Stem cells have the unique properties of both self renewal and differentiation into various lineages. In the last few years, chromatin architecture and noncoding RNAs are emerging as major players in both gene regulation and stem cell differentiation. This November, Abcam is hosting a conference in Sicily to promote this exciting field, and has teamed up with the Node to offer the chance to be the conference’s official Reporter, with free registration!
Are you fascinated by chromatin organization, transcriptional regulation and stem cell biology? Do you want the opportunity to attend a world class meeting in a beautiful setting, to promote your own work and meet leaders in the field? Are you a keen communicator of scientific research?
The Reporter will provide regular updates from the conference via Twitter, and write a meeting report to be published on the Node and Abcam’s website.
Applying is easy: just send a short paragraph (max. 200 words) to events@abcam.com, letting let us know why you think you are the ideal candidate. The winner will receive free registration to the meeting (travel and accommodation not included).
Application deadline: August 1st
Meeting information
Chairs
Luciano Di Croce & Danny Reinberg
Confirmed speakers
Salvador Aznar Benitah, Shelley Berger, Giacomo Cavalli, Victor Corces, Amanda Fisher, Xiang-Dong Fu, Kristian Helin, Taekyung Kim, Robert Kingston, Tony Kouzarides, Erez Lieberman Aiden, Diego Pasini, Ramin Shiekhattar, Ali Shilatifard, Alexander Tarakhovsky, Maria Elena Torres Padilla and Kenneth Zaret.
Marcos Simoes-Costa, Department of Molecular Biology and Genetics, Cornell University
Marianne Bronner, Division of Biology and Biological Engineering, California Institute of Technology
In the 19th century, most embryologists (i.e. precursors to developmental biologists) accepted the germ layer theory – the idea that embryos are composed of distinct layers (ectoderm, endoderm and mesoderm), which will give rise to specific tissues and cell types in the adult. For instance, it was believed that all skeletal components of the body were derived from the mesoderm, whereas the ectoderm differentiated into the nervous system and epidermis. Around the turn of the 20th century, Julia Platt discovered a remarkable exception to this. She used salamander embryos to track neural crest cells, which are an ectodermal progenitor cell population, and observed that they gave rise to skeletal elements of the head (Platt, 1883). These findings were so controversial at the time that Platt was unable to get a faculty position, quit science, and became a politician. But a hundred and twenty years have passed since Platt’s results were published, and time has been kind to her legacy. Today, we know with certainty that the neural crest is indeed an exceptional cell type. During early formation of the central nervous system, neural crest cells delaminate from the neural tube, engage in extensive migration and give rise to a large portion of craniofacial skeleton as well as several other tissues.
Cranial neural crest cells of a chick embryo. Neural crest cells (purple/light blue) can be seen migrating away from the central nervous system (red); anterior pole of the embryo is facing right.
This particular question – how ectodermal cells can differentiate into skeletal tissue– captured my imagination and drove my interest toward neural crest biology. Not that this was the only remarkable feature of neural crest cells: they have the ability to undergo epithelial to mesenchymal transition, are migratory, and can to differentiate into multiple cell types. While neural crest cells emerge from the central nervous system along the length of the body axis, only the cranial crest can give rise to the cartilage and bone of the face. This was elegantly demonstrated by Nicole Le Douarin in the 1960s and -70s through the use of chicken-quail chimeras (reviewed in Le Douarin, 1982). Le Douarin and colleagues showed that trunk neural crest cells are unable to form cartilage even if grafted to the head. Strikingly, cranial neural crest grafted to the trunk gave rise to ectopic cartilage nodules, indicating that these cells are autonomously capable of differentiating into mesenchymal tissue. These results demonstrated that the cranial neural crest was intrinsically different from the other crest subpopulations.
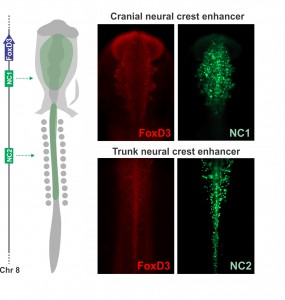
Joining Marianne Bronner’s lab at Caltech after finishing my Ph.D. in Brazil, I wanted to learn molecular biology and genomics. The lab is very interested in gene regulatory control of neural crest specification, so my first project, in collaboration with Tatjana Sauka-Spengler and Sonja Mckeown, focused on cis-regulatory control of the transcription factor FoxD3 in chick embryos. FoxD3 is a classic neural crest marker expressed by all neural crest cells along the entire body axis. However, we discovered different enhancers that drove FoxD3 expression in the head versus the trunk: a distal regulatory element (NC2) mediated trunk expression whereas a unique cranial-specific enhancer closer to the FoxD3 promoter (NC1) drove expression in the head. This suggested that axial differences were encoded in the genome (Simoes-Costa et al., 2012), bringing me right back to the question of what makes the cranial neural crest unique. Importantly, our identification of a axial-specific neural crest enhancers provided a new tool for exploring the molecular mechanisms that underlie the development of the cranial crest.
Axial specific enhancers control FoxD3 expression on the neural crest. The NC1 enhancer is active in the cranial region, while NC2 drives reporter expression in trunk neural crest cells. Adapted from Simoes-Costa et al., 2012.
The strategy to investigate the molecular basis of the cranial neural crest’s ability to give rise to cartilage emerged gradually during daily meetings with Marianne, mostly at the coffee machine. During these informal conversations, we delineated an experimental approach to identify factors unique to the cranial crest that drove differentiation into chondrocytes. Marianne had been interested in this question for a long time, and for more personal reasons – while recovering from a bicycle accident (face plant resulting in a broken nose and deviated septum), she learned from the craniofacial surgeon that facial cartilage is very difficult to replace. We decided to employ a combination of comparative transcriptomics, gene regulatory analysis, and classical embryology to identify the molecular players that endow the cranial neural crest with its ability to form cartilage. Crucial to this approach was the fact that the trunk neural crest is unable to make cartilage in vivo. We predicted that by comparing the transcriptomes of the cranial to the trunk populations – that now could be labelled and isolated with the NC1 and NC2 enhancers – we would be able to identify the transcriptional circuitry that activates chondrocytic differentiation in cranial neural crest cells.
Previously, only a couple factors had been identified as being specific to the cranial crest. Id2 was found to be a cranial neural crest gene by a differential display screen performed by Brad Martinsen in Marianne’s lab almost twenty years ago (Martinsen and Bronner-Fraser, 1998). Eric Theveneau and Muriel Altabef also had identified Ets1 as an important player in the development of this cell population (Theveneau et al, 2007). Our comparative transcriptome analysis identified multiple transcription factors that are enriched in the cranial crest, and we proceeded to organize them in a cranial-specific neural crest regulatory circuit. By analyzing their expression patterns during early embryonic development, we defined the hierarchy of the circuit; and by methodically inactivating each gene, we identified the links that integrated these multiple components into a regulatory network.
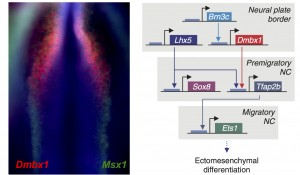
Left: Co-localization of expression patterns from a novel cranial specific gene (Dmbx1) and a pan neural crest marker (Msx1) in a neurulating chick embryo. Right: Cranial neural crest genes regulate each other and are part of a cranial-specific regulatory circuit.
We were very excited by the identification of this novel cranial-specific regulatory circuit, as it provided important insights about the biology of the cranial neural crest. For instance, it showed that expression of cranial neural crest genes depends upon anterior regulatory information that is laid out during gastrulation. Brn3c, Lhx5, and Dmbx1, which are the components of the first “early” level of the circuit, are all co-expressed adjacent to the anterior neural plate in the early gastrula. These genes are required for the expression of the “later” genes – Sox8, Tfap2b and Ets1 – which are retained in the migratory cranial neural crest. However, one fundamental question remained – could this regulatory circuit be important for the cranial neural crest’s potential to give rise to cartilage and bone?
During our coffee break conversations, Marianne and I came to the conclusion that the best way to tackle this question would be to transplant components of the cranial circuit to trunk cells and see if they would gain the ability to switch fates and differentiate into skeletal tissue. Thus, we decided to revisit the classical experiments of Nicole Le Douarin, but with a twist – we would transfect the trunk neural crest from the donor embryos with components of the cranial circuit before grafting them to the head. We would also trade the quail embryos in favor of transgenic GFP+ chicken embryos, to facilitate tracking of the grafted cells. After surgery, we would let our chimeras grow for a week and verify if the reprogrammed trunk crest differentiated into facial skeleton. At that time, I was eager for a foray into classic cut and paste embryology, thinking that perhaps I would spend a month performing surgeries and a couple of weeks analyzing the chimeric embryos.
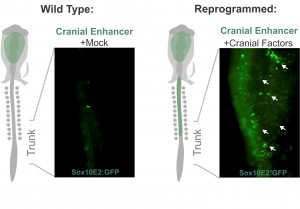
Despite my optimism, that was not the case. Grafting is an art and the chimeric embryos are unforgiving of shaky hands. In my daily meetings with Marianne, I was now drinking decaffeinated tea. Since the grafting experiments were taking an enormous amount of time, we realized we needed a more efficient strategy to identify which components of the cranial circuit were able to reprogram the trunk neural crest. And thus, we came up with the idea of using neural crest axial specific enhancers as reporters of cranial identity. The Sox10E2 enhancer from the Sox10 gene, which was identified by Paola Betancur in Marianne’s lab (Betancur et al., 2010), was perfect for this since it is active in migratory cranial neural crest cells and inactive in the trunk. Thus, we proceeded to transfect trunk cells with all different combinations of the cranial circuit components attempting to activate the cranial enhancer in the trunk neural crest.
Altering neural crest identity: mis-expression of cranial genes in trunk neural crest cells results in ectopic activity of a cranial neural crest enhancer.
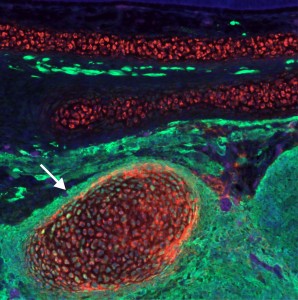
These experiments lead to the discovery that Sox8, Tfap2b and Ets1 are able turn on the cranial enhancer in the trunk and also to increase expression levels of endogenous cranial genes in these neural crest cells. When the reprogrammed trunk neural crest cells were grafted in the head of a host embryo, they not only gave rise to the normal trunk neural crest cell derivatives, melanocytes and neurons, but these grafts also differentiated into chondrocytes, forming conspicuous cartilage nodules. And thus we had demonstrated that we could use information decoded through gene regulatory studies to reprogram neural crest identity and change its fate in vivo.
I am thrilled with the publication of this paper not only because it represents a great deal of hard work, but also because it answers a long-standing question that has interested me for years. The aspect of the work that I like the most is the marriage between old and new – our results highlight the power of classical embryological approaches, especially when used in conjunction with modern techniques. Moving forward, we will employ these findings to devise novel strategies for in vitro differentiation of facial cartilage. It is our hope that regulatory information derived from embryonic development can inform upon novel protocols for cell and tissue engineering, improving strategies for repair and regeneration.
Reprogrammed trunk neural crest cells acquire the ability to form cartilage. The image shows a cross section of a chimeric embryo in which trunk neural crest cells (green) differentiated into ectopic cartilage nodules (arrow) in the craniofacial region.
Transformation of a cell into a cancer cell is a complex process along which the cell acquires hallmarks as diverse as resistance to differentiation, infinite proliferative potential, metastasis, resistance to cell death, genome instability (Hanahan and Weinberg, 2011). Moreover, tumours are usually composed of a heterogenous population of cells which may not contribute equally to the tumorigenic process. Indeed, a hierarchy exists among the tumour cells according to their potential to support tumour growth. In order to decipher the mechanisms governing the transformation process, Drosophila has been shown to be a relevant model. In 2005, Caussinus and Gonzalez elegantly demonstrated that impairing neural stem cell asymmetric division in the Drosophila larval brain triggers their amplification and the formation of large neural tumours (Caussinus and Gonzalez, 2005). Moreover, when transplanted into adult Drosophila hosts for long-term in vivo cultivation, the larval brain tumours displayed very quickly several hallmarks of cancer such as unlimited proliferative potential, genome instability or metastatic activity. In our study, we decided to use this model to determine the mechanisms contributing to the malignant transformation. Caussinus and Gonzalez impaired asymmetric neural stem cell division in a large population of neural stem cells in the Drosophila larval central nervous system (CNS). This manipulation leads to the death of the fly before it reaches adulthood. We bypassed this issue by altering only 6 neural stem cells within the larval CNS using a specific promoter. That way the Drosophila is able to survive metamorphosis and enters adulthood. In the CNS of these adults, we observe huge neural stem cell tumours expanding over and over. This observation is particularly interesting given that wild type neural stem cells possess a limited mitotic potential and terminally differentiate at the end of development, being therefore no longer present in the adult CNS. Thus, neural stem cells in tumours acquire another hallmark of cancer during metamorphosis: resistance to differentiation.
Neural stem cell tumour in the Drosophila adult central nervous system stained for GFP (grey), Chinmo (red) and Imp (blue). The tumour is composed of an heterogenous cell population
Time matters
To terminally differentiate during metamorphosis, the neural stem cell has to journey through a whole developmental program. This program is composed by sequentially expressed transcription factors known as the temporal series. They form an internal clock that governs the identity and the number of neurons generated by each neural stem cell during specific developmental windows. In Drosophila, this neural stem cell-intrinsic clock has been intensively described, particularly, during embryonic and early larval development (Isshiki et al., 2001; Maurange et al., 2008). When the progression in the temporal series is blocked (either by overexpressing or mutating one of the known temporal transcription factors), the neural stem cell stays stuck in this temporal identity and is unable to terminally differentiate during metamorphosis. As a consequence, the neural stem cell persists in the adult CNS. Given that our model of neural stem cell tumour is resistant to differentiation and that temporal series alteration prevents neural stem cells from differentiating during metamorphosis, we wondered whether temporal patterning may be altered in our tumour model.
None of the known temporal transcription factors is relevantly expressed in our tumours at the end of the development demonstrating that the temporal series is not blocked in this context. Nevertheless a target of this early temporal patterning system, the ZBTB transcription factor Chinmo (Maurange et al., 2008), is expressed in a subset of tumour cells suggesting that some cells could have retained an early temporal identity.
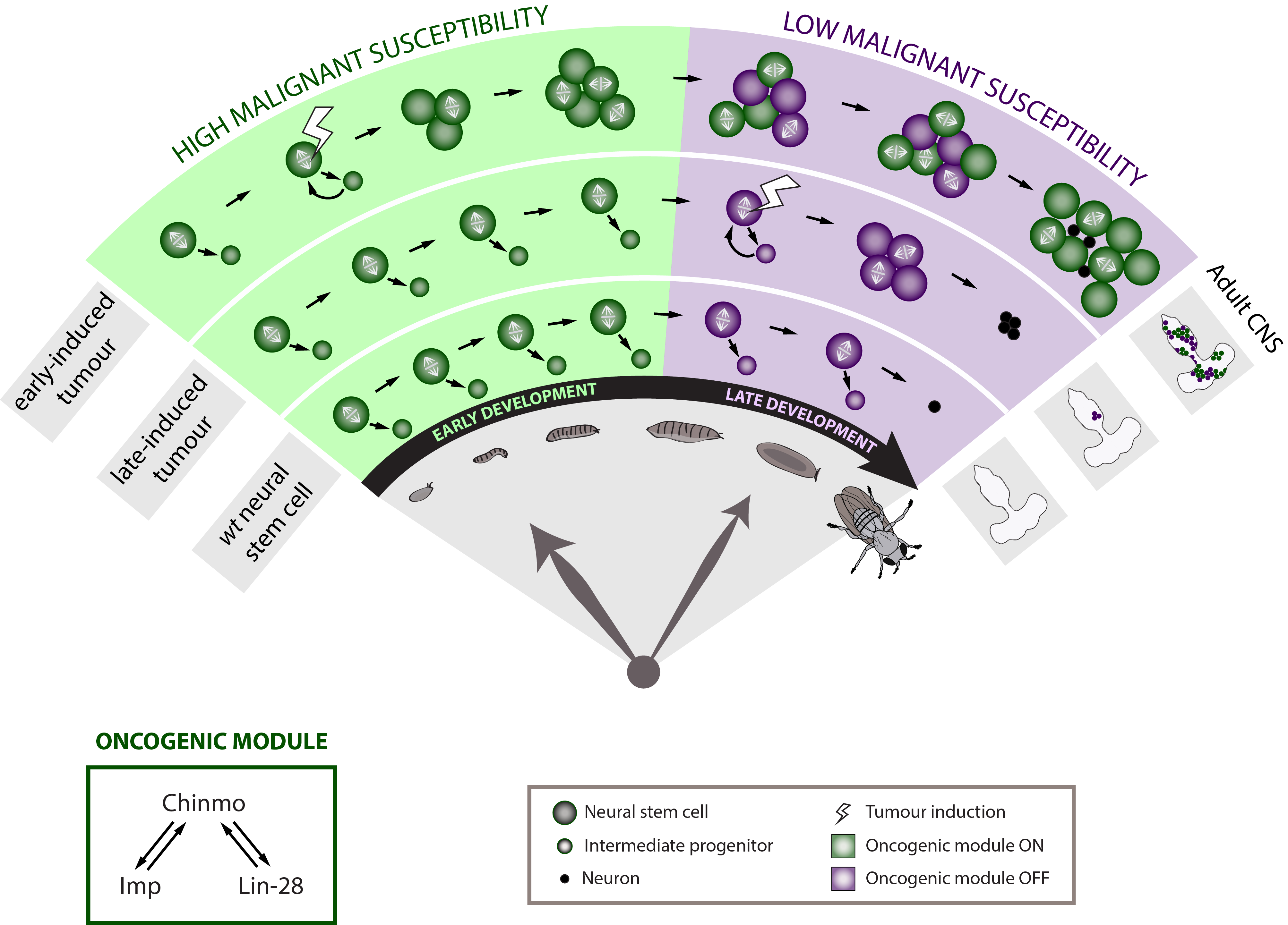
To further determine whether developmental time is a crucial component of the neural stem cell malignant transformation, we generated neural stem cell tumours at different moments along development. Tumours were initiated either during early or late larval development. Strikingly, when given the same time to grow, the late-induced tumours were largely unable to resist to terminal differentiation during metamorphosis so that few small benign tumours are found in the adult CNS whereas the early-induced tumours resisted strongly to differentiation and invaded the whole CNS. This observation demonstrates that during early development neural stem cells travel through a window of high malignant susceptibility.
Searching for the mechanisms regulating this early window of malignant susceptibility, we found that the same internal clock that limits the number of neural stem cell divisions also terminates the early window of susceptibility. Indeed, neural stem cells blocked in an early temporal identity remain prone to generate malignant tumours even when the latter are induced during late development. Thus, the temporal series must regulate genes that confer malignant susceptibility.
Malignant neural stem cells stays aberrantly young
Looking at the expression of Chinmo in the early VS late-induced tumours, we observed that Chinmo is largely expressed in the early-induced tumours but absent from the late-induced tumours. Thus, Chinmo expression correlates with malignant susceptibility in the tumorous neural stem cells. In order to establish a causality link between Chinmo’s expression in the neural stem cell and its malignant potential, we conducted gain and loss of function experiments within the early-induced tumours. We were able to show that Chinmo promotes tumour cell proliferation and resistance to differentiation. By means of a transcriptomic analysis, we identified numerous potential targets of Chinmo within the tumours. We selected the two best positive targets of Chinmo as candidates involved in the malignant process: the RNA-binding proteins Imp and Lin-28. Further genetic manipulations within the tumours allowed us to demonstrate that Imp largely contributes to the tumour growth-promoting effect of Chinmo, and Lin-28 at a lesser extent. Interestingly, Imp and Lin-28 exert also a post-transcriptional positive feedback control on Chinmo in the tumour. Thus, Chinmo, Imp and Lin-28 form an oncogenic module supporting the continuous growth of the tumour and its cancerous properties. Interestingly, we could observe that Chinmo, Imp and Lin-28 expression patterns perfectly overlap along normal development. The three genes are co-expressed in young neural stem cells but silenced in older ones. As such they define an early developmental window with specific growth and differentiation-resisting properties. However, in the developmental context, their interdependency is less clear. Together these results suggest that in our tumour model, the sub-population of cells that aberrantly maintains this oncogenic module typical from early development represents the cancer cells that propagate unlimited growth.
Relevance to paediatric cancers in humans
These results therefore demonstrate that Drosophila neural progenitors transit through a window of malignant susceptibility during early development. Moreover they reveal the timing mechanism and oncogenic module that regulate this early malignant susceptibility (Narbonne-Reveau et al., 2016). This is particularly exciting because it may explain why paediatric cancers, that are thought to originate during foetal life in humans, so rapidly become aggressive. Indeed, neural progenitors in mammals are thought to be also temporally patterned along the development although a temporal series has not been clearly identified yet (Naka et al., 2008) (Mattar et al., 2015). Moreover, Imp and Lin-28 are well conserved in mammals where they are expressed during the early development and often present in cancers (Bell et al., 2015; Carmel-Gross et al., 2016; Nielsen et al., 1999; Yang et al., 2015). Further studies would be needed to determine whether and how they promote malignant susceptibility in the early progenitors where they are expressed.
All together, this study helps identify the specific mechanisms that drive the rapid transformation of tumours with early developmental origins. In this context, work on Drosophila continues to illuminate us about the basic principles of life.
The early development defines a high malignant susceptibility window in the Drosophila neural stem cells through the activation of an oncogenic module composed of Chinmo, Imp and Lin-28.
Bell, J.L., Turlapati, R., Liu, T., Schulte, J.H., and Huttelmaier, S. (2015). IGF2BP1 Harbors Prognostic Significance by Gene Gain and Diverse Expression in Neuroblastoma. Journal of clinical oncology : official journal of the American Society of Clinical Oncology 33, 1285-1293.
Carmel-Gross, I., Bollag, N., Armon, L., and Urbach, A. (2016). LIN28: A Stem Cell Factor with a Key Role in Pediatric Tumor Formation. Stem Cells Dev 25, 367-377.
Caussinus, E., and Gonzalez, C. (2005). Induction of tumor growth by altered stem-cell asymmetric division in Drosophila melanogaster. Nat Genet 37, 1125-1129.
Isshiki, T., Pearson, B., Holbrook, S., and Doe, C.Q. (2001). Drosophila neuroblasts sequentially express transcription factors which specify the temporal identity of their neuronal progeny. Cell 106, 511-521.
Mattar, P., Ericson, J., Blackshaw, S., and Cayouette, M. (2015). A conserved regulatory logic controls temporal identity in mouse neural progenitors. Neuron 85, 497-504.
Maurange, C., Cheng, L., and Gould, A.P. (2008). Temporal transcription factors and their targets schedule the end of neural proliferation in Drosophila. Cell 133, 891-902.
Naka, H., Nakamura, S., Shimazaki, T., and Okano, H. (2008). Requirement for COUP-TFI and II in the temporal specification of neural stem cells in CNS development. Nature neuroscience 11, 1014-1023.
Narbonne-Reveau, K., Lanet, E., Dillard, C., Foppolo, S., Chen, C.H., Parrinello, H., Rialle, S., Sokol, N.S., and Maurange, C. (2016). Neural stem cell-encoded temporal patterning delineates an early window of malignant susceptibility in Drosophila. eLife 5.
Nielsen, J., Christiansen, J., Lykke-Andersen, J., Johnsen, A.H., Wewer, U.M., and Nielsen, F.C. (1999). A family of insulin-like growth factor II mRNA-binding proteins represses translation in late development. Molecular and cellular biology 19, 1262-1270.
Yang, M., Yang, S.L., Herrlinger, S., Liang, C., Dzieciatkowska, M., Hansen, K.C., Desai, R., Nagy, A., Niswander, L., Moss, E.G., et al. (2015). Lin28 promotes the proliferative capacity of neural progenitor cells in brain development. Development 142, 1616-1627.
A postdoctoral position is available in the Page laboratory in the Department of Neuroscience at The Scripps Research Institute in Jupiter, Florida to investigate mechanisms of neurogenesis & gliogenesis, axon/dendrite growth and synaptic connectivity in the developing mouse cerebral cortex. A major goal of this project is to understand the influence of genetic risk factors for autism and other neurodevelopmental disorders on these processes at the molecular/cellular, circuit and behavioral levels. For information about the laboratory, please visit: www.scripps.edu/page/.
Requirements: In addition to a PhD, highly motivated candidates with expertise in developmental biology, genetics, molecular/cell biology, imaging or behavioral neuroscience are encouraged to apply. Interested candidates should send a cover letter, CV and contact details for three references to Damon Page, Ph.D., email: paged@scripps.edu.
TSRI embraces diversity & recognizes it as being a key to our success. EOE/M/F/V/D
A fundamental property of all organisms is the remarkable consistency within the formation of the body plan during development. A handful of conserved molecular pathways are responsible for morphological patterning in embryos across different taxa, despite an enormous diversity of embryonic size in nature. Remarkably, morphological patterns are scaled to tissue size at the time of development and same structures are maintained in relative proportions to the overall size.
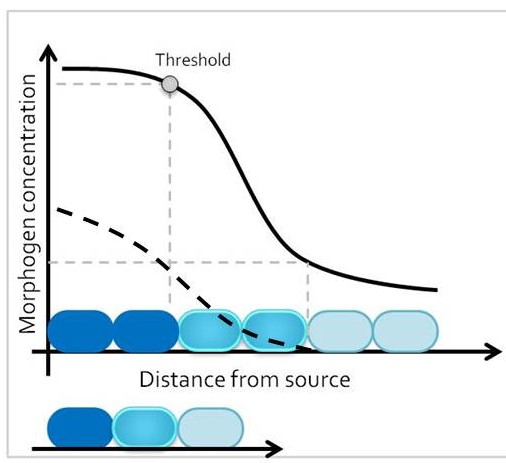
One classic mechanism of pattern formation is the morphogen model, where a molecule – the morphogen – is secreted from cells at one end of a developmental field, and diffuses into the target tissue (Lander 2007). At any given time point, morphogen concentration is different around cells that are closer versus further away from the morphogen source. The target tissue responds by activating distinct transcriptional programs in progenitor domains that are exposed to different morphogen concentrations, leading to establishment of different cell fates along the length of the tissue. Therefore, size of the target tissue is an integral component of the patterning process.
How is the morphogen mechanism adjusted, then, to trigger the same transcriptional responses in proportional domains across tissues of different sizes in closely related species? In many instances, morphological patterns are set when tissues size is similar: human embryos are roughly the same size during embryogenesis, and differential growth can account for why taller and smaller human individuals nevertheless share similar body proportions. However, in other cases members of related taxa have quite different sizes during embryonic development, when the morphological pattern is being established. How conserved pathways that regulate patterning are modified to generate a size-invariant output is a biological problem that still remains to be solved (Figure 1).
Figure 1. How domain identities are established in tissues of different sizes in proportion to the overall tissue size remains to be a solved
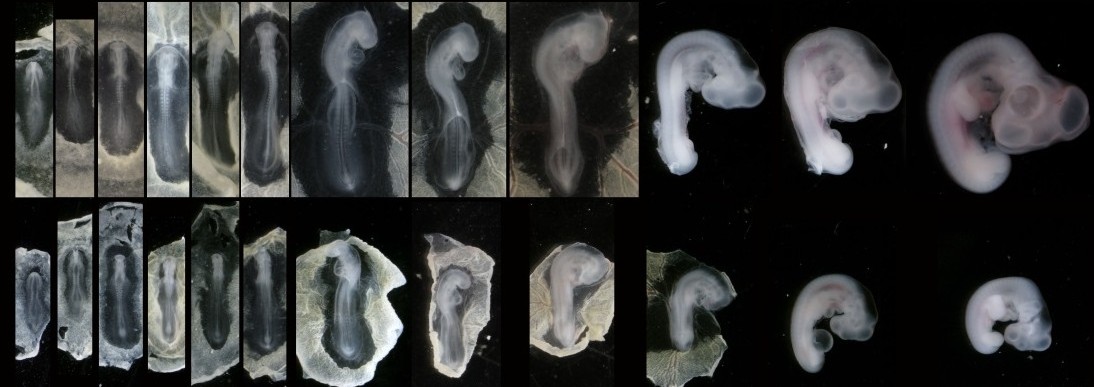
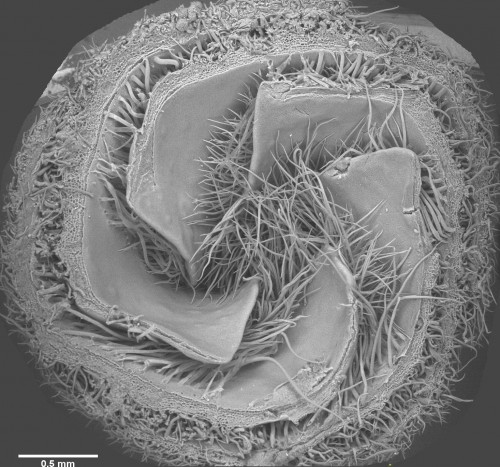
In our recent paper from the Tabin lab, we tackled this problem through the study of aves, a clad of organisms that show a great variation in size – think of the range of egg sizes from an ostrich egg to a hummingbird egg (Uygur et al. 2016). Yet, these animals are related close enough that we would expect patterning mechanisms to be highly conserved. This gave us the flexibility to study how scaling of the body plan is achieved across two organisms with a drastic difference in embryonic size, the chick (Gallus gallus) and the tiny zebra finch (Taeniopygia guttata) (Figure 2). The neural tube provided to be an excellent model system to explore this question in aves. Pioneering studies in the past two decades have shown that the sonic hedgehog (SHH) morphogen secreted from the notochord and floorplate induces distinct transcriptional programs and cell fates at different concentrations along the ventral half of the neural tube (Liem et al. 2000; Dessaud et al. 2007; Stamataki et al. 2005). As is the case with the morphogen model systems, tissue size and distance from source is critical to patterning onset in this context.
Figure 2. Chick and zebra finch embryos have a drastic difference in size across development
When we examined the establishment of pattern in the ventral neural tube of chick versus zebra finch embryos, we were surprised to find out that it is not only the tissue size that varies between the two species, but also the duration of the patterning process. Across highly similar developmental stages and timepoints, the zebra finch ventral neural tube is patterned much faster compared to the chick neural tube. However, when we compare the levels of morphogen secretion, we found out that the zebra finch notochord, which is drastically smaller than the chick notochord, is also generating a morphogen gradient of much lower amplitude throughout development. Even though the smaller species is producing less morphogen, it is patterning the tissue at a much faster rate. What is going on?
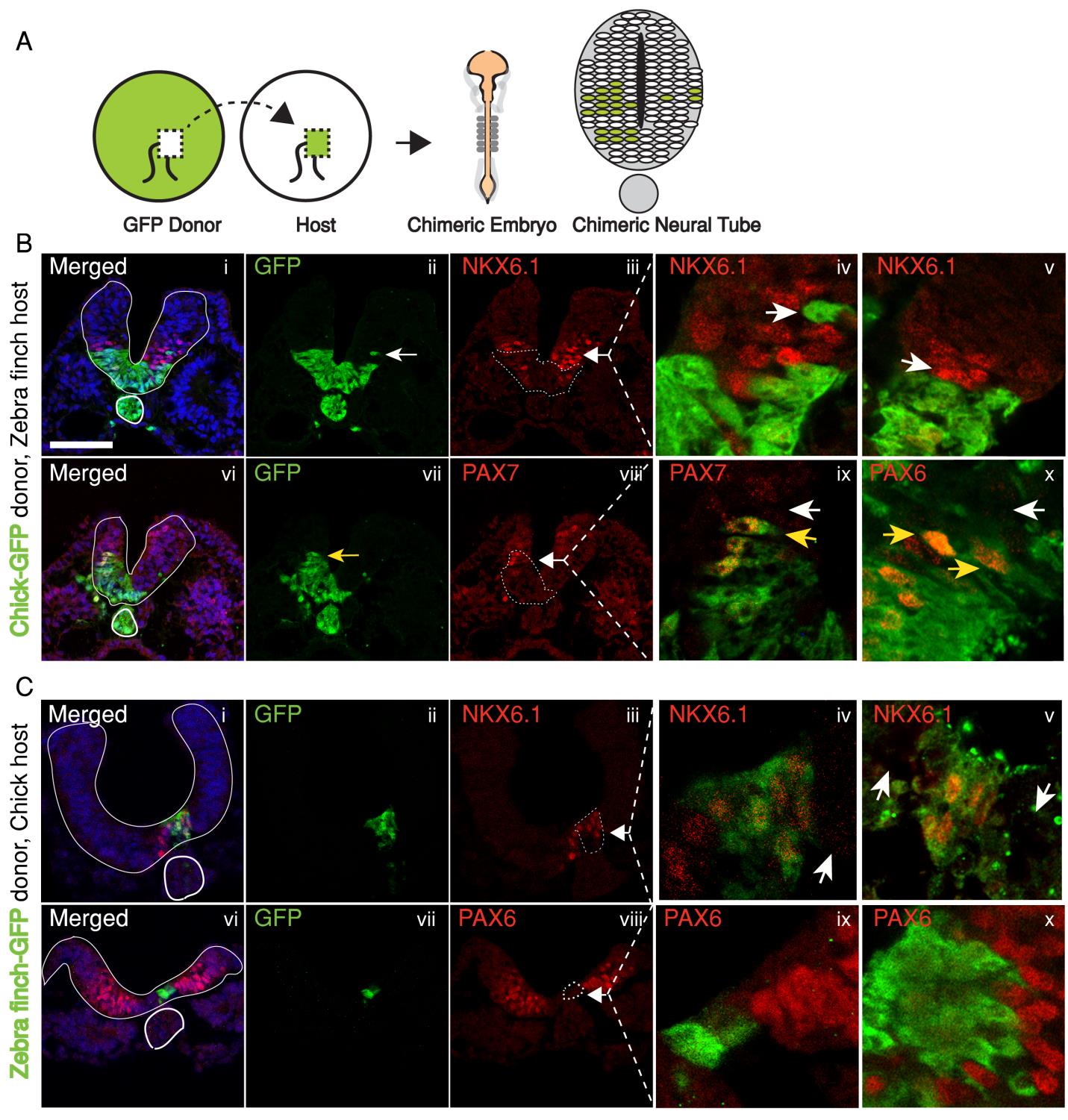
In the past, adjustments to the morphogen mediated patterning systems have been explored at the level of adjusting the morphogen gradient itself(Cheung et al. 2014; Gregor et al. 2005; Ben-Zvi & Barkai 2010). Even though this may still be an interesting aspect of the scaling of the neural tube patterning, our findings lead us to explore differences in morphogen responsiveness. According to our initial observations, patterning process in the smaller zebra finch tissue is completed much faster, with less morphogen in the tissue. When exposed to the same concentration of the sonic hedgehog morphogen, would cells from the zebra finch versus chick turn on the same transcriptional program and cell fates? We turned back to classical embryology to answer this question and generated chimeric embryos at a very early time point in development (at 12 hours, to be exact). By transplanting cells between transgenic lines from the two species, we generated embryos with chimeric neural tubes that were made up of both zebra finch and chick cells, where cells from the two species were exposed to the same developmental signals and received the same level of SHH morphogen from a single source. Remarkably, we observed zebra finch cells to be much more sensitive to the morphogen compared to adjacent chick cells that are same distance away from the source. An analysis on the expression of different transcription factors confirmed that the transcriptional profile of the zebra finch cells were of a higher level morphogen threshold, as if they had seen a higher concentration of SHH for longer durations. This shows cell autonomous differences in morphogen sensitivity account for differential response to the morphogen and altered rates of patterning (Figure 3).
Figure 3. Chick and zebra finch have cell autonomous differential sensitivity to the morphogen SHH
The strength of using the neural tube as a model is that the readout for morphogen activity is very well characterized in this system. We took advantage of this to quantify the differential response of neural progenitors to Sonic hedgehog in the two species. In an explant in-vitro culture system, we analyzed response to different concentrations and durations of SHH in neural tube naive explants isolated from either zebra finch or chick embryos. Indeed, all morphogen thresholds that have previously been identified for chick cells were lower for zebra finch cells, meaning that the zebra finch tissue can induce same cell types at lower concentrations and lower exposure durations.
Our studies on the mechanism of this difference showed that the differential response persists even when the SHH pathway is activated directly through its Smoothened receptor via an agonist. This finding revealed that the cell autonomous differential response is an intra-cellular difference downstream of the patched and smoothend receptors. We further showed that differential levels of the sonic hedgehog transcriptional effortors may be responsible for the differential response, as the two species display different levels of the GLI3 transcript. When we manipulated the zebra finch embryos to express higher levels of GLI3, we observed the speed of patterning is altered and the domain ratios are not scaled to the smaller tissue size.
Our study offers a potential explanation to how tissues of different scales achieve proportional domain sizes when using the same signaling pathways for patterning. It appears that in the case of chick and zebra finch, two avian embryos of different sizes, cell intrinsic differences in morphogen sensitivity control scaling of cell fate patterns in the ventral neural tube. We hope this work will provide new venues to study pattern scaling in order to enhance our understanding of how evolutionary mechanisms accommodate adjustments to body size.
Ben-Zvi, D. & Barkai, N., 2010. Scaling of morphogen gradients by an expansion-repression integral feedback control. Proceedings of the National Academy of Sciences of the United States of America, 107(15), pp.6924–9. Available at: http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=2872437&tool=pmcentrez&rendertype=abstract [Accessed July 11, 2014].
Cheung, D. et al., 2014. Adaptation of the length scale and amplitude of the Bicoid gradient profile to achieve robust patterning in abnormally large Drosophila melanogaster embryos. Development (Cambridge, England), 141(1), pp.124–35. Available at: http://www.ncbi.nlm.nih.gov/pubmed/24284208 [Accessed April 12, 2014].
Dessaud, E. et al., 2007. Interpretation of the sonic hedgehog morphogen gradient by a temporal adaptation mechanism. Nature, 450(7170), pp.717–20. Available at: http://www.ncbi.nlm.nih.gov/pubmed/18046410 [Accessed March 21, 2014].
Gregor, T. et al., 2005. Diffusion and scaling during early embryonic pattern formation. Proceedings of the National Academy of Sciences of the United States of America, 102(51), pp.18403–7. Available at: http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=1311912&tool=pmcentrez&rendertype=abstract [Accessed March 26, 2014].
Lander, A.D., 2007. Morpheus unbound: reimagining the morphogen gradient. Cell, 128(2), pp.245–56. Available at: http://www.sciencedirect.com/science/article/pii/S0092867407000517 [Accessed December 10, 2014].
Liem, K.F., Jessell, T.M. & Briscoe, J., 2000. Regulation of the neural patterning activity of sonic hedgehog by secreted BMP inhibitors expressed by notochord and somites. Development (Cambridge, England), 127(22), pp.4855–66. Available at: http://www.ncbi.nlm.nih.gov/pubmed/11044400 [Accessed August 12, 2014].
Stamataki, D. et al., 2005. A gradient of Gli activity mediates graded Sonic Hedgehog signaling in the neural tube. , pp.626–641.
Uygur, A. et al., 2016. Scaling Pattern to Variations in Size during Development of the Vertebrate Neural Tube. Developmental Cell, 37(2), pp.127–135.
In this new feature, we conduct a trawl for preprints to find the latest developmental biology and related research from the previous month.
Scientists are increasingly using preprints as a way of releasing research to the community without having to wait for the peer review process, and we want to promote this content here. Several sites host biology preprints, most notably bioRxiv, but also arXiv, f1000 Research, and PeerJ Preprints (for more info on preprints, ASAPbio has a useful FAQ page).
This month we found content from bioRxiv, arXiv, F1000 Research and PeerJ spanning developmental biology research, including investigations into cell fate in preimplantation mouse embryos and worm larvae, a new technique for bi-allelic gene knockouts, and a method for making midbrain-specific organoids.
Zika Virus in the Human Placenta and Developing Brain: Cell Tropism and Drug Inhibition. HannaRetallack, ElizabethDi Lullo, CarolinaArias, Kristeene A.Knopp, CarmenSandoval-Espinosa, Matthew T.Laurie, YanZhou,MatthewGormley, Walter R.Mancia Leon, RobertKrencik, Erik M.Ullian, JulienSpatazza, Alex A.Pollen, KatherineOna, Tomasz J.Nowakowski, Joseph L.DeRisi, Susan J.Fisher, Arnold R. Kriegstein
Here are the highlights from the current issue of Development:
A pioneer role for PBX1 in neurogenesis
The adult rodent subventricular zone (SVZ) is a neurogenic niche that provides new neurons and glia to the brain. A number of transcription factors, including MEIS2 and PAX6, are known to be required to promote the neuronal cell fate. PBX family proteins can interact with both MEIS and PAX family factors, and PBX1 is known to be expressed in the adult mouse forebrain. Dorothea Schulte and colleagues therefore set out to test the role of PBX1 in SVZ neurogenesis (p. 2281). They show that Pbx1expression is found in rapidly proliferating SVZ progenitors, as well as in subsets of their progeny in the olfactory bulb. Its ortholog, PBX2, is more widely expressed in the forebrain. Deletion of both genes in the adult SVZ leads to a reduction in neurogenesis and a concomitant increase in oligodendrogliogenesis – an alternative fate for SVZ progenitors. Although the authors can detect PBX1 binding to MEIS2 and PAX6, the phenotype is distinct from functional blockade of these factors, suggesting an at least partially independent function. Intriguingly, PBX1 appears to bind its targets before they are transcriptionally activated, indicating a putative pioneer factor function for PBX1. These data identify PBX1 as an important new regulator of SVZ neurogenesis.
A tyrosine kinase for FERtilisation
Mammalian sperm need to undergo capacitation to become competent to fertilise the oocyte. In vivo, this occurs in the female reproductive tract, whereas in vitro it can be triggered by culture in defined media. One of the key events during capacitation is cAMP-dependent tyrosine phosphorylation of various proteins. Although this phenomenon has been well described in many species, the kinase responsible has been elusive. Various candidates have been proposed, including SRC, FAK and PYK2; notably, inhibitors for the latter two enzymes block tyrosine phosphorylation in human and horse sperm. However, definitive evidence for the identity of the kinase has been lacking. Now (p. 2325), Pablo Visconti and co-workers show that, while FAK and PYK2 inhibitors also block tyrosine phosphorylation in mouse sperm (without blocking PKA activation), neither is the responsible factor. Instead, it is the FER kinase – also targeted by the same inhibitors – that carries out capacitation-associated tyrosine phosphorylation. However, and surprisingly, FER is not required for fertilisation in vivo, suggesting that the presumed involvement of tyrosine phosphorylation in acquisition of sperm competence may need to be revisited.
Coordinating morphogenesis and differentiation with WNT
The mammalian salivary gland is a valuable model for analysis of the morphogenetic and differentiation events that occur during branching morphogenesis of organs. Following a period of growth and branching, epithelial cells of the salivary gland differentiate down ductal or acinar routes. What are the signalling mechanisms that control morphogenesis, differentiation and lineage choice, and how are these processes coordinated to ensure appropriate size and composition of the final organ? The FGF pathway is known to play key roles in salivary gland development, including via the activation of KIT signalling – which promotes the expansion of distal (future acinar) progenitors. Moreover, WNT signalling has been implicated in salivary gland morphogenesis, although its function is unclear. On p.2311, Akira Kikuchi and colleagues use genetic and pharmacological approaches, both in vivo and in vitro, to demonstrate that mesenchymally derived WNT signals inhibit acinar differentiation and maintain end bud cells in an undifferentiated state, to promote secondary and tertiary duct formation. Mechanistically, WNT activity promotes MYB-dependent inhibition of Kit expression; KIT directs acinar differentiation through AKT. As WNT pathway activity is dynamically regulated, the authors propose this as a means by which salivary gland morphogenesis and differentiation can be spatiotemporally coordinated.
PLUS!
Cell behaviors and dynamics during angiogenesis
Vascular networks are formed and maintained through a multitude of angiogenic processes, such as sprouting, anastomosis and pruning. Only recently has it become possible to study the behavior of the endothelial cells that contribute to these networks at a single-cell level in vivo. Here, Markus Affolter and colleagues summarizes what is known about endothelial cell behavior during developmental angiogenesis, focusing on the morphogenetic changes that these cells undergo. See the Review on. p. 2249.
Generation of intestinal surface: an absorbing tale
The vertebrate small intestine requires an enormous surface area to effectively absorb nutrients from food. Morphological adaptations required to establish this extensive surface include generation of an extremely long tube and convolution of the absorptive surface of the tube into villi and microvilli. In their Review, Deborah Gumucio and colleagues discuss recent findings regarding the morphogenetic and molecular processes required for intestinal tube elongation and surface convolution in different species. See the Review on p. 2261.
Dental mesenchymal stem cells
Mammalian teeth harbour mesenchymal stem cells (MSCs), which contribute to tooth growth and repair. These dental MSCs possess many in vitro features of bone marrow-derived MSCs, including clonogenicity, expression of certain markers, and following stimulation, differentiation into cells that have the characteristics of osteoblasts, chondrocytes and adipocytes. Here, Paul Sharpe outlines some recent discoveries in dental MSC function and behaviour and discusses how these and other advances are paving the way for the development of new biologically based dental therapies. See the Review on p. 2273.
Unraveling Development
Embryonic development is a complex and regulated spatiotemporal ensemble of signaling cues that control cell differentiation. Most of what we now know comes from experimenting directly on embryos. This provides biological realism, but involves a sea of uncontrolled/unobserved variables that sometimes obscures the basic underlying mechanisms. More recently, in vitro models, based on the differentiation of stem cells, have come into favor. These sacrifice some biological authenticity but have the distinct advantage of greater experimental ease and control over variables. Nevertheless, the culture dish or titer plate falls short of what could reasonably be considered an ideal imitation of an embryonic environment. As a result, in vitro models often give inconsistent or variable results.
Our goal has been to develop an experimental platform that captures some of the realism of in vivo models but with the ease and experimental control of in vitro methods. Our mantra is, “If you create the same physiochemical environment in vitro that occurs in vivo, cells will respond in the same way.” This not only includes the creation of molecular gradients and substrates, but the ability to alter these over time. To create this “ideal” in vitro environment, we’ve been using microfluidics.
A (very) brief history of microfluidics
Microfluidics is a growing field that deals with the fabrication of devices to control the flow and handling of small liquid volumes ranging from several hundred microliters to several picoliters. The actual physical size of any single microfluidic component can vary between several millimeters to several micrometers (about the size of a single cell) depending on the intended use, i.e. channels, valves, pumps, etc. .Examples of common microfluidic devices include: microbioreactors1, microsensors2, microanalysis3, lab-on-chip4, etc.
The field traces its origins to 1959 and the first demonstration of an integrated electronic circuit by Noyce and Kilby5. This discovery ushered in the microelectronics era and fueled the development of a plethora of high precision microfabrication technologies. Microfluidics “borrowed” these technologies to micromachine microstructures that comprise channels, valves and pumps. One of the first commercial applications of a microfluidic device was in 1979 when Hewlett-Packard invented the inkjet printer.
Because of its ancestral ties to microelectronics, most microfluidic systems were initially fabricated in silicon, but both technologies and materials have since expanded past their initial origins. More details of the history of silicon micromachining and microfabrication can be found here.
The role of microfluidics in developmental biology
Despite some setbacks, microfluidics are finally being applied to biology. Applications involving drug research and so-called “Organs-on-Chip” have enjoyed considerable attention. However, using devices to address fundamental biological questions has been slower to evolve. We find this rather surprising because developmental studies are particularly well suited to the strengths of microfluidics. Recent reports have revealed the incredible ability of embryonic stem cells to self-organize given appropriate instructive environment6-9. These are at present limited to bath applications of soluble factors. Microfluidic technologies can provide a much needed additional layer of control for more biologically relevant experiments.
This background was what prompted us to develop a microfluidic device that could mimic morphogen gradients. The device is comprised of a cell culture chamber flanked by supply channels reminiscent of how tissues are perfused by capillary beds. Soluble chemicals can freely diffuse into the cell chamber establishing long-term, stable gradients. Using this simple microfluidic device and only three morphogen gradients, Shh, BMP and RA, we were able to recapitulate much of the neural tube development including motor neuron spatial organization10. However, it is clear that not all neural types are represented in our little artificial spinal cord. Something is missing. Whether this is another morphogen or signal, electrical gradients or just wrong timings for the signals we’re using, the search should be extremely interesting. For example, the anteroposterior axis develops with its own morphogen gradients concomitantly with the dorsoventral axis. It’s likely that cells in the early spinal cord simultaneously integrate signals from both anteroposterior and dorsoventral axes.
We should be able to test this. The microfluidic device we have engineered is capable of creating almost any “designer” chemical landscape11 with any number of arbitrary morphogens. So there’s a whole set of experiments we are currently doing to explore some of these ideas. It’s also an example of how useful the technique could be to to developmental biology. All that is needed is an interesting question and a microfluidic design to test it. Although we haven’t yet faithfully replicated the physiochemical landscape in vitro, I think we’ve taken an important first step. I’m not sure where the journey will end but I’m hoping that we learn some interesting biology on the way. Fortunately, there is still plenty of research to be done.
Figure 1: Mouse embryonic stem cells cultured in a four-port microfluidic device for 9 days. GFP expression indicates the presence of post-mitotic motor neurons, which clearly favor a defined spatial region as dictated by instructive cues supplied to the cells. Cells are superimposed with a red soluble fluorescent dye to illustrate the shape of the gradient formed in the microdevice. Overall chamber dimensions approximately 1.5mm at the widest point.
1. Grünberger, A., Wiechert, W. & Kohlheyer, D. Single-cell microfluidics: opportunity for bioprocess development. Curr Opin Biotechnol 29, 15–23 (2014).
2. Erickson, D. & Li, D. Integrated microfluidic devices. Anal Chim Acta 507, 11–26 (2004).
3. Reyes, D. R., Iossifidis, D., Auroux, P.-A. & Manz, A. Micro Total Analysis Systems. 1. Introduction, Theory, and Technology. Anal Chem 74, 2623–2636 (2002).
4. Haeberle, S. & Zengerle, R. Microfluidic platforms for lab-on-a-chip applications. Lab Chip 7, 1094–1110 (2007).
5. Kilby, J. S. Miniaturized electronic circuits. (1964).
6. Warmflash, A., Sorre, B., Etoc, F., Siggia, E. D. & Brivanlou, A. H. A method to recapitulate early embryonic spatial patterning in human embryonic stem cells. Nat Meth 11, 847–854 (2014).
7. van den Brink, S. C. et al. Symmetry breaking, germ layer specification and axial organisation in aggregates of mouse embryonic stem cells. Development 141, 4231–4242 (2014).
8. 3D Reconstitution of the Patterned Neural Tube from Embryonic Stem Cells. 1–34 (2014). doi:10.1016/j.stemcr.2014.09.020
9. Poh, Y.-C. et al. Generation of organized germ layers from a single mouse embryonic stem cell. Nat Commun 5, 4000 (2014).
10. Demers, C. J. et al. Development-on-chip: in vitro neural tube patterning with a microfluidic device. Development 143, 1884–1892 (2016).
11. Smith, R. L., Demers, C. J. & Collins, S. D. Microfluidic device for the combinatorial application and maintenance of dynamically imposed diffusional gradients. Microfluidics and … 9, 613–622 (2010).
I woke up this morning to a Facebook reminder of where I was 5 years ago. I was in Lille, France, on a 2 month sabbatical at Université Lille 1 from my PhD at the University of Cambridge, UK. It was supported by an EU collaborative grant to promote scientific interaction between member states.
By all measures, it was a roaring success – far better than we had dared to hope. I was working with one of the world’s least purifiable proteins, a transcription factor called Neurogenin2, and trying to study its phosphorylation status by NMR (nuclear magnetic resonance) imaging. We had gone in with the expectation that nothing would work but that it was worth a go, and I came out with an entire thesis chapter based on 2 months work, and what we would later publish as, “Phosphorylation in intrinsically disordered regions regulates the activity of Neurogenin2.”
Not only was it directly useful as a scientific endeavor, it was also a great experience. I learned about some of the intricacies of the French system, working with an independent researcher at CNRS who did not run the lab, but was nonetheless a researcher in her own right. My Parisian-French-with-a-Northern-Irish-accent proved useless in the face of a local dialect, Ch’ti, on the French-Belgian border. I went to Waterloo near its anniversary; I ate vast amounts of cheese; I marvelled at the holidays the French seemed to have every other week. I lived in dank student accommodation (about which my hosts were very apologetic) but I have nothing but fond memories for the 2 months I spent there. In the space of the last week, I have realised how much I took for granted that this was an opportunity available to me.
On June 23rd, the UK voted in a referendum to leave the EU.
I’m not going to comment on whether that is or isn’t going to happen, what it does or doesn’t mean for democracy in the UK – there’s enough on that already. What I’d like to do here is briefly address some areas crucial to science: funding, international collaboration and the mobility of scientific researchers. The implications for science from Brexit are both uncertain and unlikely to be clarified soon. And scientific research, as an enterprise, does not deal well with uncertainty.
What did I mean by “a science funding system already strained”? Last August, I made a brief visit to the UK, gave some talks on my research and scoped out the situation for academic jobs, as this was still the career track I was primarily pursuing. It became clear pretty quickly that funding was a key concern – and this was before Brexit was even really in anyone’s mind as an issue. Science funding by the UK has stagnated for years, and it was only through the major “Science is Vital” campaign that funding for science was not cut by the coalition government of 2010 – but neither was it increased. EU funding currently makes up around 15% of the UK’s research and development funding. The UK is only just behind Germany in the amount of money received from the EU for science funding and despite being a net contributor overall to the EU, has benefitted financially in terms of research funding (see Table 1 and surrounding discussion, “Examining Implications of Brexit for the UK Research Base”).
The official line at the moment is that nothing has changed; the UK is still an EU member for at least two years, and there seems to be a lot of certainty that the UK will continue to pay into the Horizon 2020 and EU research funding based on its GDP, much like Switzerland. However, I have not yet seen how this is easily reconciled with a major feature of the Leave campaign – immigration – and the Swiss have restricted their access to EU funds by restricting freedom of movement, particularly funds allowing young researchers to move around, such as Marie Skłodowska-Curie Research Fellowships and the Erasmus programme (if you click this link, note the first lines on the web page).
And this is crucial because the scientific enterprise is about so much more than just money; it is about people. More concerning, in my mind, than the issue of what will happen to funding, is what the introduction of added bureaucracy and complications will do to international collaborations, and what the question over mobility of researchers – and the recent spike in xenophobia and racism – will change in the minds of junior scientists looking to come to, return to, or stay in the UK.
Collaborations
Jo Johnson, fresh from the assurances given above about UK science, soon had to address the issue that UK researchers risk exclusion from collaborative projects in the EU. This is what I cannot reconcile in my mind about the assurances that legally, nothing has changed, and everything will carry on as before. Science funding is highly competitive. To obtain that funding, researchers will seek to minimize any risk of rejection of grant applications. The UK has stated that it is not going to trigger Article 50, the mechanism to leave the EU, until the new Prime Minister is decided, at the earliest in September. EU officials have in turn retorted that no assurances or negotiations will be carried out in that time; and negotiations will then take up to 2 years.
In this vacuum devoid of certainty, with no realistic assurances, I would be very surprised if researchers in the EU, albeit with a heavy heart, do not begin to rule out collaborations with UK researchers. It is even the case that although the official line has not changed, misinformation and requests to withdraw UK researchers from or to not involve UK researchers in projects may be filtering into the system. This is already happening anecdotally:
there is no consensus. People just don't know about future bids. I, too have heard of people being asked to leave H2020 etc
I titled this blog post using a direct quote from UK Justice Secretary Michael Gove, “people in this country have had enough of experts” (Michael Gove is one of the candidates for the leadership of the Conservative party, and therefore the position of Prime Minister). Xenophobia and racism have spiked recently in the UK. Reporting of hate crimes has increased. Issues of funding and academic job security aside, the UK is not presenting an attractive image to those of us outside it. There does not appear to be a desire to welcome those who are foreign or those with expertise, even in the very highest echelons of government. It is also not encouraging expats I have spoken with to return.
Anecdotal evidence exists that fellowship applications from the UK are being hit:
This also brings in the question of the effect on students and higher education. Current EU students and those entering in 2016 will have loan arrangements honoured, but there is no guarantee as yet with respect to entry from autumn 2017. Institutions are issuing statements along the lines of this from Vice-Chancellor Sir Leszek Borysiewicz at the University of Cambridge, and the President at University College London reacted to claims that UK universities will not suffer with fears that EU students may be lost from the UK, put off by tuition fees (students from the EU pay the same fees as UK students, and not international fees). UCL and other institutions in the UK have expressed dismay at the effects leaving the EU may have on their operation.
Concluding thoughts
In the process of negotiation, UK science and universities may end up with the relatively unchanged situation that many hope for. But that situation is a long way away, and fraught with uncertainty. Those who argue that all is fine and will continue to remain so, and that there is therefore not a problem, are not appreciating the incentives and risks that people consider as they move around to do science. Science is a highly mobile affair – look at Emmanuelle Charpentier’s movements around the world as a case in point – and the entire incentive structure for moving to the UK, and the risks involved for young researchers, have completely changed, if only temporarily, in the absence of any certainty of what is to come. Science is so competitive right now that many researchers just can’t afford to enter into uncertain positions. The longer the uncertainty continues, the more likely (and my own anecdotal evidence, admittedly small, overwhelming shows this) young scientists in the UK are to leave; and expats and foreign researchers are to stay away.
But perhaps these may not be factors that concern the UK in the end. After all, as we were told in no uncertain terms, “people in this country have had enough of experts.”
I am working on a follow-up post to this for the American Society of Cell Biology’s COMPASS blog in which I would like to include some more data as it appears, so please do help with data collection efforts.
Do you have thoughts or comments? Please feel free to get in touch below. Fact-checking and requests for clarification are particularly encouraged.
——————————————————————————————— Gary McDowell is the Executive of Director of Future of Research (FoR), a U.S. nonprofit that assists junior scientists in grassroots advocacy to promote solutions to problems they perceive with science, and the scientific enterprise. He spent 9 years discovering the joy of developmental biology working with the frog Xenopus laevis. He is currently a resident at the Moore Foundation-funded Manylabs open science skunkworks in San Francisco, CA.
This month we welcomed Aidan to the Node, who took over from Cat as Community Manager, and introduced himself here. He’s been enjoying settling in to the Company of Biologists office, and getting to know the site and its users.
We had 17 jobs and studentships posted on our jobs page, on everything from chromatin to membrane dynamics. We also had a host of other posts covering a typically wide range of content:
Our ‘A day in the life…’ series continued, and we first learned about what life is like in a gar lab. Martin Minarik journeys from Prague to Mexico to harvest eggs from these non-teleost fishes. We then shifted from the humid tropics to the clean room, where one PhD student fabricates microelectrodes for neuroscience applications.
Campaigns/Questions
We reposted a piece from the Genetics Society of America campaigning for the maintenance of funding for model organism databases (such as Flybase and ZFIN); you can sign the Letter of Support by following the link in the piece.
Finally, we asked our Questions of the Month, which really couldn’t have been about anything else: what does the UK referendum result mean to you as a scientist, and what can we as a community do about it? Get involved here. We also created a page with a bunch of post-referendum links.


 (No Ratings Yet)
(No Ratings Yet)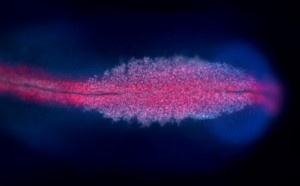
 (44 votes)
(44 votes)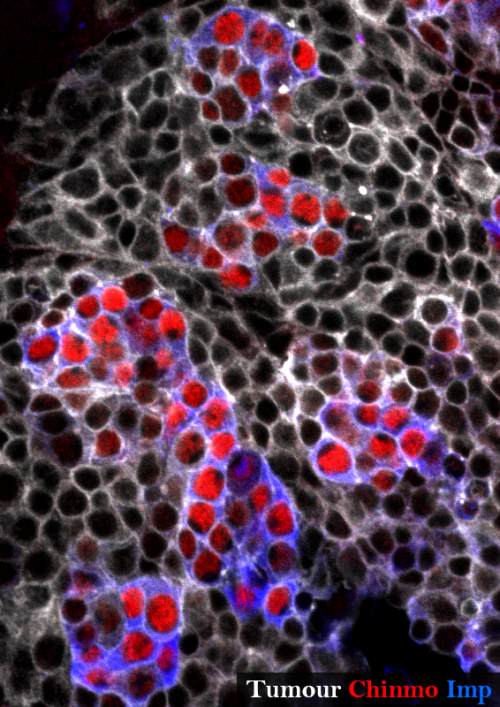
 (5 votes)
(5 votes)


 Vascular networks are formed and maintained through a multitude of angiogenic processes, such as sprouting, anastomosis and pruning. Only recently has it become possible to study the behavior of the endothelial cells that contribute to these networks at a single-cell level in vivo. Here, Markus Affolter and colleagues summarizes what is known about endothelial cell behavior during developmental angiogenesis, focusing on the morphogenetic changes that these cells undergo. See the Review on. p.
Vascular networks are formed and maintained through a multitude of angiogenic processes, such as sprouting, anastomosis and pruning. Only recently has it become possible to study the behavior of the endothelial cells that contribute to these networks at a single-cell level in vivo. Here, Markus Affolter and colleagues summarizes what is known about endothelial cell behavior during developmental angiogenesis, focusing on the morphogenetic changes that these cells undergo. See the Review on. p.  The vertebrate small intestine requires an enormous surface area to effectively absorb nutrients from food. Morphological adaptations required to establish this extensive surface include generation of an extremely long tube and convolution of the absorptive surface of the tube into villi and microvilli. In their Review,
The vertebrate small intestine requires an enormous surface area to effectively absorb nutrients from food. Morphological adaptations required to establish this extensive surface include generation of an extremely long tube and convolution of the absorptive surface of the tube into villi and microvilli. In their Review,  Mammalian teeth harbour mesenchymal stem cells (MSCs), which contribute to tooth growth and repair. These dental MSCs possess many in vitro features of bone marrow-derived MSCs, including clonogenicity, expression of certain markers, and following stimulation, differentiation into cells that have the characteristics of osteoblasts, chondrocytes and adipocytes. Here, Paul Sharpe outlines some recent discoveries in dental MSC function and behaviour and discusses how these and other advances are paving the way for the development of new biologically based dental therapies. See the Review on p.
Mammalian teeth harbour mesenchymal stem cells (MSCs), which contribute to tooth growth and repair. These dental MSCs possess many in vitro features of bone marrow-derived MSCs, including clonogenicity, expression of certain markers, and following stimulation, differentiation into cells that have the characteristics of osteoblasts, chondrocytes and adipocytes. Here, Paul Sharpe outlines some recent discoveries in dental MSC function and behaviour and discusses how these and other advances are paving the way for the development of new biologically based dental therapies. See the Review on p. 

 Julia Turan wrote about the
Julia Turan wrote about the  We heard about the Young Embryologist Network meeting in London from two perspectives:
We heard about the Young Embryologist Network meeting in London from two perspectives:  Our ‘A day in the life…’ series continued, and
Our ‘A day in the life…’ series continued, and  e asked our Questions of the Month, which really couldn’t have been about anything else: what does the UK referendum result mean to you as a scientist, and what can we as a community do about it?
e asked our Questions of the Month, which really couldn’t have been about anything else: what does the UK referendum result mean to you as a scientist, and what can we as a community do about it?