentship to Stephen Wilson and Gaia Gestri to support a student to work on the role of the Yap/Taz pathway in the morphogenesis of the eye. The project will use zebrafish as a model and will involve a combination of imaging and molecular genetic approaches to resolve the cell behaviours that regulate eye formation in health and disease.
We are looking for a talented, motivated and enthusiastic student to work in a team of scientists studying various aspects of brain and eye development (www.ucl.ac.uk/zebrafish-group/). Candidates will have suitable MSc and/or BSC qualifications and ideally experience and expertise in imaging and image analysis and in working with zebrafish as a model system.
We are seeking a highly motivated and collaborative postdoc in the area of human embryology and stem cell biology to join Dr. Kathy Niakan’s laboratory.
We have identified several transcription factors and components of key signaling pathways that are highly expressed in pluripotent epiblast cells of the developing human embryo. The pluripotent epiblast has the unique potential to give rise to the entire fetus in vivo and can self-renew indefinitely as embryonic stem cells (hESCs) in vitro. Understanding the molecular basis of lineage specification in the early human embryo is of fundamental biological importance and has significant clinical implications for infertility treatment as well as the use of hESCs to treat various diseases. Importantly, the genes we identified as enriched in human embryos are not expressed in mouse embryos at the equivalent developmental stage, further suggesting differences in pluripotency mechanisms between these species.
The aim of the project is to characterise putative regulators of human pluripotency and embryogenesis using currently the most efficient and precise genome editing technique (CRISPR/Cas9) in human embryos and stem cells. This will provide fundamental insights into human biology and facilitate the development of conditions for the establishment of novel human stem cells. We also seek to establish novel human embryonic stem cells by modulating signaling pathways that we have identified as specific enriched and functional in the development of the pluripotent human epiblast.
The successful candidate is likely to be an energetic, focused, and productive individual with a desire to work in a congenial, dynamic, and collaborative research environment. Good organisational, analytical, and communication skills are essential.
ORGANISATION
Dr Niakan’s laboratory focuses on understanding the mechanisms of lineage specification in human embryos and the derivation of novel human stem cells. Details of research projects currently being undertaken can be seen at: http://www.crick.ac.uk/kathy-niakan
Research techniques used in the laboratory include: molecular biology, advanced microscopy and image quantification, human and mouse preimplantation embryo culture and micromanipulation, genome modification, genome-wide techniques including single-cell RNA-sequencing, human embryonic and induced pluripotent stem cell derivation.
OBJECTIVES
In this project, some of the specific objectives could include, but not be limited to:
Stem cell derivation from embryos
Reprogramming using induced pluripotent stem cell approaches
Genome editing using CRISPR-Cas9
Genomic profiling of early human embryos and microdissected cells
Ensuring the design and implementation of the project
Liaising with collaborators within the Crick, the UK and abroad
Writing and contributing to the preparation of scientific manuscripts, reports, presentations and records of experimental plans and results
Working closely with the Group Leader and other team members to report on the results via publications
Supervising and providing technical advice to more junior members of the team when appropriate
ABOUT US
The Francis Crick Institute has a distinctive vision of how biomedical research is conducted. It is one of the most significant projects in UK biomedical science for a generation. The institute’s labs have an international reputation for cutting edge research into basic biology and are committed to training the next generation of research scientists.
On 1 April 2015, staff from the London Research Institute (CRUK) and National Institute for Medical Research (MRC) transferred to the Crick to form a fully functional research institute on four sites. In 2016, the Crick will move to a single new, purpose built research centre in St. Pancras which will house some 1,500 staff.
PERSON SPECIFICATION
The post holder should embody and demonstrate our core Crick values: Bold, Imaginative, Open, Dynamic and Collegial, in addition to the following:
Essential
PhD in the areas of Developmental Biology, Stem Cells, Molecular Biology or similar (or in the final stages of PhD submission)
Good knowledge and experience in molecular biology and microscopy
Technical expertise in embryo and/or cell culture
Proven track record of research (i.e. publication record)
Excellent communication skills required – both oral and written presentation
Ability to communicate ideas and results effectively and interact fluidly with computational biologists
Ability to work independently and organise own workload
Ability to design experiments, report on research progress and outcomes openly and review methodologies in response to feedback
Highly motivated, organized and analytical
Ability to update knowledge in the specialist area and implement relevant technologies to advance the project
Desirable
Experience in preimplantation mouse or human embryo culture
Experience in human and mouse pluripotent stem cell culture
Experience in preparing samples for advanced sequencing
Experience in genome editing using CRISPR-Cas9 technology
Experience in lentivirus production and transduction
Postdoctoral Training Fellows are expected to lead their own projects, contribute to other projects on a collaborative basis (both in the lab and with external collaborators) and guide PhD students in their research. The ability to work in a team is essential.
If you are interested in applying for this role please apply through our online system:
False colour SEM image of cotton trichomes from different overlapping petals that act like Velcro to hold petals in place as they grow.
Plant hairs or trichomes mean little to most people until they bite into a furry skinned peach or prick their finger on a rose bush thorn, but in the plant kingdom these versatile epidermal structures perform many essential functions that are attributable to their physical shape, location, density and sometimes chemical composition. Next time you pop on a cotton T-shirt think about where the fibres for that textile come from – it’s made from a plant trichome.
For the last 20 years or more our lab has been concerned with understanding the molecular basis for the differentiation of the long hairs or seed trichomes that develop on the seeds of the cotton plant, Gossypium hirsutum L.. These fibres can become very long, reaching up to 5 cm or more in some cotton species, so could be some of the longest cells in the plant kingdom. They are single cells of the cotton ovule epidermis that elongate and fill with a thick secondary cell wall layer that is almost pure cellulose which imparts the special feel and fabric properties of cotton textiles. Using cDNA microarrays we were able to compare the genome wide expression in cotton ovules just initiating the production of fibres and with the gene expression in ovules from a mutant plant that didn’t make any fibres (Wu et al., 2006). This identified members of two specialised classes of plant-specific transcription factors: MIXTA-like MYB genes and homeodomain-leucine zipper IV (HD-zip) genes that were reduced in expression in the fibreless mutant. These two families of genes appear to have evolved diverse functions in regulating epidermal cell differentiation in plants, including plant trichomes.
Cotton seed fibres are long single cells, tens of thousands of which grow on the surface of each seed within the cotton fruit or boll.
Detailed functional analysis of our three candidate genes in transgenic cotton plants using over-expression and gene silencing strategies indicated that the transcription factors MYB25, MYB25-like and HD-1 all played essential roles in cotton fibre development and MYB25 and HD-1 were also key components in the development of other hairs that covered the leaves and stems of the cotton plant (Machado et al., 2009, Walford et al., 2011 and 2012). The sequencing of the cotton genome (Paterson et al., 2012; Zhang et al., 2015) a few years ago (cotton is a polyploid species with two genomes A and D derived from the hybridisation of two ancestral Gossypium diploid species a couple of million years ago) enabled us to look more closely at a whole genome level at all the potential members of these specialised classes of genes. We have focussed mainly on understanding the MIXTA-MYBs. Cotton has ten (including MYB25 and MYB25-like) of these types of genes (twenty if you count both sub-genomes) – seven more than the model plant species Arabidopsis, so there has been an expansion of this gene type in cotton presumably to cope with the large variety of epidermal cell types in cotton, including the seed fibres that are a special feature found in only some plant species!
Over the last few years we have been working our way through the cotton MIXTA-MYB family trying to define their functions using the same approach of characterising their tissue and developmental gene expression patterns and silencing and over-expressing them in transgenic cotton plants. All of them except perhaps MYBML10 (MYB MIXTA-Like gene 10) appeared to be expressed predominantly in cotton ovules during the stages of early fibre development (Bedon et al., 2014). Some of the other genes appear to be part of the regulatory cascade controlling fibre development and are downstream of MYB25-like, but they may also be shared components of development pathways for other types of trichomes. That is another story, but this blog is mainly about MYBML10 that was found to be the master regulator for a very special type of trichome present on the petals of the cotton flower. These trichomes have an interesting evolutionary function and their analysis formed the basis for our recent Letter to Nature Plants (Tan et al., 2016).
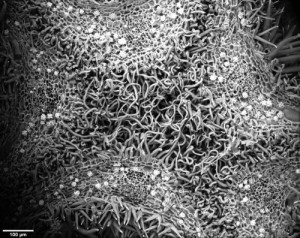
In a phylogeny of the cotton MIXTA-MYB protein sequences, MYBML10 clustered together with MYB25 and MYB25-like, so we expected it to be part of the fibre development pathway, however it was very lowly expressed in whole cotton ovules and in the outer integument layer of the ovule (a 3-4 cell layer that includes the epidermis layer where the fibres initiate and which we can painstakingly dissect away from the rest of the ovule for fine gene expression analysis (Bedon et al., 2013)). When we silenced this gene using RNAi there didn’t appear to be any effect on cotton fibre production, but the sharp eyes of postdoc, Jiafu Tan, noticed that the flower buds of the most silenced lines were abnormal and the seed set on those plants was reduced, mainly because the petals did not cover the anthers and stigma properly when they were very young and parts of these tissues dried out and died. He embarked on a detailed microscopic and molecular analysis to understand why this might be the case and in doing so stumbled on a new type of trichome in cotton that had a pretty important function of holding together the petals of the developing cotton bud to protect the immature reproductive tissues until they were ready to be exposed to the outside environment. Both the outside and inside faces of the cotton petals had trichomes with those on the outside being mostly four-celled stellate or star-shaped trichomes and those on the inside being single celled trichomes. When the petals are folded around each other, as they are in the young flower bud, the outer and inner faces overlap and the two types of trichomes become juxtaposed and entangled just like ‘Velcro’ to lock the petal edges together (false colour picture at top and black and white image below).
SEM picture of a cross-section of a small cotton flower bud (looking up towards the tip) showing how the petals are folded over one another and their trichomes entangled.
Silencing the transcription factor MYBML10 inhibited the development of the trichomes on both faces of the petal and this allowed the petals to slide past one another as they expanded inside the bud causing the bud to twist and expose the young anthers and stigma to the air. As these reproductive tissue were damaged and cotton is a self-pollinating plant, then few seeds were set, so these trichomes were a critical evolutionary development in cotton. By collaborating with a materials science engineer at a neighbouring university’s Engineering Department we were able to take a couple of these petals that were linked together by their trichomes and actuially measure the force needed to pull them apart in a specially designed tension meter – it was a wopping 1 N per petal edge, so these trichomes can withstand amazing forces that must be being generated within the flower bud as the petals expand inside. No wonder the flowers are twisted and abnormal when they don’t have petal trichomes.
Are these petal trichomes something unique to cotton? When we started looking, the answer was a resounding no. Visiting a local nursery (fortunately it was summer), we were able to find a selection of flowering plants to look at. Petal trichomes were by no means found in every flower we looked at, but they were certainly prevalent in the Malvaceae, the plant family that includes cotton and its Hibiscus relatives, but they were also present in a number of other plant families and it wasn’t just on the petals, but also the next outer layer of the flower, the sepals and even the next layer, the bracts.
SEM image of the top of the flower bud of Hibiscus syriacus showing interleaved trichomes from different sepals that hold the bud together as it grows.
Petal trichomes have therefore been an important development during plant evolution, particularly in species, like those in the Malvaceae whose large flowers expand rapidly before the reproductive organs are fully adapted to drier environments. Trichomes have also been found in some other species like tomato to hold together separate anthers into something like a salt shaker (anther cone) to allow the pollen to all be released at one time when an insect visits a flower (Glover et al., 2004) so trichomes are one form of Nature’s glue to effectively bind organs together to create the diversity of structures we appreciate as flowers, and while we didn’t find a new gene that we could manipulate to improve cotton fibre production we did unearth some fascinating biology that illustrates the simple beauty of Nature’s engineering.
References
Bedon F, Ziolkowski L, Osabe K, Venebles I, Machado A and Llewellyn D (2013) Separation of integument and nucellar tissues from cotton ovules (Gossypium hirsutum L.) for both high- and low-throughput molecular applications. BioTechniques54, 44-46.
Bedon F, Ziolkowski L, Walford S-A, Dennis ES and Llewellyn DJ (2014) Members of the MYBMIXTA-like transcription factors may orchestrate the initiation of fibre development in cotton seeds. Frontiers in Plant Science5, e179 doi:10.3389/fpls.2014.00179
Glover BJ, Bunnewell S and Martin C (2004) Convergent evolution within the genus Solanum: the specialised anther cone develops through alternative pathways. Gene331, 1-7.
Machado A, Wu Y, Yang Y, Llewellyn D and DennisES (2009). The MYB transcription factor GhMYB25 regulates early fibre and trichome development. The Plant Journal59, 52-62.
Paterson AH, Wendel JF, Gundlach H, Guo H, Jenkins J, Jin D, Llewellyn D, Showmaker KC, Shu S, Udall J, Yoo M-J, Byers R, Chen W, Doron-Faigenboim A, Duke MV, Gong L, Grimwood J, Grover C, Grupp K, Hu G, Lee T-H, Li J, Lin L, Liu T, Marler BS, Page JT, Roberts AW, Romanel E, Sanders WS, Szadkowski E, Tan X, Tang H, Xu C, Wang J, Wang Z, Zhang D, Zhang L, Ashfari H, Bedon F, Bowers JE, Brubaker CL, Chee PW, Das S, Gingle AR, Haigler CH, Harker D, Hoffmann LV, Hovav R, Jones DC, Lemke C, Mansoor S, ur Rahman M, Rainville LN, Rambani A, Reddy UK, Rong J-K, Saranga Y, Scheffler BE, Scheffler JA, Stelly DM, Triplett BA, Van Deynze A, Vaslin MFS, Waghmare VN, Walford S-A, Wright RJ, Zaki EA, Zhang T, Dennis ES, Mayer KFX, Peterson DG, Rokhsar DS, Wang X and Schmutz J (2012) Repeated polyploidization of Gossypium genomes and the evolution of spinnable fibers. Nature492, 423–427.
Tan J, Walford S-A, Dennis ES and Llewellyn DJ (2016) Trichomes control flower bud shape by linking young petals together. Nature Plants2, Article number: 16093. Published online June 20 2016. http://dx.doi.org/10.1038/nplants.2016.93
Walford SA, Wu YR, Llewellyn DJ and Dennis ES (2012) Epidermal cell differentiation in cotton mediated by the homeodomain leucine zipper gene, GhHD-1. Plant Journal71, 464-478.
Walford S, Wu Y, Llewellyn DJ and Dennis ES (2011) GhMYB25-like: a key factor in early cotton fibre development. The Plant Journal5, 785-797.
Wu Y, Machado A, White RG, Llewellyn DJ and Dennis ES (2006). Expression profiling identifies genes expressed during early lint fibre initiation in cotton. Plant and Cell Physiology47, 107-127.
Zhang T, Hu Y, Jiang W, Fang L, Guan X, Chen J, Zhang J, Saski C, Scheffler BE, Stelly DM, Hulse-Kemp AM, Wan Q, Liu B, Liu C, Wang S, Pan M, Wang Y, Wang D, Ye W, Chang L, Zhang W, Song Q, Kirkbride R, Chen X, Dennis E, Llewellyn DJ, Peterson DG, Thaxton P, Jones DC, Wang Q, Xu X, Zhang H, Wu H, Zhou L, Mei G, Chen S, Tian Y, Xiang D, Li X, Ding J, Zuo Q, Tao L, Liu Y, Li J, Lin Y, Hui Y, Cao Z, Cai C, Zhu X, Jiang Z, Zhou B, Guo W, Li R, and Chen ZJ (2015). Sequencing of allotetraploid cotton (Gossypium hirsutum L. acc. TM-1) provides a resource for fibre improvement. Nature Biotechnology33, 531–537 doi:10.1038/nbt.3207
My informal review of YEN this year is by necessity a bit rushed but, for what it is, here it is. Most reviews are very short and pithy/jealous and only exist for some of the talks where, for a combination of good and bad reasons, I paid attention (I have tried to make this review resemble my actual thoughts on the day, rather than a diplomatic diatribe of the sort I hate). I hope it is as inoffensive as it is informative, but preferably more so.
Session 1: Cell fate determination
Rayon (Crick)
Biology of enhancers driving Cdx2 in the very early mouse embryo. Conventional and nicely done – the kind of work that makes me feel nice. There should be more fundamental work like this.
Laura Hardwick (Cambridge)
I should confess, I read some work on Ngn2 from the Philpott lab as a postdoc working on neurogenesis in the hindbrain and its regulation by bHLH transcription factors, so this is completely biased and self-interested, but I really like this work. Laura’s talk built on an original finding (that I read about in 2011) showing how a single transcription factor could both define proliferating progenitors and drive post-mitotic differentiation in the same embryonic territory, and perhaps, even the same cell lineage. This talk outlined a very thorough suite of biochemical experiments suggesting that post-translational regulation by phosphorylation of bHLH transcription factor activity might actually be a very general mechanism that applies beyond neural tissue. Interesting stuff.
Session 2: Polarity/asymmetry
Vijayakumar (KCL)
This was a really cool talk given by a very good speaker. What was really nice is that she had the grace to attribute credit elsewhere in relation to a lovely project started by someone I know, who always underestimated their work. The finding that playing around with the ECM can significantly affect the location and distribution of neural proliferation always struck me in informal pub conversations as really cool (seriously, proliferation at the basal as well as apical surface?). Add in some high end video microscopy of these progenitors in real time and this will be a very compelling phenotype. I look forward to reading about it.
Faro (UCL)
Characterisation of an insertion mutation that causes an asymmetric brain defect. Turns out Wnt signalling drives asymmetric patterns of neurogenesis in the habenula (posterior forebrain). A potentially boring positional cloning project turned into something actually very interesting.
Fols (F1000)
Obligatory F1000 talk at a conference. The business model is that ‘the best way to read science is to start with the opinions of a small group of people educated in the 70s/80s’. Ironically quite forward-looking though with post-publication review as standard.
‘Guest’ session: Human and mouse embryonic development
Niakan (Crick)
Human embryos, the Crick Institute, oodles of newspaper coverage. Everything that in this day and age counts as good science. Turns out that early human development is very different to mouse: early development is very plastic during evolution. Who knew?* Still, ended by emphasising how important understanding a diversity of animal species was (evo-devo anyone?). I am so spineless. I loved it.
Srinivas (Oxford)
This guy is awesome, though I got rather lost. Anterior visceral endoderm migration with super cool in vivo live imaging. If only I liked morphogenesis…
Plenary
Martin (Bristol)
Sometimes speakers rather misjudge (or don’t care about) the purpose of YEN. Sometimes senior PIs get a platform that they didn’t need to publicise work that is already famous. Not here. Just right. Self-deprecating and quality in equal measure (‘in those days you didn’t need 15 Cell papers to get a job, you just went over the road and asked for one’). Just about hilariously, he also spoke about penis development (‘if you don’t have one to look at, you can ask a friend’). In all seriousness though, I feel terrible about the way that everyone seems to be, or even worse, actually is, obsessed with the way that research funders have become obsessed with translation or ‘economic impact’. It is actually sad that so many PIs pay lip service to whatever strategic priority ResearchUK (or whatever they are called) come up with next. However, Paul Martin’s work is genuinely very very impressive (this is like one of those facebook posts where an erstwhile sensible and suitably cynical friend posts about how much they love their new boyfriend/house/dog. I always write ‘vomit’. React to this as you will.): studying a fundamental biological processes of cellular behaviour (albeit behaviour of very ‘translational’ cells – macrophages) leads to super-significant insights into both wound healing and cancer, while at the same time being interesting for its own sake. A really nice combination of in vivo imaging-based phenomenology and mechanistic dissection of the underlying processes.
Two HFSP-funded postdoctoral positions (each for a duration of 3 years) are available from 1 December 2016 to study the development, robustness and evolution of QR neuroblast migration in C. elegans and related species, in the laboratories of Hendrik Korswagen at the Hubrecht Institute, The Netherlands and of Marie-Anne Félix at the Ecole Normale Supérieure, Paris, France. The collaborative project is based on the recent finding by the Korswagen group that this cell migration is controlled through the timed expression of a Wnt receptor (see Mentink et al. Dev Cell 2014 for more information) and an important aim of the project is to understand how such precise timing of gene expression is achieved.
If you are interested in one of these positions, please send a CV, a statement of research accomplishments and interests, and the contact information of three references to
Model Organisms such as yeast, worm, fly, fish, rat, and mouse are key drivers of biological research, providing manipulable and cost-effective experimental systems that continuously yield fundamental insights into human biology and health. These discoveries rely on the accumulated wealth of genetic, genomic and cellular knowledge for each organism, which is made accessible via the Model Organism Databases (MODs). Foundational and consistent funding of the MODs by the National Institutes of Health (NIH) enables tens of thousands of researchers throughout the United States, as well as from the international community, to uncover basic, conserved biological mechanisms relevant to new medical therapies. These discoveries have been recognized by many Nobel Prizes over the last two decades alone.
NHGRI/NIH has recently advanced a plan in which the MODs will be integrated into a single combined database, along with a 30% reduction in funding for each MOD (see also these Nature and Science news stories). While increased integration will present many advantages, the plan will result in a loss of critical organism-specific datasets. The funding cut will also cripple core functions such as high quality literature curation and genome annotation, degrading the utility of the MODs. Given the large number of scientists that this policy change would affect and the importance of their work, this is a matter of extreme concern.
Leaders of several Model Organism communities, working with the Genetics Society of America (GSA), have come together to write a Statement of Support for the MODs, and to urge NIH to revise its proposal. All scientists who value the community-specific nature of the MODs are being asked to sign this open Letter of Support.
Your name will be added to those of the Charter Signatories that includes Model Organism Community Leaders, the Genetics Society of America, American Society for Cellular Biology, and the Society for Developmental Biology, plus a dozen Nobel Laureates, and many other scientists (in fact, we have 6524 signatures as of June 23rd!).
The letter, along with all signatures, will be presented to NIH Director Francis Collins at a GSA-organized meeting on July 14th 2016 during The Allied Genetics Conference in Orlando. We urge you to add your name to the Letter of Support, spread the word to all researchers who value the MODs, and let YOUR voice be heard.
This content was originally posted at: http://www.genetics-gsa.org/MODSupport/
Developing Future Biologists (DFB) is a student-led organization at University of Michigan dedicated to ensure that the next generation of biologists regardless of race, gender, or socioeconomic status have the opportunity to learn the core concepts of developmental biology. During the last week of May, DFB held its 2nd successful week-long short course aiming to inform students from diverse backgrounds about developmental biology and career opportunities in science.
DFB is a graduate student-founded and led initiative that seeks to introduce the core concepts of developmental biology to students from a diverse set of backgrounds. The idea of bringing this course to Puerto Rico was conceived by a former University of Michigan (UM) graduate student, Andrea Ramos, who attended the University Puerto Rico at Ponce for her undergraduate studies. With four additional UM graduate students and three faculty members planning began and within a year the first iteration of DFB was held in Ponce.
As she carefully peeled away the shell of yet another unfertilized chicken egg, she was quickly learning that science requires persistence. After 10 unsuccessful attempts, this young scientist gently cut into her last egg to unveil a tiny, beating heart. Success! For an entire week, the graduate student instructors of DFB, watched stories such as this one unfold, justifying the months of design and practice to make the course a reality.
This year, thanks to funding from sources such as The Society for Developmental Biology and a number of sources at the University of Michigan, the course was able to extend its reach to undergraduate students from different campuses of the University of Puerto Rico across the island. Throughout the week, 24 Puerto Rican undergraduate students were exposed to various key topics in developmental biology through lectures and hands on lab activities using different model organisms and research techniques.
Our developmental biology course is designed to ensure that students understand how an organism develops from a single cell to a fully developed organism, and what happens when developmental processes are disrupted. During the week we covered topics such as: vertebrate embryogenesis, cell signaling, gene expression, organogenesis, and developmental defects. The lectures led by UM faculty and laboratory sessions led by UM graduate students are highly interactive in order to capture the interest and enthusiasm of the students. For example, the gene expression lecture included a Wnt dance and labs included activities ranging from staging Xenopus embryos to observing limb structures in various mouse mutants.
Learning about developmental biology was however only half of what we aimed to achieve. Another component of our program is based in establishing long term mentoring relationships with the students to help them learn about careers and educational opportunities in science. This year students were also able to attend sessions on career development topics including mock interviewing, a CV workshop, a panel discussion on grad school, and a session on presentation design. In addition, to these informative sessions, there were also several social events throughout the week, such as an ice cream social, and a bowling night, where the graduate student instructors from UM and the DFB students could network.
Currently the mentoring from both the first and second iterations of DFB has continued, with some students already inquiring about application formatting and some alumni from the first DFB attending summer programs at various school in the states including UM. In the future, DFB aims to organize a local iteration of the course in the Detroit and at other institutions with underrepresented minorities. The group further aspires to continue having the weeklong course at the end of May in Puerto Rico.
However, whatever the future of DFB, one thing is for certain: the memories and excitement made thought the week will never be forgotten, and lives on through the mentoring and networking that is still ongoing between the DFB alumni and the graduate student instructor of the course.
Article Contributors: Eden A. Dulka, Martha Echevarria Andino, Samhitha Raj, & David Lorberbaum of the 2016 DFB Team
My name is Martin Minařík and I am a PhD student in Robert Cerny’s lab at Charles University in Prague, Czech Republic. Our lab focuses mostly on the development of non-teleost fishes, namely bichirs, sturgeons, and gars. The advantage of having these animals as model organisms is that their breeding seasons alternate throughout the year, so that we can always focus on one species at a time. Thus, while bichirs and sturgeons keep us busy during winter and spring, respectively, the gar season spans throughout the summer.
Anaesthetized female of the Mexican gar (Atractosteus tropicus). Photo: M. Minařík
Unlike zebrafish and medaka, gars did not undergo the teleost-specific genome duplication, rendering them a suitable model for approximating the ancestral ray-finned genome. The same holds true for a variety of phenotypic traits that have been lost in teleosts, ranging from the amphibian-like mode of gastrulation through the thick, enamel-covered ganoid scales to the asymmetric tailfin. Recent years have therefore seen a revival of gar research, with the genome of the spotted gar (Lepisosteus oculatus) being available since 2011. While the spotted gar is reasonably accessible both in the wild and in the pet trade, it has rather slow generation time with females starting to spawn in their fourth year of life. The tropical gar (Atractosteus tropicus), on the other hand, reaches sexual maturity within the first (males) or second (females) year. Moreover, for some reason the tropical gar meat is popular in one particular place in the world – the swampy state of Tabasco, Mexico. The local aquaculture facilities produce millions of eggs each year to sustain the wild stocks thriving in the lakes around the city of Villahermosa, making it the ideal place to reach vast quantities of embryos suitable for wide variety of experiments.
The fact that gars are being bred down in the Mexican tropics is a mixed blessing. Of course travelling each summer to the land of cocoa and iguanas is extremely exciting for a Central European. On the other hand, it brings some difficulties, too. The first is the working temperature, with the spawning room heated up to 40°C. It gets better in the lab, where the temperature is set to 28°C, which represents a compromise between the long-term survival of the researcher and the embryos.
To induce spawning, both males and females are injected with hormones (here under the kind supervision of Lenin Arias Rodriguez). Photo: R. Cerny
The typical day starts in the hatchery by injecting the fishes with hormones. Once spawning behaviour is detected in females, they have to be anaesthetized and spawned manually to avoid releasing the adhesive eggs to the artificial vegetation. It means that sometimes we have to wake up every hour or two during the following night and check the tanks. This gets particularly annoying at the beginning of rainy season, as the ten-metre passage from the lab building to the hatchery is enough to turn your skin into a mosquito banquet. Nowadays this is especially unpleasant, with dengue, chikungunya and zika hiding in mosquitoes’ salivary glands (a thick layer of repellent helps, but one has to be careful to not contaminate the tanks).
It is hard to wake up at 4 a.m. if you know that you have to spend next few hours sweltering in the hatchery. Photo: M. Minařík
One of the local BSc. students, Adriana, injecting DiI to gar embryos. Photo: M. Minařík
Once we get the embryos, the work paces up. We have 24 hours until the neurula stage; the experiments have to be set up and the embryos dechorionated, which is kind of tricky in this fish. The outer membrane is soft and sticky, and it tends to bend inwards when broken so that it adheres to the embryo and tears it apart. To prevent this, one can steal some dried milk from the office and wash the eggs in a milky water to saturate the adhesive layer. Lenin, the lab PI, helps us a lot to get things ready on time. The easiest thing to do in a lab which is not equipped for a serious embryological work is to soak the embryos in selected inhibitors of key signalling pathways. They can also be easily injected with cell tracking dyes – there is no need for micromanipulator as the eggs are double the diameter of Xenopus’s. But if you are loading your needle from a drop of DiI in a saccharose solution, be careful to prevent ants from finding it. The smallest ones, called lelitas in Villahermosa, are difficult to notice, but when they spot the piece of parafilm, they immediately form a swarming highway to the sweet, fluorescent drop. You can run out of your cell tracking dye really quickly this way!
Whereas the neural folds are flat and incospicuous in the gar, in the early tailbud stage the embryo already protrudes well above the yolk sac. The large archenteric cavity can be visualised in this stage by injecting ink (heads point to the left). Photo: M. Minařík
Self-staining fluorescent ants are among the most common companions in the lab. Photo: M. Minařík
For someone who is used to working with teleosts, the gar eggs are really different and fascinating. The embryo is sitting on a huge, empty archenteric cavity, which you can easily inject and visualise, simply with pen ink to start with. Soon it starts to develop prominent attachment glands which the larvae use to adhere to the surface of the water. They tend to aggregate in thousands, forming rafts in the tanks spanning tens of centimeters in diameter – it might sound counterintuitive, but their yolk is fairly poisonous so the clusters of larvae are probably less tempting for predators than it might seem at first glance. As the development proceeds, they grow large opercula, which make them look like tiny umbrellas. Living in extremely warm waters low in oxygen, they use this to generate a flow of water to the gills.
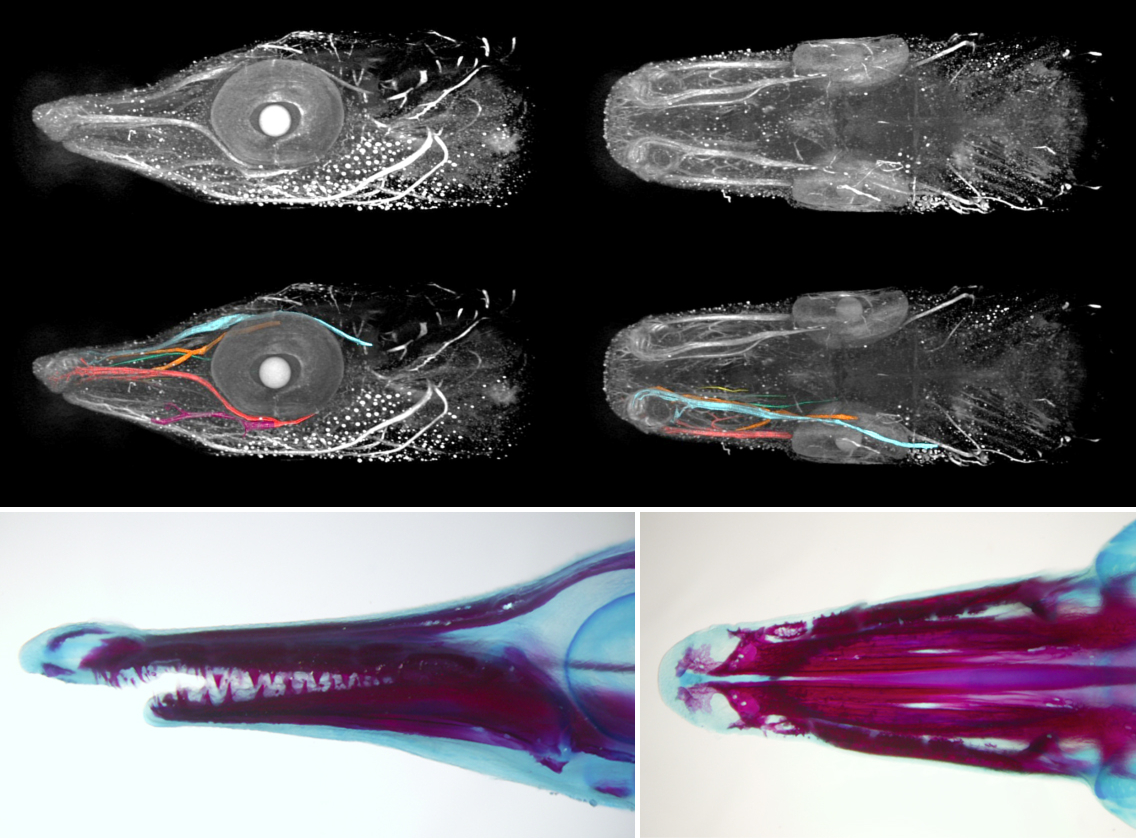
When the larvae hatch, they tend to aggregate below the surface of the water using their cement glands. The gland shows an extensive innervation by a branch of the trigeminal nerve (left). The large opercula (op) develop in later stages (middle, right) to facilitate breathing in the low-oxygenated water. Photo: M. Minařík
The adult gars are kept in outdoor tanks together with tilapias on which they feed occasionally. The largest tilapias, however, are too big for them to catch, so from time to time it is necessary to eliminate them. The best way apparently is to make a tilapia barbecue. With loads of chilli and limes, of course. Meanwhile, our embryos develop to the desired stage. Thanks to the high temperatures it takes only four days until they hatch in the lab, or two days in the hatchery, respectively. Unfortunately, we have to wait with the analyses until we get back to Prague with our samples, so it might be hard to adjust the experiments sometimes, as we can only have a superficial look at the results in the aquaculture lab.However, working in tropical gar in the middle of Mexican swamps is indeed worth all the difficulties.
Back in Prague we can use variety of techniques to further analyse the samples, ranging from microCT imaging (top) through ISH and immunohistochemistry to alizarin staining (bottom). Photo: M. Minařík & B. D. Metscher, A. Pospíšilová
Finding ways to combine the study of gar embryology with the culture of keeping this traditional fish species in Mexican aquaculture might be challenging sometimes, but the exchange of skills and ideas is beneficial for both parties. It is so refreshing to get out of the lab in Prague and jump into the warm fish tanks in Villahermosa to catch fishes for your own experiments! And as the collaboration tightens, we have also our first Mexican students visiting Prague, using sturgeon embryos to adopt techniques they could later use in their own gar projects. So hopefully this intercultural and interspecific enrichment will successfully continue.
In the outdoor tank, trying to catch some fresh gar males (and tilapias for the barbecue). Photo: R. Cerny
The whole process of the artificial fertilization in the Mexican gar in two minutes.
This post is part of a series on a day in the life of developmental biology labs working on different model organisms. You can read the introduction to the series here and read other posts in this series here.
Epithelial Cell and Developmental Biology – Mechanotransduction at tight junctions
Applications are invited for the posts of two postdoctoral research fellows. One will be based in the Laboratory for Epithelial Cell Biology (with Karl Matter and Maria Balda) of the UCL Institute of Ophthalmology and the other in the Zebrafish Group of the Department of Cell & Developmental Biology (with Masazumi Tada).
The successful applicants will join a collaborative research programme that focuses on the role of mechanotransduction at epithelial tight junctions in epithelial differentiation and dynamics, and early development. The aim is to analyze the molecular mechanisms of how tension at tight junctions is transmitted between adhesion proteins and the cytoskeleton, and the roles such processes play during epithelial cell dynamics and differentiation in vitro and during zebrafish gastrulation. (J Cell Biol 208, 821-838, 2015; J Cell Biol 204, 111-127, 2014; Nat Cell Biol 13, 150-166, 2011; Development 136, 383-392, 2009; Development 139, 3897-3904, 2012; Nat Rev Mol Cell Biol 2016).
The positions are funded by the BBSRC and require a PhD in Cell/Molecular/ Developmental Biology, Biophysics, or another related discipline. The persons appointed will have experience in developmental biology, biophysical approaches, molecular cell biology and/or advanced microscopy. Good communication skills and interest in collaborative research are essential.
Informal enquiries about the posts may be made to Karl Matter (email: k.matter@ucl.ac.uk) or Masazumi Tada (m.tada@ucl.ac.uk).
Applications can be be submitted at:
http://www.ucl.ac.uk/hr/jobs/
Ref No: 1559085 and 1557441
Closing date: July 15, 2016
 (No Ratings Yet)
(No Ratings Yet)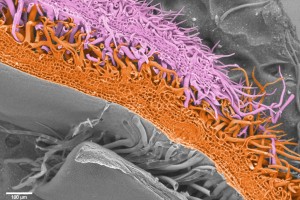


 (6 votes)
(6 votes)





 (11 votes)
(11 votes)







 day in the life of developmental biology labs working on different model organisms. You can read the introduction to the series
day in the life of developmental biology labs working on different model organisms. You can read the introduction to the series