From autumn 2016, a postdoctoral research position is available in the Panfilio lab to investigate morphogenesis of the insect extraembryonic (EE) membranes. These simple epithelia are highly dynamic in their role as transient, protective covers for the embryo. The aim is to understand how EE morphogenesis works at multiple levels of biological organization, from cellular ultrastructure to coordination between tissue sheets. The primary research species for the project is the red flour beetle, Tribolium castaneum, an excellent model for studying interactions between developing epithelia via fluorescent live imaging and sophisticated genetic approaches (see the lab’s recent publications in eLife5:e13834, and Developmentdoi:10.1242/dev.133280).
extraembryonic cell shape changes during tissue remodeling
The lab is in the Institute for Developmental Biology, University of Cologne, Germany (lab web page: http://tinyurl.com/c24gpte), and has research and collaboration links with other evolutionary, developmental, and insect labs across the Biology Department of the university. With one million inhabitants, Cologne is an international, vibrant city that is well connected within western Europe.
Successful applicants will have a strong interest in developmental genetics and imaging, demonstrated by holding a Ph.D. degree and at least one first author publication in a relevant field. Required skills include standard developmental genetics techniques associated with RNAi, tissue staining, and conventional light microscopy. Experience with electron microscopy, quantitative analysis of multidimensional imaging data, transgenesis, and/or transcriptomics is desired. Depending on expertise and interest, there are possibilities for the applicant to develop his/her own research program among the topics of dynamic tissue adhesion, remodeling of the extracellular environment, and comparative transcriptional profiling. The working language of the lab is English, and strong oral and written communication skills are expected.
The position is for one year in the first instance with the possibility for annual extensions. The University of Cologne is an equal opportunity employer in compliance with German disability laws. Women and persons with disabilities are strongly encouraged to apply.
To apply send a research statement, CV, starting date availability, and contact details (including e-mail address and phone number) for two references as a single PDF file to Kristen.Panfilio@alum.swarthmore.edu. Informal enquiries to further discuss the position are welcome. All applications will be considered until 20 August 2016.
It seemed like an ordinary morning, with the lecture on Drosophila genetics due to start at 8 o’clock. But when the professor walked in something remarkable happened: instead of starting the lecture right away, he posed an unusual question: “One of the students of this class has a publication on this topic. George Streisinger, where are you?” But the student he was looking for was not there – he slept in. Lotte Sielman, sitting in the audience, was just as puzzled as her classmates about the curious incident. She could not have guessed that George Streisinger would soon become her husband and would contribute a number of exciting and important chapters to the history of modern molecular genetics.
Not to forget, the other author of the above mentioned Drosophila paper was one of the giants of the 20th century biology, Theodosius Dobzhansky. And if we look at the later works of Streisinger, this is not even unusual: being one of the most talented geneticists of his era, he built strong friendships and professional relations with almost all of the celebrated biologists of the 1950s.
From Budapest to Manhattan
Streisinger was born in Budapest on 27th December 1927. His parents were merchants: his father, Andor Streisinger dealt with leather goods and his mother, Margit Freund was selling ties. Little Gyuri (as he was called at that time) started his life in the city: his family first lived in the centre in one of the later yellow-star houses, then in the Castle district, followed by a few years in the center city again, where his father kept pigeons on the roof. (Andor Streisinger probably had a serious injury during WWI, as he was entitled to an injury allowance.) Gyuri started school at the Lutheran Primary School next door, and later followed brother Ervin (7 years his senior) to the legendary Fasori Lutheran Secondary School. (The fame of the Fasori School is due to the fact that it was also the Alma Mater for a number of other Hungarian-born luminaries of the era: Nobel-prize winners Eugene Wiegner and John Harsanyi, the mathematician John von Neumann, and the physicist Edward Teller.)
The Streisingers lived a typical upper-middle-class life. A family cook and a maid helped with the housework, but also took care of Gyuri – perhaps this was the origin of Streisinger’s later fondness for culinary delights. He was an omnivorous gourmand, who was ready to prepare or taste any kind of food. Well, almost any: kohlrabi was the exception. He hated it as a child, but if he did not eat it for lunch or dinner, it was back on his plate next morning.
The little boy was diagnosed with a heart murmur at an early age, which prohibited him from taking part in the PE classes or playing soccer with his friends. Instead, together with Ervin, they went out to catch butterflies in the mountains of Buda. These early voyages as a budding amateur naturalist became the inspiration for his later career.
His attraction to natural sciences would have been supported by the teachers in the Fasori School, however the turbulent history of the 20th century did not let this happen. After a few months in the famed institution, the increasingly severe discrimination against Jews forced the family to pack up and leave everything behind for a better life on the other side of the Atlantic.
Margit Freund and her sons George (on the left) and Ervin (on the right) a few years after their immigration. (Photo courtesy of Cory Streisinger).
They were not jumping in the great unknown, though. Margit’s two siblings, Fred and Viola have fled Hungary several years before, and they had already established a new life in New York. Fred worked for the General Oil Burner company, while Viola married a dry-cleaner, Anthony Burachio. Margit visited them in 1936, although probably she did not really expect to join them as an immigrant in a few years time.
The first to set foot in the New World was Ervin, freshly out of high school. He arrived on 8th October 1938, followed by his father on 25th November. Margit and George were held back for several months, most likely due to the difficulty to secure the necessary visas. (Being born in Losonc, at that time already part of Czechoslovakia, Andor Streisinger was under the jurisdiction of the Czechoslovakian travel quotas, whereas his Budapest-born wife and younger son had to wait for their turn within the Hungarian quota queues. Andor, with the help of Fred Freund, tried to lobby the American authorities several times, to issue the visa for Margit and Gyuri faster, but for little avail.)
Finally, on March 1939, the USS Manhattan arrived from Le Havre to Ellis Island, with the missing members of the family on board. The reunion, sadly, was not the happy meeting they had all hoped for. By this time Andor was quite sick, with metastasized gut cancer that spread to his spine. He was hospitalized the next day and in less than two months he died in the Long Island College Hospital.
The freshly widowed Margit Freund found herself with her two children in the middle of a huge and unknown metropolis. Erwin was able to look after himself, first as an employee in a tailor shop, but after a few years of college he enrolled into the Army. Margit and his younger son stayed for a few months with their relatives, and later moved to the 87th street. The mother, with the help of her sister found jobs as a dry cleaner and tailor. The housework was all left to young George.
The allurement of science
George mastered English extraordinarily fast, and just in a few years time, after taking one of the most competitive entrance exams in the city, he got enrolled into The Bronx High School of Science. Here, perhaps due to the earlier inspiration of the trips to the Buda mountains, he joined the Herpetology Club, which organized collecting trips to the Pine Barrens of New Jersey. His fondness for natural history was so obvious that he was soon hired by one of the fish experts of the New York Zoological Society, Dr. Myron Gordon. (The fish experience will become an important point of reference in Streisinger’s later life.) He relished working with Gordon, and due to his enthusiasm, after finishing high school, on the summer of ‘44 he got the opportunity of a lifetime: to work with Dobzhansky in the latter’s Columbia laboratory. The joint work resulted in the aforementioned paper, which was published in the Proceedings of the National Academy of Sciences.
Dobzhansky was fascinated about the possible role of sexual selection on the evolution of species, and the work of Streisinger was key to understand this. Young George has spent the summer with pairing males and females from different populations of a South-American Drosophila species to find out whether they attracted each other or not, by dissecting the females. As far as we can tell the renowned naturalist soon became more than a simple mentor for Streisinger, and he acted more like a father, taking the boy to his family holidays to California. But the most important influence on George’s life was his extended professional network: Dobzhansky helped Streisinger to gain access to Cold Springs Harbor, the cradle of modern molecular genetics, which became the defining moment in the young scientist’s life.
En early CV of George Streisinger. (Courtesy of Cory Streisinger.)
In the fall of 1944, aged only 16, Streisinger enrolled to Cornell University’s School of Agriculture. This might sound as an odd choice, but unlike other schools at Cornell the School of Agriculture was free to attend, with the small caveat that students had to spend some of their time with farmwork. And in the financially precarious situation faced by the Streisingers this was the only way George could actually enroll to the prestigious institution. Even this way he had to find other sources of income to get by: he worked regularly in the local coffee shops, and later tested the bacterial content of fresh milk in the Dairy Department. Ironically, the small caveat about the farmwork almost wrecked Streisinger’s career: while his classmates accomplished this work during the summer break, he was too busy toiling away in the laboratories of Cold Springs Harbor. And while this summer work helped him enormously in his later career, at Cornell he was forced to postpone his graduation and worked at a farm, shoveling manure during the coldest winter of the decade (something that made him proud even years later). Being a staunch pacifist, he also refused taking part in ROTC sessions, a decision that also contributed for his delayed graduation, but finally in the summer of 1950 he received his degree.
The Sielmans arrived to the US in 1938 also escaping the deportations of nazi Germany. Lotte and George found out soon that they had a lot of common, and after the first date they became an inseparable couple. They went together for bird watching in the morning, and in the spring their night rendez-vous often ended in the nearby marshes, where they observed the mating rituals of salamanders and frogs. They married in 1949, the day before Lotte’s graduation. A good friend was playing their favourite tunes on the bells of the chapel, and the wedding party was hold in a teacher’s garden, where an accordionist provided the music.
In the shadow of the bomb
The aftermath of the two atomic bombs, dropped after Germany’s capitulation (when most considered the war to be effectively over) caused serious soul-searching for many physicists working for the Manhattan project. Many considered the bombing unnecessary, a cynical exercise of power that was intended to send a message to the Soviet Union sacrificing tens of thousands. Many scientists turned away from physics for good and searched for other research topics. Influenced by Erwin Schrödinger’s essay, ‘What is Life?,’ for many of them biology would provide the next frontier. In his highly influential little book, Schrödinger tried to explain how simple physical and chemical reactions could explain the behavior of biological systems. One of the key problems, identified in the essay, was the nature of genetic information. No one understood at the time how this information was encoded on a molecular level (Schrödinger posited the existence of aperiodic crystals), and the book clearly catalyzed efforts to reveal this great unknown.
Answering the question required a new biological model, and under the influence of John von Neumann many opted to study the simplest model available, phages. (In a letter to Norbert Wienernek, the father of cybernetics, von Neumann pointed out that if scientists want to understand the information flow in living organisms, they need a simple model, such as a phage.)
Bacterial and phage genetics dominated the coming decades and delivered results (often followed by Nobel prizes) at an astonishing pace. Phages played a central role in demonstrating that DNA is the molecule of inheritance. Alfred Hershey’s and Martha Chase’s experiment complemented beautifully the bacterial work started by Frederick Griffith and finished by Oswald Avery. And while after Avery’s results there were still some who were adamant about the role of proteins in inheritance (DNA was considered just “too boring” to play such a central role in biology), the Hershey-Chase experiment silenced all the skeptics.
Hershey, together with two other colleagues, Salvador Luria and Max Delbrück (themselves immigrants, just like Streisinger), also founded the legendary “Phage group”. During the 1940s they organized summer courses at Cold Springs Harbor, and the list of attendees now reads as who-is-who for the molecular genetics of the 20th century. Seymour Benzer, Sydney Brenner and James Watson were amongst the first to come to CSH, as was Leo Szilard who also became interested in biology at the time. And amongst all these luminaries there was also George Streisinger.
Aaron Novick and Leo Szilárd (in the middle) during the 1953 Phage course. On the left, Max Delbrück is reading a newspaper, whereas on the right a young James Watson is sitting on the ground. (Photo courtesy of Cold Spring Harbor Library.)
Luria and Delbrück escaped the fascist and nazi regimes by emigrating to the US, where they soon became central figures of molecular biology. (Together with Hershey they received the Nobel prize in 1969 for their pioneering work). After a short detour at Columbia, in 1943 Luria started to work at the University of Indiana, where his first PhD student was Watson himself. The same year, in a series of inspired experiments, Luria and Delbrück demonstrated that mutations arise spontaneously in bacteria (something that was far from trivial in an era when many even questioned the existence of genes in prokaryotes).
The summer courses at CSH had a profound influence on the young Streisinger, and after finishing at Cornell, he started a PhD in Luria’s group. Probably equally importantly his friendships from the Phage group provided him with a fantastic social network for the rest of his life.
Phages
Streisinger followed Luria to the University of Illinois, where finally he could reunite with his wife. According to Lotte’s memoirs, this was also the time when George became involved in progressive political activism. He pursued this activism with a characteristic passionate rationalism till the end of his life. This was the time when he became deeply involved in the fight against desegregation – with motivated activist friends they provided evidence that in several neighbourhoods landlords choose renters based on skin color.
After receiving his PhD, George moved to California for a postdoc in Max Delbrück’s group. It wasn’t so much the research topic that attracted him to the West Coast as the realisation that everybody who counted in molecular genetics those days had gone through the Delbrück and Pauling labs at Caltech. During these three years (1953-1956) the family also expanded: soon after their arrival Streisinger’s first daughter, Lisa was born at the Pasadena Clinic, soon to be followed by Cory.
Streisinger’s research project at this time was to establish an experimental system for plant viruses, akin to the one that was already used for phages. The work progressed slowly, and in spite of the many experimental failures he established a fruitful barter system with his office roommate, Harry Rubin. Working with Rous’ Sarcoma Virus Rubin used chicken as his model, which complemented (in the kitchen, that is) very nicely the coconuts of Streisinger (used for their milk), thus for several weeks the Rubin and Streisinger families ate copious amounts of coconut chicken. This wasn’t the only extravagant gastronomical experiment, in which George took part at the time: during New Year’s Eve parties he cooked suckling pig in the departmental autoclave.
Finally, by the time everyone was fed up with chicken and coconuts, George, frustrated by the experimental failures, turned back to work again with phages. This led him to his first appointment, back at his alma mater, Cold Springs Harbor.
Thanks to the fame of the Phage group, by this time CSH has left behind the stigma of eugenics for good and became the Mecca of molecular biology. After the slow and quiet winter months the summer always brought in new people from all over the world, eager to collaborate with the local scientists, or to attend the Phage course. By this time, instead of the original Delbrück-Luria-Hershey trio, the course was taught by the resident researchers, including Streisinger himself.
The closing ceremony for one of the later Phage courses. Frank Stahl can be seen on the left and Geroge Streisinger on the right.
The three idyllic years at CSH were interrupted by a sabbatical year that the family spent in England. George was invited by Francis Crick to spend some time at the famed Cavendish Laboratories, together with Benzer and Brenner. Cambridge had a bubbling social scene at the time, with an endless number of programs and parties, often organized at the Crick’s home, “The Golden Helix”.
During their stay Benzer, Brenner and Streisinger came to agree that the English kitchen in the 1950s was awful. After his arrival Benzer suggested Crick that they should try out every day another restaurant, however soon they realised that in the absence of quality places the exercise was futile. Therefore, when a month later the freshly arrived Streisinger enthusiastically suggested the same thing, he was just met sarcastic laughs. But George did not give up the hope to eat quality food, so soon together with the Benzers and Renato Dulbecco they started a gourmet dinner-club. They went to great lengths to get the best local venisons – George once traveled to Yarmouth to by fresh clams for a dish of moules marinière. In the end the members of the dinner club were spending almost as much time with preparing meals as doing actual research.
Back from England Streisinger started looking for a new job, and the first offer came from Brandeis. It offered everything George needed, yet neither him, nor Lotte felt enthusiastic about the poshy Boston suburbs that were hard to square with the progressive worldview of the Streisingers.
During their sabbatical in Cambridge, the Streisingers toured the British countryside on a motorcycle with sidecar. (Photo courtesy of Cory Streisinger.)
However, while considering Brandeis an irrefutable offer came from Aaron Novick, the freshly appointed director of the Institute of Molecular Biology (IMB), in Oregon. Novick asked Streisinger to join him, to become one of the “Founding Fathers” of the new research center.
Novick graduated as a chemist in Chicago, where he was promptly recruited to the Manhattan project in 1943. This is where he met Leo Szilárd, who became soon a mentor for the young scientist. As for many fellow scientists, the detonation of the atomic bombs over Nagasaki and Hiroshima caused immense consternation and soul-searching for Novick. He wanted to get out of nuclear research as soon as possible and also became an ardent supporter of nuclear disarmament till the end of his life (for many years Novik edited the Bulletin of Atomic Scientists promoting a pacifist worldview).
The change in career was facilitated by Szilárd’s invitation to join him in attending the phage course at CSH, or in other words to join him for “an adventure in biology”. The opportunity excited Novick, who saw in it a chance “to exchange death for life”.
After the war Novick returned to the University of Chicago, where in 1959 he got the offer from University of Oregon to become the director of the freshly minted IMB (as the story goes, this was the first institution with “molecular biology” in its name). He also got a free hand in selecting his coworkers, and this is how he met Streisinger, who was suggested by a former colleague, Frank Stahl, himself a new IMB fellow.
Eugene, the home of IMB, and its surroundings enchanted the Streisingers who accepted the offer almost immediately. Soon they were settling into a small loghouse, close to the city. The new house and the nearby barn became the home to a number of animals as well, and George took great pleasure in his Nubian goat herd. This actually brought him fame, and soon he spent several weekends every year acting as a goat judge at different country fairs.
The idyllic life had a positive impact on George’s research as well. At IMB he continued his phage research, and his new results were getting the acclaim of his peers. This is demonstrated by a recommendation letter that Watson wrote for him when George was applying for a promotion: “George Streisinger is now one of the most productive geneticists in the United States. The work of his group at Eugene on the circular nature of the T4 chromosome is first rate. I wholeheartedly concur in your proposal to promote him to the rank of full professor.” The same esteem can be found in Streisinger’s correspondence with Francis Crick, where they discuss some of George’s more recent results that contributed to the deciphering of the genetic code, but also recognized that mutations at repetitive DNA sequencing are due to the stuttering of the polymerase, a phenomenon still recognized as ‘Streisinger slippage’ in the literature.
Watson’s recommendation letter for Streisinger. (Courtesy of Cold Spring Harbor Library.)
The more collaborative and much less competitive spirit of the era is also demonstrated by the fact that while Streisinger happily shared his data at conferences and private correspondence, he was much slower in writing them up into formal papers. This was the reason why at an IMB jamboree he was presented with “Doctor of Delayed Publications” honor. However, the greatest gap in his publication record in the 1970s was not due to his tendency to procrastinate, but because of his desire to “do something big”. And this was nothing less then establishing a new vertebrate model organism.
Fish
By the mid-1960s the phage as a genetic system showed signs of exhaustion. Most of the work instrumental in understanding the basics of gene function and regulation have been either already accomplished, or were close being so. Obviously there were still many questions to be answered, but many grandees of the phage field decided that they needed other models to properly examine these questions. As Sydney Brenner put it in a letter to Max Perutz, it was only a matter of time before “all the chemical details of replication and transcription will be elucidated. Because of this, I have long felt that the future of molecular biology lies in the extension of research to other fields of biology, notably development and the nervous system.“
Accordingly, some of the biggest names decided to leave the crowded phage-field and started pioneering work to find other model systems. Seymour Benzer started to study the nervous system of the fruitfly, whereas Brenner elevated a simple nematode, Caenorhabditis elegans, to the pantheon of genetic model organisms. Streisinger was also enthusiastic to change, but unlike his friends, he firmly believed that due their individual characteristics non-model organisms will be only of limited use in understanding and studying the complex nervous system and behaviour of vertebrates. To understand vertebrates, one needs a vertebrate model, and that is how zebrafish came into the picture.
Truth to be told, it wasn’t Streisinger who first used the zebrafish as an experimental organism. This honor most likely goes to a researcher at Wayne State University, Charles W. Creaser. Back in 1934 Creaser already argued that this little fish from Southeast Asia, a favourite of aquarists all over the world would be a suitable model to study embryology. Its small size, external fertilization and fast development make it an ideal model species for vertebrate developmental biologists. During the next couple of decades zebrafish research progressed at a slow space, with an occasional paper here and there. But at this time, it was only one of the many bony fish species that were used in research (and its relatives, goldfish and carp were far more popular). Nothing suggested that soon it would have a prominent role in biology.
Streisinger learned a lot about fish as an assistant of Myron Gordon, and due to that experience he became a hobby aquarist himself. Thus his choice of zebrafish might not seem too surprising. However, despite the fact the zebrafish was already used in lab experiments, the success was far from guaranteed. Due to the high risk of failure, at the beginning it was really hard for Streisinger to find colleagues who would join him in his new enterprise. Considering that it took many failures, and almost nine full years to get the system going, the critics might have had a point. (It is worth pointing out that probably the IMB lead by Novick was the only place at that time that would have had the patience and confidence to sustain Streisinger’s work for such a long time. But Novick not only personally supported George, but also convinced other IMB groups to help him out as they could.)
The university provided a World War II quonset hut to facilitate the new project. This is where Streisinger and his assistant, Charlene Walker worked out the standard conditions for maintaining and breeding zebrafish. As the hut became overheated during the summer, and too cold during the winter, the biggest challenge was to provide constant temperature. Against the heat they sprinkled water on the surface of the hut, whereas during winter dozens of electric heaters were installed (and often got short-circuited in the humid environment). Yet the problem of the temperature was easy compared with the headache that parasites caused to the experimental duo. These wiped out fish tanks again and again. In order to fight them, George and Charlene started to add more and more chemicals to the water. These restrained the parasites, but also interfered with fish development. Finally, after many trials and errors, Charlene was able to standardize the conditions, and George could start with the real experiments.
As he put it in a 1974 NSF application, his goal from the beginning was to “study features of the organization and embryological development of the vertebrate nervous system through the use of mutant strains.[He was] particularly interested in the mechanisms leading to the formation of specific synaptic connections and in the nature of the signals that guide specific axons to particular target sites.” He wanted to create mutants with nervous system abnormalities, but in order to create homozygous animals for rare mutations he had to overcome their diploidity.
A huge advantage of the worm, C. elegans, is that most of the individuals are hermaphrodites, capable of self-fertilization. This makes them well suited to study recessive mutations. Although hermaphroditism is almost nonexistent in vertebrates, there are quite a few examples for parthenogenesis, which also makes the analysis of mutations much easier. However, zebrafish is not one of the parthenogenic species. Streisinger, obviously, was aware of this, but he was convinced that he could find a way to jump start the development of unfertilized zebrafish eggs, thus in effect making them parthenogenic. After a few abandoned trials George and Charlene figure out the right conditions for the procedure, and they started producing mutants with vigour. The first results were published on the frontpage of Nature in 1981, and a new science was born: zebrafish genetics.
The first zebrafish clone aroused the interest of the general press, exemplified by the cartoon of the Chicago Tribune.
The popular press also enthused, but for different reasons: as the new fish became practically the clones of their mothers, soon Streisinger was inundated with questions about the ethics and politics of cloning. (The most unexpected admirers of the work were activist from the radical feminist groups of the 1970s. In their interpretation Streisinger’s work proved that the “male part of the conception process is irrelevant.)
In one of the many interviews he said: “We’ve developed a vertebrate system for doing interesting genetics. It just hadn’t occurred to us by any stretch of imagination that our work would generate any of this… There is a general worry among scientists, that overuse of cloning would lead to a loss of variety in food crops, but almost anything could be misused so it is very difficult to know at the time whether the good you get out of something will outweigh the bad.” He considered human cloning at the time unrealistic, but he also said that he had much greater, immediate worries. “I see the danger of global nuclear war imminent. The use of poison warfare, the widespread use of chemicals that may be hazardous, the lack of any serious attempt to deal with population growth, the lack of any real concern about just incredibly unequal distribution of wealth.” These were all things that should have been more important for newspapers than zebrafish cloning.
After the University of Illinois years Streisinger never stopped to be civil rights activist, and the atmosphere at IMB was very supportive for this sort of activities. Novick’s campaign against nuclear proliferation was only the most prominent of the campaigns originating within the walls of the institute. IMB researchers became important allies of EPIC (Eugene Peace Information Center), and none of them was as vocal as Streisinger himself. He became the local representative of the national Turn Toward Peace lobby group, but his major contribution came in the fight against toxic herbicides.
In 1962 Rachel Carson’s “Silent Spring” successfully focused the attention of the public to the potential health hazards of agricultural chemicals. The book that ushered the creation of EPA, was drawing attention to the hazard of DDT, but it was obvious from the start that there are many other potentially dangerous pesticides. In the Oregon woods the local Forest Service used dioxin-based chemicals against weeds, and this unsettled the locals. Many of them were afraid that these chemicals (all related to the infamous Agent Orange) could accumulate in the plants and get into the groundwater. EPA originally concluded that there was no reason to worry, but many were unconvinced. They asked IMB researcher to help to prove that dioxin is toxic and dangerous. Streisinger became the star witness of the trial, and argued successfully that in the case of dioxin there is no such thing as “safe dosage”, as even at very low concentrations that chemical causes chromosome breakage, thus it must be mutagenic. The judge accepted this, and ordered the EPA to perform new, systematic studies. These, in turn, lead to the banning of dioxin-based herbicides in a few years time.
Streisinger’s opinion carried weight, because as a geneticist he studied often the mutagenic potential of different molecules. These observations were important not merely as toxicological observations (although, undoubtedly, zebrafish is still an important model in toxicology). George wanted to understand how mutagens work, so he could use them in his project to create mutant fish lines.
One of the last photos of George and Erwin Streisinger was taken in 1981 on the porch of George’s loghouse. (Photo courtesy of Cory Streisinger.)
Besides research and civic duties, George took teaching extremely seriously as well. Usually he started his lectures at 8 am, so he could do experiments the rest of the day. But even after several decades of experience he still worried how the students will receive the material, thus in order to get well prepared, he woke up at 4. His methods of teaching were way ahead of his time: one could easily think the Dance your PhD competition is something original, a creative 21st century idea, yet four decades ago Streisinger collaborated with the local Dance Department to illustrate his lecture about protein synthesis.
By the early 1980s as the founder of a zebrafish research, Streisinger was becoming one of the star researchers of IMB. His efforts were not only recognized by his peers (by this time many other IMB groups started to work with fish), but also by the wider public of Oregon. However, as the IMB was preparing to celebrate it’s 25th birthday a tragedy happened: on 11th August 1984, a few weeks before the ceremony Streisinger died during a scuba diving exam near the Oregonian coast (that old heart murmur never disappeared completely). The IMB birthday promptly became a Streisinger memorial. It was attended by the very best of the 20th century molecular biology. From Hershey to Watson they all wanted to pay their tributes.
Without its father, the future of zebrafish research became in peril. It was up to the IMB colleagues to carry the torch, and they promptly did so. Thanks to their work as well, today it is obvious that zebrafish fulfilled the expectations. It became one of the most widely used genetic model organisms, used as much to study simple developmental processes as to model complex human disease. But the thousand or so zebrafish labs around the world today exist only because of that one quonset hut in Eugene, where George and Charlene heroically worked out the basics of zebrafish research.
Epilogue
Although Streisinger emigrated at a very young age, he never completely forgot where he came from. The recipes of Hungarian pastries were always held in high esteem in the family cookbook, and with Margit and Erwin George they always spoke Hungarian (after many decades he still spoke English with a characteristic Hungarian accent). Yet, we know of a single occasion when he visited Hungary after 1939. In the 1960s he arrived to Budapest for a lecture, together with Frank Stahl. The visit was a good opportunity to revisit his city of birth and meet some relatives. As Frank Stahl remembers, they walked a lot on the banks of the Danube, ate in small restaurants, and Streisinger was fond of recalling his memories of Gellért bath. At the lecture George was adamant to speak in Hungarian, but probably his mixed accent and the topic that was completely novel to his audience (biology in the Eastern bloc was still under the influence of lysenkoism) just confused the people in the room. So in a few minutes time he was asked to continue in English.
This is an edited version of a Hungarian language text that appeared last year. We are republishing it because George Streisinger is such an important person for the zebrafish community – and next year we want to celebrate his 90th anniversary at the 10th European Zebrafish Meeting. (If you are interested in zebrafish genetics, please come!)
To write this post I relied heavily on Lotte Streisinger’s book “From the sidelines”, Jim Endersby’s “A Guinea Pig’s History of Biology” and Errol Friedberg’s “Sydney Brenner – A biography”. Some facts were also taken from George Streisinger’s autobiographical essay, “Terminal Redundancy: All’s Well That Ends Well”. I am extremely grateful to Cory Streisinger for sharing the countless family stories and photos. I also want to thank Frank Stahl, Ferenc Müller and Julianna Vig for helping me with this story. Photos are used with the permission of the Streisinger family and Cold Spring Harbor Library.
The Paridaen lab at the European Research Institute for the Biology of Ageing (ERIBA) in Groningen, The Netherlands, is looking for a PhD student on Stem cell biology.
Stem cells act in developing and adult organisms to produce the proper number of specialized cells in the body. It is important that stem cells select an appropriate division mode to ensure proper development, maintenance and repair of tissues. In the Paridaen lab, we aim to understand the mechanisms underlying stem cell decisions in developing and ageing tissues. We employ single cell analyses techniques to study stem cell behaviour in the developing brain, using zebrafish as our main model system. In particular, we are interested in the role of fate-determining factors such as signalling pathways and subcellular structures in symmetric and asymmetric stem cell divisions.
ERIBA is a newly established institute at the University Medical Centre Groningen (UMCG). The mission of ERIBA is to better understand the molecular mechanisms that result in age-related decline and age-related diseases. We aim to develop novel strategies to prevent or combat age-related diseases and to provide evidence-based recommendations for healthy ageing. The research teams are working with different model systems and technology platforms and share their knowledge and expertise to accelerate discoveries. The ERIBA is housed in a new building and is part of the central UMCG campus in the vibrant University City Groningen in the north of the Netherlands.
Fertilization marks the start of life. This is followed by highly coordinated epigenetic reprogramming that allows protamine-histone exchange, zygotic genome activation, and the generation of a totipotent embryo. However, the true state of chromatin at the level of DNA during this crucial period is a long-standing mystery.
Our lab is dedicated to understanding epigenetic reprogramming in early development. But for a long time, we were struggling to find the right tools. When I first heard about ATAC-seq developed by the Greenleaf and Chang groups in Stanford, I was very excited as this is exactly the approach that we were looking for. I brought this up to my PI and we quickly decided to give it a try. We started by testing it in various numbers of mESCs, using either crosslinked or native chromatin from either frozen or fresh samples as well as different kinds of detergent in lysis buffer. In our hands, it appeared that ATAC-seq worked best on native chromatin of fresh samples from 100 to 1,000 cells with 0.15% to 0.5% NP-40 in lysis buffer.
We then began to apply ATAC-seq to mouse early embryos. Working together with another graduate student Bo Huang who has extensive experience in mouse embryos, we collected the early 2-cell, 2-cell, 4-cell, and 8-cell embryos, as well as ICMs isolated from blastocysts. However, we were surprised to see that the sequencing libraries were strongly contaminated by mitochondrial DNA, which occupied up to 99% of the reads. This is mainly due to the large quantities of mitochondria inherited from oocytes. My colleagues and I then spent almost a year to test several approaches to reduce mitochondrial contaminations. Most methods we tried did not meet our expectation, until we developed CARM, the Cas9-Assisted Removal of mitochondrial DNA. The 114 sgRNA evenly targeting 16kbp mitochondrion genome is designed to introduce the cutting event on mitochondrion DNA fragment in ATAC-seq library, so that these reads could not be sequenced due to the lack of sequencing adaptor on the both end of the fragments. We were very happy to see that CARM significantly reduced the about 70% of mitochondrial DNA at that time. Developing a new method is never easy, but the support from my mentor enabled me to proceed without any hesitation. He always encourages us to take the challenge to solve the problem that matters.
This story is now published on Nature as an article in June 30 (Wu & Huang et al., Nature 534 (652-657)). There are several interesting discoveries that we have made in this study.
Schematic of ATAC-seq and CARM for probing accessible chromatin in mouse preimplantation embryos
Firstly, we found that unlike asymmetric DNA methylomes between the two alleles throughout preimplantation, accessible chromatin landscapes already become comparable between the two parental alleles after the 2-cell stage. Secondly, the open chromatin in early embryos occurs not only at the transcription start sites (TSSs) but also at the transcription end sites (TESs) of active genes, indicating a unique and unknown regulation mechanism in early development. Thirdly, we identified putative enhancers that are active in early development, through which we derived candidate transcription factors that may regulate early development. We validated the roles of two such regulators, Gata4 and Nr5a2, in lineage-specific transcription programs in preimplantation development. Finally, in early 2-cell embryos where major ZGA has not started, we found many large open chromatin domains (up to 110kb) over regions showing repeat-driven promiscuous transcription. These data support an unusually permissive chromatin state at this stage.
Taken together, our findings unveiled highly dynamic spatiotemporal configuration of chromatin states in early mammalian development. However, we believe this is just the beginning. Future investigations are warranted to fully unlock the mechanisms and the functions of these unique regulatory modes. As a graduate student, this was truly an exciting journey for me. The happiest thing is to make discoveries in a world full of unknowns.
Here are the highlights from the current issue of Development:
Gestational stress: at the heart of birth defects
Congenital heart disease (CHD) is the most common form of human birth defect, yet the genetic and environmental factors that contribute to CHD remain poorly understood. Here, Sally Dunwoodie and colleagues investigate how gestational hypoxia affects heart development in mouse embryos (p. 2561). They reveal that the exposure of developing mouse embryos to short-term hypoxia in uteroresults in heart defects, notably perturbations to the outflow tract (OFT). These changes are mediated by altered cell proliferation and FGF signalling in the secondary heart field (SHF), which contains progenitor cells that contribute to the OFT. The authors further report that hypoxia leads to rapid induction of the unfolded protein response (UPR) in SHF cells. This, in turn, causes a global decrease in protein translation and may contribute to the reduced levels of FGFR1, and hence FGF signalling, observed in SHF cells following exposure to hypoxia. Together, these results suggest that hypoxia-mediated UPR induction during pregnancy can give rise to CHD. Given the key role of FGF signalling during embryogenesis, these findings also have important implications for understanding birth defects that affect other organs.
Shox2 goes out on a limb
Hox-TALE factors are involved in patterning the vertebrate limb but precisely how they regulate specific and regional gene expression patterns is unclear. Here, on p. 2548, YiPing Chen and co-workers uncover a limb patterning transcriptional programme that is coordinated by the transcription factor Shox2. The researchers demonstrate that, although Shox2 is expressed in mesenchymal progenitors of multiple cell types in the proximal limb, its deletion specifically in the osteogenic lineage causes limb defects and loss of the stylopod – the most proximal region of the limb. ChIP-Seq analyses indicate that Shox2 binds predominantly to limb-specific enhancers that are involved in skeletogenesis; these regions are also co-occupied by Hox-TALE factors. Finally, the authors show that Shox2 is expressed in a gradient that is complementary to that of TALE factors and that it represses the expression of TALE factors in the stylopod. Overall, these observations, together with other findings, highlight the existence of a Shox2-coordinated transcriptional programme that functions to pattern the vertebrate limb and provide insights into the ‘enhancer grammar’ that is used to mediate specific transcriptional outputs.
Pinning down spindle orientation
Spindle orientation is regulated by a conserved mechanism in which Pins/LGN anchors Mud/NuMA to the cell cortex. This pathway has been assumed to operate in nearly all animal cell types but here (p. 2573) Daniel St Johnston and colleagues reveal that Pins is not required for spindle orientation in the Drosophila wing disc. Using live imaging, they first discover that spindle angles in the wing disc vary widely; spindle angles are initially random but gradually align with the plane of the tissue as cells enter anaphase, highlighting that spindle angles are not accurate predictors of division orientation until this point. Importantly, the researchers reveal that spindle orientation does not require Pins, or aPKC, Dlg or Lgl. They further report that Mud is able to localize to the cell cortex in the absence of Pins, suggesting that a parallel but as yet unknown mechanism must act to localize Mud in wing disc cells. In summary, these surprising results indicate that a Pins-independent mechanism can orient the mitotic spindle in the Drosophila wing disc and lead the authors to propose that this system provides robustness to this rapidly developing epithelial tissue.
A sticky situation in plants
Cell-cell adhesion in plants is known to be regulated by pectins, and levels of homogalacturonan (HG; the main component of pectins) within the cell wall have generally been linked to cell adhesion. But how is cell-cell adhesion maintained and regulated in the face of the dynamic cell wall remodelling that takes place during cell growth and division? Here, Grégory Mouille and colleagues investigate this issue (p. 2536). Using a cell adhesion defect suppressor screen, they identify a putative O-fucosyltransferase – an enzyme that mediates the transfer of sugar residues onto substrates – that regulates cell adhesion in Arabidopsis thaliana. They further reveal that mutations in the gene encoding this enzyme or another putative O-fucosyltransferase perturb cell adhesion. Importantly, a comparison of mutant and suppressor lines suggests that cell adhesion does not rely on HG content per se. Based on their findings, the authors propose a model in which a pectin-related signalling pathway, rather than simply HG levels, contributes to the control and maintenance of cell adhesion during plant growth and development.
Plus:
An interview with Enrico Coen
Enrico Coen CBE FRS is a Project Leader at the John Innes Centre in Norwich, UK, who uses a variety of approaches to study patterning and morphogenesis in plants. We met with Enrico at the Spring Meeting of the British Society for Developmental Biology, where he was awarded the Waddington Medal, to ask him more about his career and his passion for art and book-writing. See the Spotlight article on p. 2479.
Exosomes in developmental signalling
Cells can signal by releasing exosomes – extracellular vesicles containing bioactive molecules such as RNA, DNA and enzymes. Recent work has suggested that exosomes can also carry signalling proteins, including ligands of the Notch receptor and secreted proteins of the Hedgehog and WNT families. Here, Ian McGough andJean-Paul Vincent describe various types of exosomes and their biogenesis and critically assess the role of exosomes in developmental signalling. See the Review on p. 2482.
Direct neuronal reprogramming: learning from and for development
The key signalling pathways and transcriptional programmes that instruct neuronal diversity during development have largely been identified. Here, Magdalena Götz and co-workers discuss how this knowledge has been used to successfully reprogramme various cell types into distinct types of functional neurons, and the extent to which direct neuronal reprogramming recapitulates embryonic development. See the Review on p. 2494.
Axons in mature nervous systems regenerate poorly after injury, creating a major obstacle for recovery from neuronal injury. We know, however, that their regenerative capabilities are affected by both cell-extrinsic and intrinsic factors. Understanding these processes is crucial to provide future therapeutic intervention for neuronal regeneration. We recently found a cell-intrinsic factor inhibiting axon regeneration in a large-scale genetic screen. Several lines of evidence suggest that the activity of this factor is regulated by phosphorylation. Having spent a year as a post-doc in UC San Diego, working on the genetic side of the project, I decided to make a collaborative visit to the laboratory of Dr. Fumio Motegi (Temasek Life Sciences Laboratory, Singapore) to biochemically test our hypothesis.
Temasek Life Sciences Laboratory
I am grateful to the journal Development and Company of Biologists for the generous support to make the trip possible. I had a successful trip to Singapore and have some great memories working in a friendly lab in Singapore. With very kind guidance and help from the lab members, I have learnt several useful techniques such as in vitro protein purification and in vitro kinase assay. Together, we have tested several kinase candidates and are starting to have some clues on how we should advance in this project.
The Motegi lab
This trip has also allowed me to make new connections with different research groups in Singapore. During my visit in Singapore, I presented my work in the Singapore Worm Club and attended a number of seminars to get to know what other groups are working on. I also visited several institutes in Singapore, including the Mechanobiology Institute and Lee Kong Chian School of Medicine. I was very impressed with the high standard of research conducted and outstanding facilities provided in these institutes. Importantly, by sharing my research work with others, I have found a few interesting people who may turn out into close collaborators in the future.
In conclusion, this travel fellowship has given me the chance to experience research in Singapore. I have not only learnt techniques that are useful for my project, but have also made many new friends in Singapore. I would like to encourage other young scientists to look out for opportunities to experience research in different countries. Finally, I would like to thank the Company of Biologists for awarding me the fellowship.
This week I attended a meeting with Paul Nurse, director of the The Francis Crick Institute, who spoke about the potential implications of the pending Brexit on scientific research at the Crick. Like many other postdocs, I never envisaged that I would be sitting in the Fletcher Hall at Mill Hill following a referendum outcome that meant the UK would eventually leave the EU….but that is what happened, and here we are. So what happens next?
The Crick was implemented to facilitate “Discoveries Without Boundaries”. Behind this mantra, the organisation thrives on the exchange of people, infrastructure and funding – elements that support individuals to reach their full potential, which in turn, facilitates the best possible research outcomes. At a postdoctoral level, 56% of scientists at the Crick are EU nationals (myself included). In addition, the Crick receives very generous funding support from EU sources such as the ERC and Marie Curie grant schemes. This support currently amounts to 5 million pounds per annum – a figure that had been projected to rise to 10 million, as the Crick becomes fully operational in central London.
Science requires vast sums of money – a fact that one can hardly ignore, especially when the UK benefits from contributing less to the EU than it receives in return (see the report from the Royal Society). A deeper concern, however, is that it is not yet clear what support UK scientists could expect to receive from the government moving forward (see related post “People in this country have had enough of experts”). It remains to be seen what mechanisms will be put into place, and how these will be managed into the future. But the very essence of what science is all about (education, innovation, discovery, just to name a basic few), culminates in multifaceted ways that can enhance quality of life. How long can this be ignored by Westminster politics??? If we want a “country that works for everyone”, the new buzz words on the lips of newly appointed PM Theresa May, we simply can’t have our cake and eat it too.
Although Brexit has come as a terrible shock to almost everyone I know, and no matter the perceived restrictions, the actual situation is that UK science currently represents a thriving and highly diverse workforce (the Crick alone employs people from over 70 different nations). Working in this environment is exciting, challenging and highly rewarding. While Brexit was not an outcome favoured by most scientists, including Paul Nurse, it doesn’t change the fact that I am currently surrounded by some of the world’s most talented individuals. That and the fact that I’m not ashamed to admit, in the words of Dr Who,
“I am and always will be the optimist. The hoper of far-flung hopes and the dreamer of improbable dreams”.
As the whole is greater than the sum of its parts, I remain hopeful that as a scientific community, although we move forward with many variables left unknown, we can still strive to make valuable contributions to shape the future of UK science. This is vital to ensure the continued prosperity of scientific pursuits and to minimise the impact of the challenges that lie ahead.
Amelia Lane, David A. Parfitt, Conor M. Ramsden, Peter J. Coffey, Michael E. Cheetham
Parfitt et al. Cell Stem Cell 1–13 (2016)
Making eyes
Methods to differentiate human stem cells into retinal cell types have been under development for almost a decade. Stem cell derived retinal cells provide a rich resource to study unique and otherwise inaccessible cells. Reprogramming somatic cells into iPSC allows us to study retinal cells carrying pathogenic mutations that cause retinal degeneration. This enables the interrogation of disease mechanisms and allows us to screen mutation-specific therapies such as antisense oligonucleotides on patient cells. The substantial technical challenges of consistently and uniformly generating terminally differentiated photoreceptors are gradually being overcome allowing us to make use of this new and powerful experimental paradigm.
The retina is derived from the anterior neural plate. During vertebrate eye development a common pool of progenitor cells invaginate to form a double walled optic cup. The cup is patterned by interactions with the adjacent surface ectoderm to form neural retina (NR) and an overlying monolayer of pigmented cells known as retinal pigment epithelium (RPE) (Fuhrmann et al., 2000). Mature photoreceptors in the human retina have a unique structure suited to their purpose as sensors that transduce light into an electrochemical signal. Rod and cone cells have distinct compartments. Their cell bodies are arranged in an outer nuclear layer (ONL), consisting of tightly packed columns approximately 10 nuclei in thickness. At the apical edge they have tight junctions forming an outer limiting membrane (OLM). Above the OLM are inner-segments (IS), which house much of the ER and Golgi and are densely packed with mitochondria. Protruding from the IS of each photoreceptor cell is a specialised cilium, the outer segment (OS), a 25um stack of membranous discs packed with photosensitive opsin proteins and the phototransduction machinery. The OS is joined to the IS by a transition zone referred to as the connecting cilium (CC).
The first differentiated retinal cell type to be derived from human embryonic stem cells (hESC) were RPE, the cells which overlay the photoreceptors in the retina. Pigmented RPE were first detected following spontaneous differentiation of confluent hESC cells (Klimanskaya et al 2004). It has since been demonstrated by several labs around the world that induced pluripotent stem cells (iPSC) and hESC will spontaneously differentiate into RPE within a month. Making photoreceptors requires longer periods of time similar to the process in utero. In the first published attempts at photoreceptor differentiation, hESC cells were treated with Wnt and nodal inhibitors followed by long term treatment (more than 130 days) with retinoic acid and taurine (Osakada et al., 2008, Lamba et al., 2006). These protocols generated immature photoreceptor progenitor cells and/or opsin-expressing cells with no specific morphological resemblance to photoreceptors in a two-dimensional format.
Pioneering work by the Sasai lab at the RIKEN Centre for Developmental Biology in Japan, using a three dimensional ‘organoid’ approach for cell differentiation, greatly improved the quality of the photoreceptor cells derived. First with mouse, then with hESCs, Sasai’s lab was able to create photoreceptors from embryoid bodies (EBs) that formed three-dimensional (3D) ‘optic cups’ with RPE and several neural retina layers (Eiraku et al., 2011, Nakano et al., 2012). This was rapidly followed by similar demonstrations in human iPSC (Zhong et al., 2014, Reichman et al., 2014). These opsin-expressing photoreceptor cells developed mitochondria-rich inner segments with cilia easily observed, raising the tantalising possibility that they may be capable of generating OS in vitro.
Figure 1. Stages of iPSC-photoreceptor differentiation. Bi-laminated neuroepithelium (sox2, nestin) develops a layer of recoverin expressing photoreceptor progenitors which mature into rhodopsin expressing photoreceptors.
Disease modelling
Our lab’s research interest in inherited retinal dystrophy disease mechanisms prompted us to attempt to make 3D retinas from cells donated by a patient with a type of retinal degeneration known as Leber congenital amaurosis (LCA). LCA is a recessively inherited retinal dystrophy resulting in severe visual loss in early childhood. This patient is homozygous for a deep intronic mutation in the gene encoding CEP290 (c.2991+1665A>G), which is the most common LCA associated allele. CEP290 is found at the transition zone of all ciliated cells and in the CC of photoreceptors. Its position indicates a role in formation of the Y-shaped linker structures that anchor the ciliary axoneme to the plasma membrane. Mutations in CEP290 can cause syndromic ciliopathy involving several organs or LCA alone. The common c.2991+1665A>G mutation is only associated with LCA and leads to mis-splicing of the CEP290 transcript and inclusion of the stop-codon containing cryptic exon. Some correctly spliced transcript remains and it is thought that the residual protein level is sufficient to prevent a syndromic ciliopathy in affected individuals. Fibroblasts and lymphoblast cultures derived from these patients display a cilia defect; with reduced numbers of shortened cilia (Collin et al., 2012, Gerrard et al., 2012, Garanto et al., 2016). In patients however, it is not clear why retinal cells appear to be affected more than other tissues.
Our first attempts at the 3D organoid method used embryonic stem cells (ESC) and control iPSC derived from neonatal foreskin fibroblasts. The ESC EBs generated very promising bi-laminated optic cups with retinal ganglion and neuroblastic cell layers as well as RPE. The neuroblastic layer contained photoreceptor progenitors, but we did not continue the differentiation for long enough to generate mature photoreceptors with IS.
We then began large scale and long-term differentiations using both control and CEP290 patient derived iPSC lines. We tried both Zhong and Nakano protocols from several patient and control lines and carried out immunocytochemistry (ICC) and gene expression analysis to follow the time course of photoreceptor differentiation.
CEP290 patient fibroblasts reprogrammed efficiently into iPSC. Similar to the fibroblast cells, there was a significant reduction in ciliation in iPSC; however, this did not appear to affect their pluripotency. CEP290 iPSC efficiently differentiated into early derivatives of all three germ layers. We found that different clonal lines from both control and patient iPSC formed EBs with varying efficiency (50-100%). Of these, a proportion could be seen to generate transparent, radially aligned neuroepithelium (NE) – which again varied between lines (5-30%). We were able to distinguish these NE-producing EBs morphologically with a light microscope and dissect out the neuroepithelial buds (Figure 1). Unfortunately, a proportion of these did not survive the dissection procedure or spontaneously collapsed and became necrotic. However, the precious surviving organoids were cultured for up to 21 weeks.
We processed EBs early on in the differentiation process and saw that the basal bodies and associated cilia had aligned at the apical surface of the EBs with neuroepithelial domains, thereby fulfilling their roles as sensors that detect and relay signals from the outside environment, and facilitating polarised cell division. Excitingly we detected a phenotype in our CEP290 EBs – significantly fewer of the pericentrin positive basal bodies at the apical surface possessed ARL13B positive cilia. This did not appear to reduce the efficiency with which CEP290 EBs generated organised neuroepithelial domains; an interesting observation given the role of primary cilia in embryonic forebrain development (Willaredt et al., 2013).
We extracted RNA from individual EBs and analysed gene expression as well as cryosectioning them for ICC analysis. As the EBs developed it became clear that the differentiating cells were following the highly conserved sequence of eye development; similar to what has been observed in other studies of iPSC-retina differentiation (Meyer et al., 2009). Various photoreceptor cell markers such as recoverin and arrestin were being switched on over time (Figure 1). Comparing the time course over which markers were expressed it was clear that the patient cells were behaving similarly to controls and our model did not display any ‘developmental’ defects in vitro.
We used electron microscopy to analyse the ultrastructure of these developing photoreceptors; at week 13 tight junctions were visible all across the apical layer forming an OLM and above this mitochondria rich buds had appeared, with the morphology of inner segments.
At the latest time point (21 weeks), ICC revealed the presence of rhodopsin positive rods (Figure 2), as well as red/green and blue cone cells . These were arranged within a compacted apical layer, reminiscent of the ONL and approximately 5 nuclei thick. By this time, the interior of the cups was largely empty or necrotic. RPE cells could also be seen in several of the cups but it was generally adjacent to the photoreceptors cells and very rarely overlaying them as in the 2 walled optic cup seen in human development and reported by Sasai’s lab (Nakano et al., 2012).
Looking at this latest time point by electron microscopy we were astonished to see the presence of connecting cilia that were elaborating into partially stacked discs similar to rudimentary OS. The discs were aligned at the base but disorganised and broken at the tip. In vivo the outer segments are enveloped by the microvilli of the RPE and encased in an extracellular matrix with a unique composition known at the interphotoreceptor matrix (IPM) (Ishikawa et al., 2015). This proteoglycan-rich substance is secreted by both the RPE and the photoreceptors in vivo and, in contrast to the majority of ECM in other tissues, is lacking in a collagenous meshwork with neither laminin nor fibronectin as major components. We observed a lot of broken cilia and opsin positive debris at the apical edge and hypothesise that if we could place the developing photoreceptors in an environment that would stabilise the nascent OS, akin to the IPM, we might observe more organised and abundant OS structures. This could be a powerful tool for studying the numerous mutations in retinal dystrophy genes that lead to OS trafficking disorders and retinal degeneration.
There was a reduction in cilia number in CEP290 patient optic cups using ARL13B to mark the connecting cilia of opsin positive photoreceptors. In addition the connecting cilia were now significantly shorter than those in controls. However the most interesting insight came from looking at the relative levels of mis-splicing in the various cell types derived. While the ratio of correctly spliced: mis-spliced transcript in fibroblast, iPSC and iPSC-RPE was approximately 50:50, in photoreceptors the quantity of correctly spliced transcript was severely reduced down to only 10-20%. Cell type specific levels of mis-splicing could explain the specificity of retinal involvement characteristic of this particular mutation, with less full-length CEP290 protein in the retina than in other tissues. The reasons for this different mRNA processing are currently unclear, but it was recently shown that the human retina has unexpectedly high levels of splicing diversity (Farkas et al., 2013) and mutations in splicing factors can cause dominant retinal dystrophy. This finding raises the possibility that the retina could be especially sensitive to intronic variants that affect splicing.
Figure 2. iPSC derived photoreceptors develop rudimentary outer segments (OS) which is joined to the inner segment by the connecting cilia (CC), the location of many ciliopathy proteins such as CEP290.
Drug testing
The ability to make human retinal cells in vitro opens up lots of possibilities to study human development, disease mechanisms and test novel therapies. Antisense oligonucleotides (AON) that bind to and sterically block the aberrant splice donor site in the mutated CEP290 pre-mRNA have been developed and tested in patient fibroblasts (Collin et al., 2012 Gerrard et al., 2012). We were able to show that 10mm of an antisense morpholino (MO) specific to the LCA CEP290 mutation (CEP290-MO) transfected in EndoPorter solution (from Genetools) reduced the amount of cryptic exon containing-mRNA produced and increased ciliation in patient fibroblasts. We also showed that the treatment lasts up to 6 days.
Testing this potent MO on the CEP290 optic cups was the next logical step. In order to assess the capacity of the MO to enter the optic cup we used a fluorescein-tagged MO and imaged all layers of the cup 48 hours later using live confocal microscopy. The tagged MO was able to penetrate all layers of the optic cup and access the nuclei. Therefore, we treated 90-day-old patient optic cups with CEP290-MO every 3-4 days for 4 weeks and then assessed cilia length, cryptic exon expression, CEP290 levels and the amount of various CEP290 interaction partners at the basal body. CEP290-MO treatment significantly increased CEP290 protein levels, reduced cryptic exon expression and all but restored cilia numbers and length to wild type levels. Critically it also restored the localisation of the important CEP290 interacting partner, RPGR, to the connecting cilia. RPGR is located in the same ciliary compartment as CEP290 and mutations in RPGR are a major cause of retinitis pigmentosa (Breuer et al., 2002). We used high magnification confocal stacks to compare RPGR and CEP290 fluorescence at the ciliary base. We observed a significant depletion of RPGR and CEP290 in patient optic cups and a significant restoration following CEP290-MO treatment, an important functional read-out for the treatment efficacy.
In the absence of an animal model that accurately recapitulates the mis-splicing mutation, the patient iPSC-photoreceptor technique has yielded unique and important mechanistic insights and provided an explanation as to why this most common CEP290 mutation leads to a retina-only phenotype in human cells. In addition we have demonstrated the efficacy of AONs in rescuing correctly spliced CEP290 levels. Hopefully these studies will encourage the use of iPSC-derived organoids to study disease mechanisms and facilitate the development and clinical application of new therapies.
Breuer DK, Yashar BM, Filippova E, et al. A Comprehensive Mutation Analysis of RP2 and RPGR in a North American Cohort of Families with X-Linked Retinitis Pigmentosa. American Journal of Human Genetics. 70(6),1545-1554 (2002),
Collin, R. W. et al. Antisense Oligonucleotide (AON)-based Therapy for Leber Congenital Amaurosis Caused by a Frequent Mutation in CEP290. Mol. Ther. Nucleic Acids1, e14 (2012).
Eiraku, M. et al. Self-organizing optic-cup morphogenesis in three-dimensional culture. Nature472, 51–56 (2011).
Farkas, M. H. et al. Transcriptome analyses of the human retina identify unprecedented transcript diversity and 3.5 Mb of novel transcribed sequence via significant alternative splicing and novel genes. BMC Genomics14, 486 (2013).
Fuhrmann, S., Levine, E. M. & Reh, T. a. Extraocular mesenchyme patterns the optic vesicle during early eye development in the embryonic chick. Development127, 4599–609 (2000).
Garanto, A. et al. In vitro and in vivo rescue of aberrant splicing in CEP290 -associated LCA by antisense oligonucleotide delivery. Hum. Mol. Genet. (2016).
Gerard, X. et al. AON-mediated Exon Skipping Restores Ciliation in Fibroblasts Harboring the Common Leber Congenital Amaurosis CEP290 Mutation. Mol. Ther. Nucleic Acids1, e29 (2012).
Ishikawa, M., Sawada, Y. & Yoshitomi, T. Structure and function of the interphotoreceptor matrix surrounding retinal photoreceptor cells. Exp. Eye Res.133, 3–18 (2015).
Klimanskaya, I. et al. Derivation and Comparative Assessment of Retinal Stem Cells Using Transcriptomics. Cloning Stem Cells6, 217-245 (2004).
Lamba, D. a, Karl, M. O., Ware, C. B. & Reh, T. a. Efficient generation of retinal progenitor cells from human embryonic stem cells. Proc. Natl. Acad. Sci. U. S. A.103, 12769–74 (2006).
Meyer, J. S. et al. Modeling early retinal development with human embryonic and induced pluripotent stem cells. Proc. Natl. Acad. Sci. U. S. A.106, 16698–703 (2009).
Nakano, T. et al. Self-formation of optic cups and storable stratified neural retina from human ESCs. Cell Stem Cell10, 771–85 (2012).
Osakada, F. et al. Toward the generation of rod and cone photoreceptors from mouse, monkey and human embryonic stem cells. Nat. Biotechnol.26, 215–24 (2008).
Willaredt, M. A., Tasouri, E. & Tucker, K. L. Primary Cilia and Brain Development. Mech. Dev.130, 373–380 (2013).
I am Yoshimasa Hamada, a Research Fellow in Okayama University Graduate School in Japan, working with Prof. Kenji Tomioka, Prof. Hideyo Ohuchi, Prof. Sumihare Noji and Dr. Tetsuya Bando. Our research focuses on the molecular mechanisms underlying leg regeneration, embryonic development, and circadian rhythm using the two-spotted cricket, Gryllus bimaculatus (Figure 1).
The two-spotted cricket, Gryllus bimaculatus
The two-spotted cricket Gryllus bimaculatus (De Geer) was newly described by Baron Charles de Geer in 1773. The latin name Gryllus means cricket, and bi-maculatus means two-spots, because Gryllus bimaculatus has a white spot in each forewing. The cricket has a global distribution, being found in European, African and Asian countries. Gryllus bimaculatus (and the house cricket Acheta domesticus) is a popular food source of meat-eating arthropods, amphibians, reptiles and other pet animals. Hence, we easily buy the crickets from a pet shop at a low price (we can buy 5~10 crickets for £1). Interestingly, the cricket will be a diet even for us as well as for pets: the Food and Agriculture Organization of the United Nations has proposed that insects would be a good food source for human. In fact, BBQ-flavored chips made from cricket flour are delicious for me!
The cricket is a hemimetabolous insect, with adults sized ~3cm. We easily feed them with goldfish chow and collect eggs using wet towel paper, and they hatch approximately two weeks after egg laying. Nymphs and adults are kept in the insect cages with folded paper as shelters (Figure 2). The growth rate depends on temperature: in standard breeding conditions (28 degrees Celsius), first instar nymphs hatch within two weeks, and nymphs grow up to adult by one month. All developmental stages are suitable for observation of detailed morphology, behaviors and gene expression during development.
Since crickets are relatively big and in comparison with other insects have strong vital force (which is suitable for dissection to detect action potential from targeted neurons), they have been used as a model insect for physiological research. Neuronal activities are measured by electrophysiological methods combined with pharmacological approaches.
In 1998, double-stranded RNA (dsRNA)-mediated interference (RNAi) was reported as a useful method for gene silencing by C. elegans researchers. RNAi is widely applicable for other organisms, including insects. Professor Noji’s group in Japan reported that RNAi is a powerful tool to analyze gene function during cricket embryonic development. His group also established transposon-based transgenics and genome editing techniques using zinc finger nucleases, TALE nucleases (TALEN) and CRISPR/Cas system (Nakamura et al., 2010; Watanabe et al., 2012), and analyzed whole genome sequence using next generation sequencing in Gryllus. These new methods established for the cricket open new avenues to understanding genetic basis of embryogenesis, regeneration, circadian rhythm, neurobiology, oogenesis, and so on.
Recently, we reported that epigenetic modifiers regulate leg regeneration (Development), embryonic development (Biology Open) and circadian clock (Zoological letters) by modulating specific gene expression via methylation on 27th lysine residue of histone H3. In this post, I introduce the biological features and benefits of the cricket, especially its use as a model animal in several scientific fields, e.g., regenerative and developmental biology, and chronobiology.
The cricket is a model insect of regeneration biology
Arthropods have a remarkable regenerative ability. The first scientific paper describing tissue regeneration was on the limb regeneration process of crayfish, written by René Réaumur in 1712. Hemimetabolous insects such as dragonflies, stinkbugs, grasshoppers and cockroaches were used for regeneration studies in 19th and 20th centuries. During the 1970s, the group of Prof. Vernon French documented limb regeneration and intercalary regeneration of the cockroach leg, and proposed a theory about positional values of organs; however, no molecular analyses were carried out on the cockroach. Among insect species, methods to analyze the molecular basis of organogenesis were only established in Drosophila melanogaster until the 1990s. However, the regenerative ability of Drosophila is quite limited: it can regenerate only imaginal discs but not adult legs or wings, and the body size of Drosophila is too small to be manipulated compared with crickets. Gryllus bimaculatus has remarkable regenerative ability (similarly to the cockroach) and methods to analyze molecular basis of regeneration is established, hence we start to analyze molecular basis of leg regeneration using the cricket.
To study the molecular basis of leg regeneration, we perform RNAi in the third instar nymph and then amputate the metathoracic leg (T3) of the nymph. The crickets grow up to third instar nymphs one week after hatching at 28 degrees. Just after molting from second to third instar, the cuticle of the third instar nymphs is whitish and soft, then turns brownish and blackish, and hard by a few hours (Figure 1, window). The nymphs are around 5 mm, and look like ants. We collect whitish nymphs into 9 cm petri dish and chill on ice to anesthetize by 30 minutes. To perform RNAi in cricket nymphs, we inject dsRNA into anesthetized cricket nymphs by grass capillary held by a mechanical injector. Injecting is very easy: just insert the capillary into the abdomen of the cricket nymph and push the bottom on the mechanical injector three times (Figure 3). Then we can inject approximately 200 nL of dsRNA solution into the cricket nymph. Two days after performing RNAi, expression of endogenous target gene was decreased.
Each of the cricket prothoracic (T1), mesothoracic (T2) and T3 legs consist of the coxa, trochanter, femur, tibia, tarsus and claws, while the tarsus consists of tarsomere 1 (Ta1), tarsomere 2 (Ta2), and tarsomere 3 (Ta3). To observe regeneration process of the cricket leg, we amputate the legs using ophthalmic scissors. The lost part of the leg is completely restored by the sixth instar (within 2-3 weeks post amputation, Figure 3), and the shape of regenerated legs is indistinguishable from that of intact contralateral legs (Nakamura et al., 2008). Transplantation experiments such as intercalation, reversed intercalation, and tripod formation can be performed on cricket legs (host: T3 leg; graft: T2 leg). In the “tripod experiment”, supernumerary legs formation is induced between the host stump and graft piece (Nakamura et al., 2007) (Figure 3). To transplant a graft piece of T2 leg into the host stump of T3 leg, we just cut T2 leg at the tibia and put the T2 graft piece on the T3 host stump aligning the dorsoventral and anteroposterior axes between host and graft. Since diameters of T3 host stump is bigger than that of T2 graft piece, T2 leg easily fit to T3 leg. There is even no need to use glue to connect the host and graft. After transplantation, the cricket is put on ice 30~60 minutes to anesthetize again, avoiding dislocation of the graft piece from the host stump. The transplanted cricket will molt to next instar nymph after three days, and then newly-formed cuticle continuously covers the host stump and graft piece.
The expression pattern of target genes in the regenerating leg is detectable in the regenerating leg at two, five, or six days post amputation (dpa) by in situ hybridization. Since the cuticles of the insect inhibit penetration of RNA probes, we remove the cuticles from fixed cricket legs. The cuticles of regenerating legs at 2, 5, or 6 dpa are removed using tweezers and micro-scissors under the stereo microscope. The cuticle-removed regenerating legs can be processed for immunostaining using anti-methylated histone H3 lysine 27 (H3K27) (Hamada et al., 2015), anti-5-ethynyl-2′-deoxyuridine (EdU) (Bando et al., 2009), and anti-phosphorylated histone H3 serine 10, and so on (Figure 4).
There are some morphological differences between the T2 and the T3 legs: the T3 leg has three pairs of tibial spur at the distal tibia and one pair of tarsal spur at the distal Ta1. In contrast, the T2 leg has two pairs of tibial spur at the distal tibia and has no tarsal spur at the distal Ta1. Thus, we can select the T3 leg from the T2 leg depending on the purpose of studies. For instance, when I performed RNAi against E(z) and amputation experiments at distal T3 tibia in the third instar nymph, the regenerated leg has an ectopic leg segment between the tibia and tarsus. However, I was not able to determine whether the ectopic segment is tibia or Ta1. If the ectopic segment of T2 regenerated leg has spurs at the end of the segment, we can predict that the segment has a tibia identity. In control crickets, a T2 leg was amputated at a proximal tibia in the third instar nymph, the regenerated leg restored the lost part of tissues including tibia, tibial supers, tarsus and claws. As I expected, in E(z)(RNAi) crickets, the regenerated T2 leg restored the lost part of tissue and the ectopic segment had spurs. These results suggest that the ectopic segment has a tibia identity and morphological character of T2 leg is suitable for regeneration research (Hamada et al., 2015) (Figure 5).
The cricket shows short germ segmentation
A large number of the cricket eggs are collectable at all times of the year. We supply water to the crickets using wet towel paper put into petri dishes. Adult female crickets lay eggs into the wet towel paper. An adult female cricket lays ~3,000 eggs in a few weeks. Usually we keep ~100 adult males and females, hence we can get ~100 eggs within one hour. If we quickly move the towel paper in the water, the laid eggs are removed from the paper, then we can collect the cricket eggs from the water using a tea strainer. We can inject dsRNA for embryonic RNAi, transposons for transgenesis, RNA encoding TALEN or CRISPR/Cas for genome editing, pharmacological reagents and others into the ventroposterior side of the fertilized eggs (Figure 6).
The egg diameter is approximately 3 mm x 0.5 mm. The developmental stage of cricket embryos is well documented from fertilization to hatching. The early embryo is formed on the ventroposterior side of the egg. We dissect fertilized eggs to get cricket embryos in the desired developmental stages (Donoughe and Extavour, 2016). These embryos are applicable for whole mount in situ hybridization to detect gene expression. While the regeneration processes of the leg can be observed during molting, the embryo can be continuously observed and collected anytime for molecular analysis. For efficient RNAi during early embryonic development, we inject dsRNA into the body fluid of adult female (Matsuoka et al., 2015). Early embryos in the ovary intake dsRNA, called parental RNAi. The model animal Drosophila melanogaster shows the long germ segmentation in its developmental mode, while the cricket shows the short germ segmentation in its developmental mode. Thus, in Evo-Devo studies, the cricket is expected to be a more representative model insect.
The cricket shows characteristic circadian rhythm
The cricket shows diurnal activity in nymphal stages, but after imaginal molting, the activity phase is shifted to be nocturnal (Tomioka and Chiba, 1982). Activities of cricket individuals are documented by using small cricket cages with seesaw-like flower, that is a magnetic sensor switch on the flower (Figure 7).
To record the actgram of more smaller animals (the body length of G. bimaculatus is less than 25mm), I have succeeded to utilize LAM25 with an infrared laser sensor for Modicogryllus siamensis (Trikinetics inc). The data from the seesaw and infrared laser locomotor recorder can be analyzed by Actogram J, a plug-in in Image J (unpublished). Control and/or RNAi-treated crickets are reared in the small cage to measure continuous actogram for more than a month. Previously, it was reported that the central clock system is located in the optic lobe (Tomioka and Chiba, 1992), where clock component gene expression is oscillated daily (Tomioka, 2014). In the cricket, the optic lobes locate bilaterally in the brain, at the inside of compound eyes. The clock genes and their expression profiles are similar to those of model animals including mice and flies. For instance, the clock component genes of mice, such as Clock, Bmal1, period1/2/3, Timeless and Cryptochrome are conserved in fly and cricket as Clock (Clk), cycle (cyc), period (per), timeless (tim) and cryptochrome2 (cry2), respectively, and the transcriptional/translational molecular feedback loops for period of 24 hours are consistent between mice and insects. When we measure the activities of per(RNAi) crickets, they show arrhythmic actogram. Furthermore, it is easy to obtain the whole brain, which is also a central clock component, and used for in situ hybridization (Hamada et al., 2009), immunostaining, and so on. Recently, we address the big challenges that elucidate the relationship between leg regeneration and photoperiodism, and systematize knock-in cricket lines using CRISPR/Cas9 system.
The cricket is one of the most familiar insects to enjoy insect songs in autumn in Japan. A Japanese author, Sei Shonagon mentioned that “Evening is the best time in the day in autumn, because we can enjoy the flight of wild geese and songs of insects” in her essay named “Makurano soushi (The Pillow Book)”, written in the 10th century. Now the cricket is used as a model insect of regeneration biology and RNAi-based analyses show new insights into tissue regeneration in addition to his song. I am thinking about new experiments listening to crickets’ chirp songs today.
References
Bando, T., Mito, T., Maeda, Y., Nakamura, T., Ito, F., Watanabe, T., Ohuchi, H. and Noji, S. (2009). Regulation of leg size and shape by the Dachsous/Fat signalling pathway during regeneration. Development136, 2235-2245.
Donoughe, S. and Extavour, C. G. (2015). Embryonic development of the cricket Gryllus bimaculatus. Dev Biol411, 140-156.
Hamada, A., Miyawaki, K., Honda-sumi, E., Tomioka, K., Mito, T., Ohuchi, H. and Noji, S. (2009). Loss-of-function analyses of the fragile X-related and dopamine receptor genes by RNA interference in the cricket Gryllus bimaculatus. Dev Dyn238, 2025-2033.
Hamada, Y., Bando, T., Nakamura, T., Ishimaru, Y., Mito, T., Noji, S., Tomioka, K. and Ohuchi, H. (2015). Leg regeneration is epigenetically regulated by histone H3K27 methylation in the cricket Gryllus bimaculatus. Development142, 2916-2927.
Matsuoka, Y., Bando, T., Watanabe, T., Ishimaru, Y., Noji, S., Popadic, A. and Mito, T. (2015). Short germ insects utilize both the ancestral and derived mode of Polycomb group-mediated epigenetic silencing of Hox genes. Biol Open4, 702-709.
Nakamura, T., Mito, T., Bando, T., Ohuchi, H. and Noji, S. (2008). Dissecting insect leg regeneration through RNA interference. Cell Mol Life Sci65, 64-72.
Nakamura, T., Mito, T., Tanaka, Y., Bando, T., Ohuchi, H. and Noji, S. (2007). Involvement of canonical Wnt/Wingless signaling in the determination of the positional values within the leg segment of the cricket Gryllus bimaculatus. Dev Growth Differ49, 79-88.
Nakamura, T., Yoshizaki, M., Ogawa, S., Okamoto, H., Shinmyo, Y., Bando, T., Ohuchi, H., Noji, S. and Mito, T. (2010). Imaging of transgenic cricket embryos reveals cell movements consistent with a syncytial patterning mechanism. Curr Biol20, 1641-1647.
Tomioka, K. (2014). Chronobiology of crickets: a review. Zoolog Sci31, 624-632.
Tomioka, K. and Chiba, Y. (1982). Post-embryonic development of circadian rhythm in the cricket, Gryllus bimaculatus: A rhythm reversal. J Comp Physiol147, 299-304.
Tomioka, K. and Chiba, Y. (1992). Characterization of an optic lobe circadian pacemaker by in situ and in vitro recording of neural activity in the cricket, Gryllus bimaculatus. J Comp Physiol A171, 1-7.
Watanabe, T., Ochiai, H., Sakuma, T., Horch, H. W., Hamaguchi, N., Nakamura, T., Bando, T., Ohuchi, H., Yamamoto, T., Noji, S., et al. (2012). Non-transgenic genome modifications in a hemimetabolous insect using zinc-finger and TAL effector nucleases. Nat Commun3, 1017.
This post is part of a series on a day in the life of developmental biology labs working on different model organisms. You can read the introduction to the series here and read other posts in this series here.
This post was originally published on the DMDD blog.
The Zika virus has raised global awareness of birth defects more than at any time in the last 50 years [1]. A recent Nature Editorial explores the opportunities this presents to increase support for vaccination programmes, compulsory fortification of food staples and investment in population-scale databases to mine information on the causes of birth defects. But what does it mean for research into the genetic basis of developmental disorders and rare diseases?
HOW RARE IS RARE?
Some birth defects, such as Zika-linked microcephaly, are infectious in origin. But many are related to genetic mutations, which can also prevent an embryo from developing as it should. The rare diseases resulting from genetic mutations can be chronic and life threatening, and 30% of rare disease patients die before their fifth birthday [2]. Many mutations can also lead to miscarriage. Genetic cardiac conditions affect around 1% of newborn babies [3], but it’s estimated that these defects prevent around ten times as many foetuses from surviving until birth.
A rare disease is difficult to study, because of the relatively small group of patients – by definition less than 1 in 2000 of the general population are affected. But with more than 6000 known rare diseases, 80% of these with a genetic component, it’s likely you or someone you know has a rare disease. 7% of the population will be affected at some point in their lives, which equates to around 3.5 million people in the UK alone [3].
CLUES FROM SYSTEMATIC SCREENS
We have a huge challenge to find and prevent the causes of rare diseases. For those diseases linked to genetic mutations it’s vital that we understand their genetic basis, and basic research in developmental biology is fundamental to this.
Mouse research in particular offers a wealth of information, thanks to systematic studies that would not be possible in human patients. The mouse genome can be manipulated to delete (knock out) a specific gene. The resulting embryos and adult mice can then be studied to look for abnormalities in the way they develop. It’s a unique way to understand the role that a specific gene plays in development from embryo through to adult, and the developmental abnormalities that may arise from a fault with the gene. Around 30% of gene deletions cause abnormalities so severe that the embryo does not survive until birth, so study of these genes also provides clues about the genetic basis of miscarriage.
Systematic screens by the DMDD and IMPC are working to study the effects of individually knocking out every gene in the mouse genome. With all data freely available online, they are a rich resource for those researching human birth defects, miscarriage and developmental disorders.
The Zika virus has put birth defects back on the political agenda, and scientists have a challenge to find and prevent their causes. In this context, systematic screens of mouse gene knockouts and other gene mutations are more important than ever.
DMDD provides a free, online database of embryonic-lethal mouse gene knockouts: see dmdd.org.uk.
 (No Ratings Yet)
(No Ratings Yet)








 (23 votes)
(23 votes)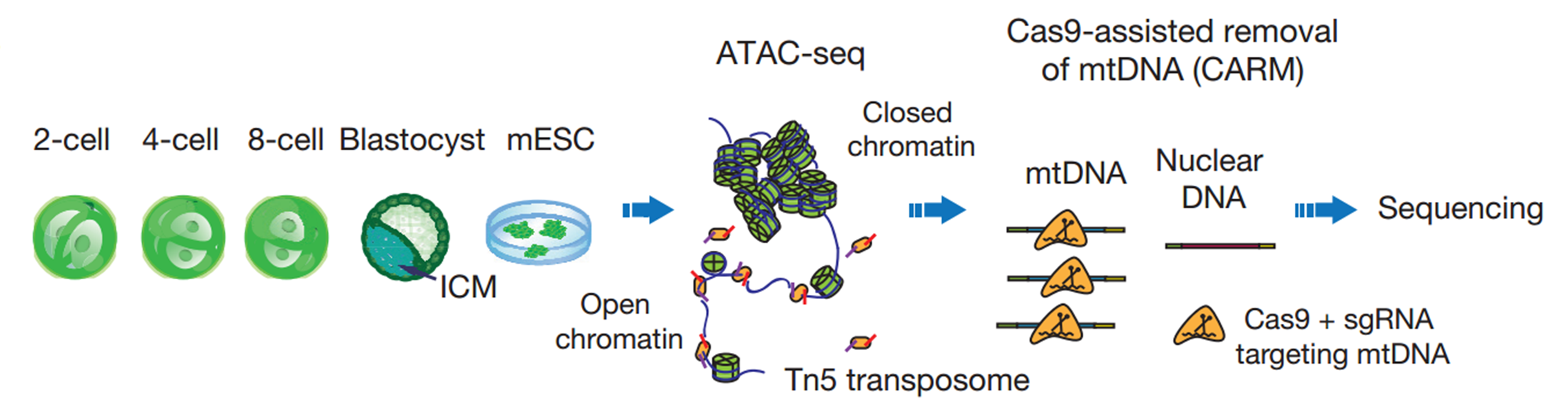




 Enrico Coen CBE FRS is a Project Leader at the John Innes Centre in Norwich, UK, who uses a variety of approaches to study patterning and morphogenesis in plants. We met with Enrico at the Spring Meeting of the British Society for Developmental Biology, where he was awarded the Waddington Medal, to ask him more about his career and his passion for art and book-writing. See the Spotlight article on p.
Enrico Coen CBE FRS is a Project Leader at the John Innes Centre in Norwich, UK, who uses a variety of approaches to study patterning and morphogenesis in plants. We met with Enrico at the Spring Meeting of the British Society for Developmental Biology, where he was awarded the Waddington Medal, to ask him more about his career and his passion for art and book-writing. See the Spotlight article on p.  Cells can signal by releasing exosomes – extracellular vesicles containing bioactive molecules such as RNA, DNA and enzymes. Recent work has suggested that exosomes can also carry signalling proteins, including ligands of the Notch receptor and secreted proteins of the Hedgehog and WNT families. Here,
Cells can signal by releasing exosomes – extracellular vesicles containing bioactive molecules such as RNA, DNA and enzymes. Recent work has suggested that exosomes can also carry signalling proteins, including ligands of the Notch receptor and secreted proteins of the Hedgehog and WNT families. Here, 


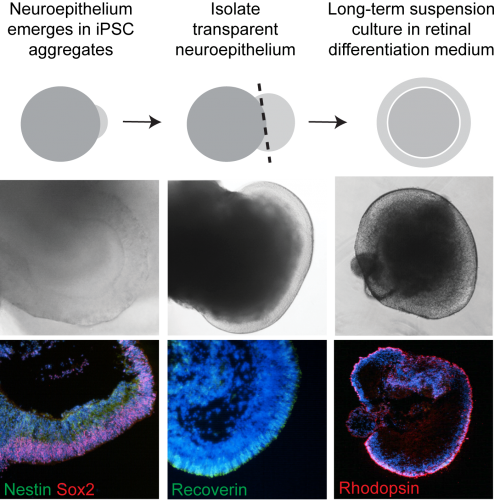
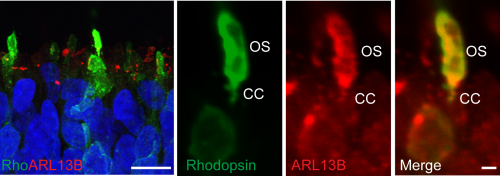
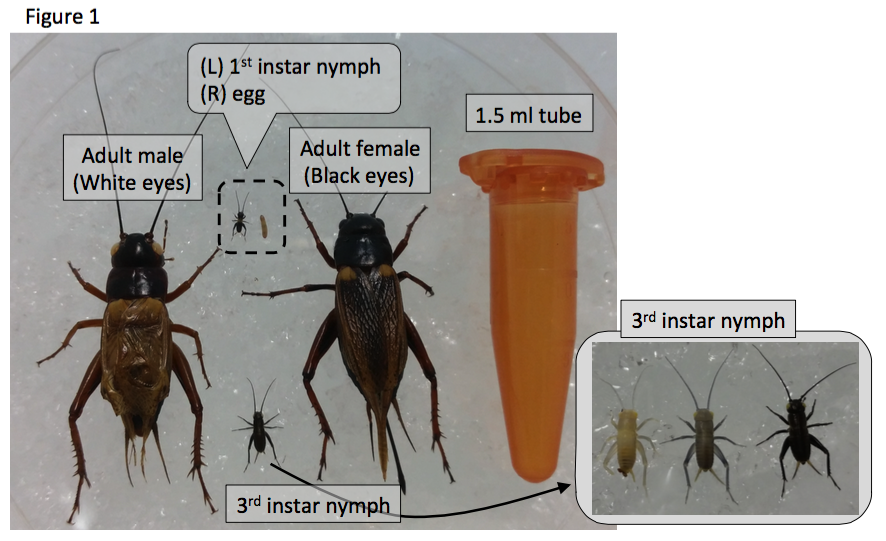

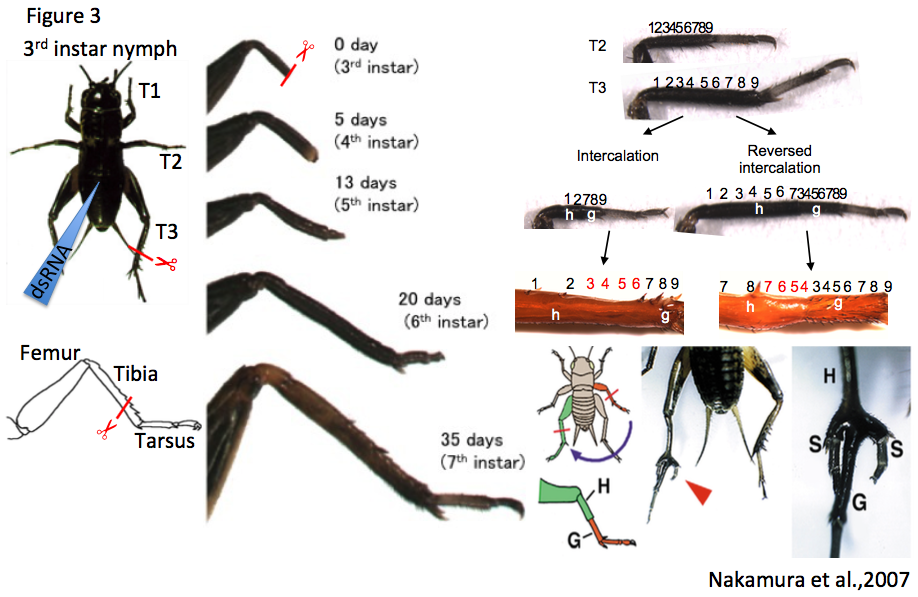
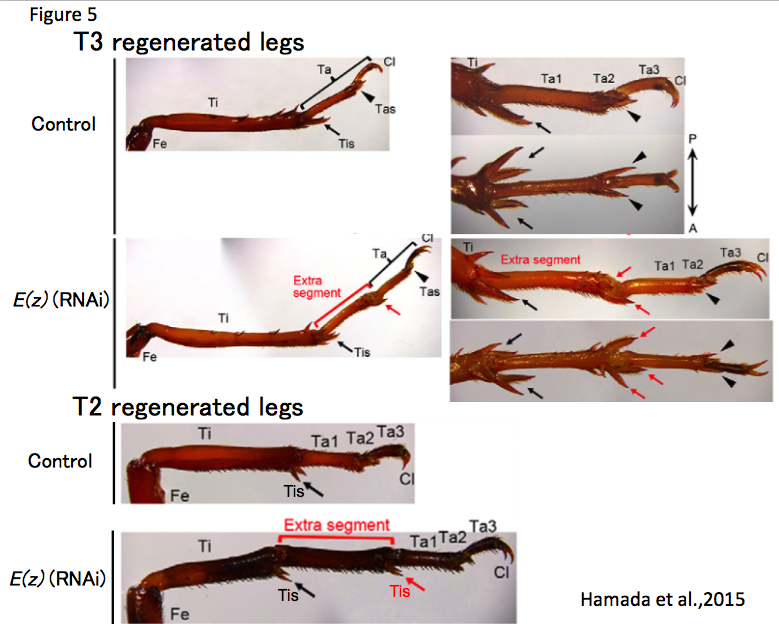
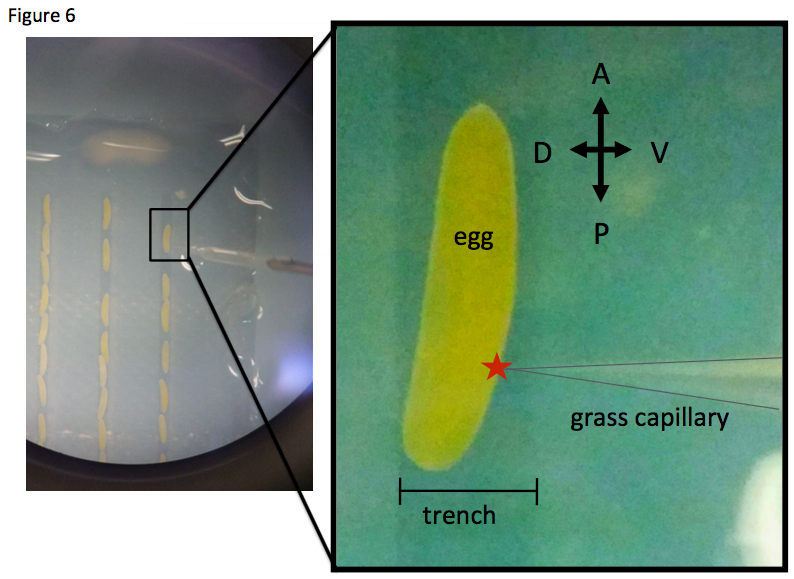
 day in the life of developmental biology labs working on different model organisms. You can read the introduction to the series
day in the life of developmental biology labs working on different model organisms. You can read the introduction to the series 