Disease Models & Mechanisms invites you to submit original research for consideration for an upcoming Special Collection ‘Neurodegeneration: From Models to Mechanisms to Therapies‘. This ongoing collection will focus on mechanistic insights into neurodegenerative diseases using model systems, with an emphasis on key translational advances made in recent years. This collection will be launched with a dedicated issue that will benefit from high visibility to basic researchers through to clinicians interested in understanding and treating neurodegenerative diseases.
High visibility and impact (2014 Impact Factor 5)
Open Access (CC-BY licence) and PMC deposition
Rapid peer review and publication
Indexed in Medline, ISI and Scopus
Not-for-profit publisher
Here are the highlights from the current issue of Development:
A new paradigm for Wnt/β-catenin signalling
The Wnt/β-catenin signalling pathway is a key pathway involved in a myriad of developmental processes, from body axis patterning to cell migration and fate specification. The role of the Wnt pathway is to regulate transcription of specific target genes, and it has long been thought that the driving event for this is the recruitment of β-catenin to specific gene loci on the chromatin. Now, however, on p. 1914, Stefan Hoppler and colleagues provide in vivo evidence that, rather than β-catenin recruitment, it is instead the context-specific events that occur subsequent to β-catenin binding that enable gene-specific regulation. The authors use ChIP-seq to show that β-catenin is recruited to many genomic loci in Xenopus early embryonic development, while RNA-seq reveals that many of these β-catenin-bound loci are not transcriptionally regulated by Wnt signalling in this context. Instead, the transcriptional response depends on the presence or absence of additional mechanisms, for example FGF and BMP signalling. This important study advances the current understanding and interpretation of how Wnt/β-catenin signalling operates to regulate gene-specific transcription in different developmental contexts.
Oocyte maturation: cAMP and PKA called into question
The production of a mature vertebrate egg is a lengthy process in which the developing oocyte undergoes meiotic arrest followed by a long incubation period, before finally resuming meiosis in preparation for ovulation. The prevailing dogma in the field has been that, in Xenopus, meiotic arrest is released through a drop in cyclic adenosine monophosphate (cAMP) levels and protein kinase A (PKA) activity, which occurs following exposure to progesterone. In this issue (p. 1926), Khaled Machaca and colleagues provide evidence that challenges this dogma, as they demonstrate that no change is detectable in cAMP levels and PKA activity as meiotic arrest is released in the Xenopus oocyte. The authors use in vivo reporters to detect cAMP and PKA levels in real time in single cells, and show that there is no correlation between the rate of meiotic resumption and levels of cAMP or PKA inhibition. Furthermore, the authors develop conditions in which meiotic release is indeed possible in the presence of high levels of cAMP. These surprising results provide a new model for the release of meiotic arrest in the Xenopus oocyte and suggest that this phenomenon occurs through a positive signal downstream of the progesterone receptor that overcomes cAMP/PKA inhibition of meiosis resumption.
Uncovering the mechanisms of Müller glia activation
Radial glia cells in the central nervous system serve as an important source of progenitor cells for generating a range of neural cell types. Retinal regeneration in vertebrates relies on a specialised type of radial glia, called Müller glia, which are normally quiescent but can be stimulated to undergo proliferation and differentiation in order to generate new neurons. In this issue (p. 1859 and p. 1874), two studies shed light on the molecular mechanisms that regulate Müller glia activation and proliferation in response to injury and in different vertebrate species.
In the first study, Andy Fischer and colleagues investigate the role of mTor signalling in the formation of Müller glia-derived progenitor cells (MGPCs) in the chick retina. The authors use NMDA to induce a cytotoxic response, and observe that mTor signalling is transiently activated upon activation of Müller glia cells. Inhibition of mTor signalling in vivo prevents the proliferation of the Müller glia cells and blocks the regenerative response. Using a range of inhibitors and readouts, the authors show that mTor signalling is required for the proliferation of MGPCs. The authors further show that mTor signalling is activated in response to insulin, IGF1 and FGF2, and that this response is most likely independent of the MAPK pathway.
In the second study, Joachim Wittbrodt and colleagues look at the role of a single factor, Atoh7, in directing Müller glia cells to proliferate and differentiate in the absence of an injury. The authors use a fluorescent transcriptional reporter of atoh7 to demonstrate atoh7 expression in proliferating Müller glia cells after retinal injury in medaka. The authors then use an inducible system to activate expression of atoh7 in vivo in the Müller glia cells, and find that this is sufficient to drive the cells to re-enter the cell cycle and undergo proliferation. Forced expression of atoh7 in these cells activates Notch signalling, and indeed the authors show that overexpression of the Notch intracellular domain can recapitulate the effects seen by atoh7 overexpression. Importantly, not only did atoh7 overexpression in Müller glia lead to cell cycle re-entry and proliferation, but the authors also observed the formation of neurogenic clusters and subsequent de novo neurogenesis following atoh7 overexpression in these cells. Together, these two studies bring together novel and exciting findings regarding the regulation of Müller glia proliferation following injury and the subsequent regenerative response.
PLUS…
Cell fate control by pioneer transcription factors
Recent studies have shown that pioneer factors are crucial for cellular reprogramming and that they are implicated in the marked changes in gene regulatory networks that occur in various cancers. Here, Makiko Iwafuchi-Doi and Kenneth Zaret provide an overview of the contexts in which pioneer factors function, how they can target silent genes, and their limitations at regions of heterochromatin. See the Development at a Glance poster article on p. 1833
Circular RNAs: analysis, expression and potential functions
In early 2012, circular RNA (circRNA) was shown to be a transcriptional product in thousands of human and mouse genes and in hundreds of cases constituted the dominant RNA isoform. Subsequent studies revealed that the expression of circRNAs is developmentally regulated, tissue and cell-type specific, and shared across the eukaryotic tree of life, suggesting important functions for these molecules. Here, Steven Barrett and Julia Salzman describe major advances in the field of circRNA biology, focusing on the regulation of and functional roles played by these molecules. See the Primer on p. 1838
The regulation and plasticity of root hair patterning and morphogenesis
Root hairs are highly specialized cells found in the epidermis of plant roots that play a key role in providing the plant with water and mineral nutrients. Many studies have shown that the fate of root epidermal cells, which differentiate into either root hair or non-hair cells, is determined by a complex interplay of intrinsic and extrinsic cues. Here, Wolfgang Schmidt and colleagues review these studies and discuss recent evidence suggesting that environmental information can be integrated at multiple points in the root hair morphogenetic pathway and affects multifaceted processes at the chromatin, transcriptional and post-transcriptional levels. See the Review on p. 1848
Earlier this month, two papers were published (from the Brivanlou and Zernicka-Goetz labs) that reported in vitro systems to study development of the human embryo through implantation stages. These experiments have kept human embryos developing for longer than any previous work, and close in on the 14-day limit imposed by many governmental and regulatory bodies. 14 days were set as a limit (at a time when it was technically impossible to keep embryos alive this long) because this is the stage at which the primitive streak emerges – the time at which embryos can no longer split or fuse – and has been considered by some to be the stage at which a ‘morally significant individual’ comes into being (see this discussion in Nature). But now that we can culture human embryos up to this point, is this 14-day rule still appropriate? Would it be ethically ‘wrong’ to try and study human gastrulation in culture, or do the potential advances in our understanding of human development (along with their possible therapeutic implications) outweigh any ethical concerns involved? This month we are asking:
Should the 14 day limit on human embryo culture remain in place, or should it be extended or even dropped?
Share your thoughts by leaving a comment below! You can comment anonymously if you prefer. We are also collating answers on social media via this Storify. And if you have any ideas for future questions please drop us an email!
Mammalian development and tissue homeostasis are complex and dynamic biological processes. Focusing on Wnt signal transduction, our research aims to understand the genetic and epigenetic mechanisms that control gene activation in the mammary gland. We are now recruiting a postdoctoral fellow that fits one of the following profiles:
You are a ‘dry-lab’ candidate with extensive and proven expertise in complex bioinformatics analyses and an interest in analyzing large datasets, who can complement our wet-lab skills.
You are a ‘wet-lab’ candidate with proven expertise in embryonic stem cell targeting and the generation of genetically engineered mouse models.
You will be part of a young and growing research team that uses a combination of in silico, in vitro and in vivo approaches to under- stand normal (stem) cell behavior, with the ultimate goal of trans-lating principles from developmental and stem cell biology to regenerative medicine and breast cancer research. We are embedded within the section of Molecular Cytology at the Swammerdam Institute for Life Sciences and collaborate with other groups in the institute, but also with other labs in the Netherlands (e.g. at the Netherlands Cancer Institute) and abroad. We have strong ties to the field of mammary gland biology (active member of the ENBDC), as well as to the Wnt-signaling community.
Details of the job description (including tasks and requirements) and the appointment are available at the University website, which is also where you can submit you application.
In September, Development will be holding the second of our ‘From stem cells to human development‘ meetings – following on from the hugely successful first event in Autumn 2014 (see here and here). This year’s meeting promises to be equally exciting, with a fantastic line-up of speakers (check out the preliminary program) and a great venue just outside Boston, USA.
This should be a great event for anyone interested in how we can apply stem cell technologies to understand the development of our own species, spanning a broad range of topics from pluripotency and early human embryo patterning through to applying the lessons we’ve learned from development for therapeutic purposes.
Registration closes at the end of next month, but with a limited number of spaces available, we’d encourage anyone interested to apply as soon as possible to secure your place!
We look forward to seeing you at Southbridge in September!
A major challenge in cell and developmental biology is to understand the mechanisms whereby cells interact with each other to form the variety of complex tissue forms present in organisms. This requires visualizing and analysing different cellular processes across multiple scale levels -from the subcellular to the tissue, i.e. generating cell and tissue models at different levels of complexity. The different cells as well as their essential sub-cellular components can be observed by fluorescence microscopy and reconstructed by means of image processing techniques. Then, three-dimensional (3D) digital representations of tissue can be generated using the structural information extracted from the microscopy images. A set of 3D digital representations of tissue at multiple scale levels constitutes a geometrical model of the tissue. Such a geometrical model provides the means of extracting detailed morphological and structural information of the tissue as well as its spatial variability. Based on such information, new hypotheses about tissue organization can be generated (Figure 1).
In our recent study published in eLife, we developed a software platform for the reconstruction and quantitative analysis of tissue architecture in 3D (Morales-Navarrete et al., 2015). We combined newly developed and established image analysis algorithms in a pipeline that allows for a flexible workflow to reconstruct and analyse different structures forming the tissue with high accuracy. It is implemented as part of the stand-alone freely available software MotionTracking (http://motiontracking.mpi-cbg.de; the name is historical as it was first developed for the tracking of endosomes in cultured cells (Rink et al., 2005)) and addresses several unmet computational needs, such as 1) high accuracy of 3D geometric reconstruction, 2) image processing of large volumes of tissue, 3) fast image analysis amenable to middle-throughput, 4) it can be run on a regular PC, 5) it is adaptable to various tissues and imaging conditions, 6) it provides a flexible tool for tissue reconstruction without need of further programming or scripting and 7) it allows quantitative morphometric and spatial analysis at cellular and tissue level.
As proof of principle, the pipeline was applied to the analysis of liver tissue of adult mice. The resulting geometrical model provided a complete description of different morphological parameters across multiple scales from tissue to subcellular level (Figure 1A and http://dx.doi.org/10.7554/eLife.11214.003, http://dx.doi.org/10.7554/eLife.11214.012, http://dx.doi.org/10.7554/eLife.11214.022) and uncovered a significant amount of new biological information. For example, we could map the distribution of different populations of hepatocytes with different number of nuclei and DNA content within the mouse liver lobule. It has been reported that hepatocytes are remarkably heterogeneous in terms of ploidy but whether they are randomly distributed or follow a specific spatial organization remains controversial (Gentric and Desdouets, 2014). Our quantitative and spatial analysis of the geometrical model revealed an unexpected spatial distribution of hepatocytes with distinct DNA content. Whereas hepatocytes with low ploidy were enriched in the peri-central and peri-portal regions, high ploidy hepatocytes were preferentially located in the middle region (Figure 1B and http://dx.doi.org/10.7554/eLife.11214.027). This is the first time that polyploidy is found to be zonated and follow a specific pattern.
Such an unexpected zonation suggests the existence of mechanisms underlying the functional specialization of hepatocytes during tissue homeostasis. One of the most remarkable features of liver tissue is the zonation of metabolic functions (hepatocytes located in the vicinity of the portal vein have different metabolic activities than hepatocytes located near the central vein). Our findings therefore suggest a possible role of polyploidy in liver metabolic functions (Figure 1C). Moreover, low ploidy hepatocytes (enriched in the peri-central and peri-portal regions) may correspond to the cells recently implicated in hepatocyte renewal (Wang et al., 2015, Font-Burgada et al., 2015). Altogether, these results show how tissue geometrical models can be used to unravel principles of tissue architecture but also generate hypotheses about tissue structure and function.
We expect our pipeline to be very useful to the biology community since it can be used for several applications from basic (understanding of how cells form tissues) to clinical research (diagnosis of diseases by 3D digital histology).
Figure 1: Using tissue geometrical models to derive principles of tissue organization and function. A) Geometrical model of liver tissue generated from confocal microscopy images. The main components of liver tissue architecture are central vein (cyan), portal vein (orange), bile canaliculi (green), sinusoids (magenta) and hepatocytes (random colours). B) Measurements of hepatocytes volume, number of nuclei per cell and DNA content were extracted, and the spatial analysis of the localization of hepatocytes with different ploidy (estimated from the volume, number of nuclei and DNA content per cell) revealed zonation patterns within the lobule. C) These zonation patterns showed a correlation with the metabolic zones in liver, which lead to the generation of the hypothesis of the existence of a possible correlation between cell polyploidy and liver metabolic functions.
Software resources:
All resources for our platform, including user manuals, examples and the latest version of the software, can be found on http://motiontracking.mpi-cbg.de/get/.
FONT-BURGADA, J., SHALAPOUR, S., RAMASWAMY, S., HSUEH, B., ROSSELL, D., UMEMURA, A., TANIGUCHI, K., NAKAGAWA, H., VALASEK, M. A., YE, L., KOPP, J. L., SANDER, M., CARTER, H., DEISSEROTH, K., VERMA, I. M. & KARIN, M. 2015. Hybrid Periportal Hepatocytes Regenerate the Injured Liver without Giving Rise to Cancer. Cell, 162, 766-79.
GENTRIC, G. & DESDOUETS, C. 2014. Polyploidization in liver tissue. Am J Pathol, 184, 322-31.
RINK, J., GHIGO, E., KALAIDZIDIS, Y. & ZERIAL, M. 2005. Rab conversion as a mechanism of progression from early to late endosomes. Cell, 122, 735-49.
WANG, B., ZHAO, L., FISH, M., LOGAN, C. Y. & NUSSE, R. 2015. Self-renewing diploid Axin2(+) cells fuel homeostatic renewal of the liver. Nature, 524, 180-5.
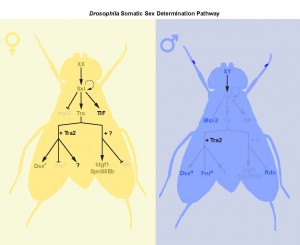
Male and female fruit flies differ in many aspects of development and physiology. For example, males and females differ in abdominal pigmentation, sex comb formation, courtship behaviours, the development and wiring of the central nervous system, internal reproductive organs, and genital morphology. Many elegant studies on the genes that regulate sex differences in these traits have led to the prevailing model of sex determination in Drosophila where the number of sex chromosomes determines male or female sexual identity1,2.
Briefly, the presence of two X chromosomes in female cells causes the production of an X-derived protein called Sex-lethal3. Sxl is a splicing factor, and introduces a splice into the pre-mRNA of an autosomal gene called transformer (tra) that allows a functional protein to be produced4-6. Like Sxl, Tra is a splicing factor, and together with its co-factor transformer2 (tra2), binds to the pre-mRNA of two genes: doublesex (dsx) and fruitless (fru)7-10. Tra-dependent splicing of dsx pre-mRNA causes the production of an mRNA encoding the female-specific isoform of Dsx protein (DsxF). In contrast, Tra binding to fru pre-mRNA introduces a stop codon into transcripts derived from the P1 promoter, and no Fru P1 proteins are produced in females. In males, with one X chromosome, Sxl is not induced, and consequently no functional Tra protein is produced. Lack of Tra means that dsx and fru P1 pre-mRNA undergo default splicing, and the male-specific isoforms of each protein are made (DsxM and FruM).
This sex determination cascade explains most aspects of sexual development and reproduction. However, male and female flies also differ in body size, stress responses, immunity, physiology, and lifespan11-14. Yet the mechanisms underlying sex differences in these additional traits remain unclear. The focus of this blog post is to highlight the findings from our two recent papers on sex differences in Drosophila body size (Rideout et al. 201515) and adult intestinal stem cell homeostasis (Hudry et al. 201616). We will review how these findings advance our understanding of the genetic control of sexual identity in flies, and the use of Drosophila as a model to study the molecular mechanisms underlying sexual dimorphism in body size and gut physiology.
Figure 1 – Drosophila Somatic Sex Determination Pathway. In flies, the number of sex chromosomes (X) determines sex. XX animals develop as females, whereas XY animals develop as males. Two X chromosomes causes the production of an X-derived protein called Sex-lethal (Sxl). Sxl introduces a splice into transformer (tra) pre-mRNA that allows a functional protein to be produced. Many aspects of sexual identity are controlled by Tra and its co-factor transformer2 (tra2) via regulation of doublesex (dsx) and fruitless (fru) pre-mRNA. However, our two recent papers, Rideout et al. (2015) and Hudry et al. (2016), identify at least two additional branches downstream of tra in the regulation of sexual identity: one that is tra2-dependent but dsx-/fru-independent, and another that is independent of tra2 and dsx/fru. These findings reveal additional complexity in the Drosophila sex determination pathway.
Sex determination: different tissues, different rules
Studies on courtship behaviour, abdominal pigmentation, and sex comb formation show a clear role for the canonical Sxl→tra→dsx/fru splicing cascade in the creation of sex differences3,17. However, our findings, together with data from other studies18,
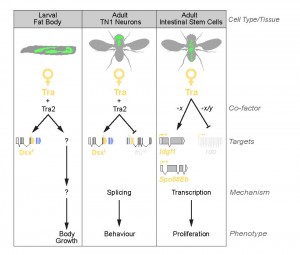
demonstrates that the genetic control of sexual identity involves additional downstream effector pathways (Fig. 1). These pathways operate in different tissues, and at different stages of the life cycle, suggesting that the process of sex determination in flies is more complex, and includes more genes, than the central core of Sxl/tra/dsx/fru (Fig. 2).
Beyond dsx and fru: new downstream targets of tra
Although there is growing evidence to support a tra-independent branch of the sex determination pathway downstream of Sxl, whether an additional branch exists downstream of tra is unclear. Previous studies on dissatisfaction (dsf), a gene with effects on sex determination in males and females, suggested a tra-dependent, dsx-independent branch of the sex determination pathway19. Yet the authors were not able to rule out the possibility that dsf may be target of fru20. Further, there was no evidence supporting sex differences in dsf expression, transcript levels, or splicing. Our two papers now provide concrete evidence of the existence of a tra-dependent, but dsx-/fru-independent, branch of the sex determination pathway in two different contexts: larval body growth, and adult intestinal stem cell (ISC) homeostasis.
In Rideout et al. (2015) we show that tra promotes the increased body size of female flies. Loss of Tra in female larvae decreases body size, whereas ectopic expression of Tra in male larvae increases body size. Specifically, we found that Tra functions in the larval fat body to control overall body size in a non-cell-autonomous manner, and that these effects on body size were reproduced in larvae with fat-specific loss of Tra2. Surprisingly, body size was unaffected in dsx mutants, females ectopically expressing FruM, and in males lacking FruM. Thus tra/tra2, acting independently of fru and dsx, control male-female differences in body size.
Similarly, in Hudry et al. (2016), we show that tra promotes higher ISC proliferation rate in females. This sex difference in adult stem cell divisions controls sex differences in organ size, plasticity during reproduction, and response to tumorigenic insult. Using a series of rescue, and gain- and loss-of-function experiments with dsxF transgenes, dsx/fru mutants, and RNAi transgenes, we ruled out both direct and indirect actions of dsx and fru on ISC proliferation.
Figure 2 – Sex Determination in Drosophila: Different Tissues, Different Rules. The classical sex determination pathway of Sxl/tra/dsx/fru operates in many tissues, including the adult TN1 neurons (middle panel). Interestingly, the classical pathway operates in parallel to the tra-/tra2-dependent, dsx-/fru-independent pathway in the larval fat body. In contrast, in the adult intestinal stem cells, dsx and fru are not present. Thus sexual identity is governed by multiple branches of the sex determination pathway that act in a highly context-dependent manner.
Together, our studies provide strong evidence that Tra’s effects on body size and ISC proliferation are independent of both dsx and fru (Fig. 2). Interestingly, though our findings converge on the existence of a fru-/dsx-independent branch of the sex determination pathway downstream of tra, we found important differences in the molecular mechanisms of tra’s effects on larval growth and ISC proliferation. For example, in larvae, the dsx-/fru-independent effects of tra on body size require tra’s partner tra2. In contrast, tra2 is not required for tra’s effects on ISC proliferation in the adult gut. How tra activity acquires tissue- and cell-type-specificity is not yet understood and emerges as an important new question.
One possibility is that tra acts via co-operation with tissue-specific partners. For example, one study found that among 217 Drosophila genes with Entrez Gene identifiers related to RNA-binding proteins (including splicing regulators), 83 produce multiple transcripts21. The temporal and spatial regulation of the production of these different isoforms is far from being elucidated. However, among these 217 RNA-binding proteins, 35 display very specific expression pattern in adult tissues. Some of these alternative splice variants, or tissue-specific RNA binding proteins, could be context-specific regulators of tra-mediated splicing.
Alternatively, tra might influence ISC proliferation and body size via regulation of other aspects of RNA metabolism. Tra is a member of the SR family of proteins, which play diverse roles in RNA metabolism, including effects on transcript stability, mRNA translation, and transcriptional regulation of gene expression22. A non-splicing function for Tra in ISCs is supported by lack of Tra/Tra2 binding sites near tra-regulated splice junctions in adult ISCs. On the other hand, a genome-wide search successfully identified 18 candidate targets with multiple Tra/Tra2 sites, including tra’s known targets dsx, fru, and Sxl. Indeed, the three functional targets of tra identified in adult ISCs are regulated at the level of mRNA expression rather than splicing, suggesting one explanation for why tra2 is not required for adult ISC proliferation.
Drosophila as a model for sex differences in development and physiology
Sex differences in body size, stress responses and physiology are common to many animals other than flies, including mammals. Decades of research have examined the important contribution that sex hormones make to sex differences in traits such as adiposity and stem cell biology. For example, estrogen in female mammals causes a smaller body size, limits visceral fat accumulation, and increases haematopoietic stem cell self-renewal in females in both homeostatic conditions and during pregnancy23-25.
However, when sex hormones are absent, data from new mouse models shows that chromosomal sex (e.g. XX vs XY) dictates male-female differences in body size and adiposity. For example, in the Four Core Genotypes mouse model (FCG), which allows the effects of sex chromosomes to be directly assessed independently of sex hormones, gonadectomized XX animals achieve a larger body weight than XY animals26. This mirrors the situation in flies, where XX females are larger than XY males. Thus, in the absence of sex hormones, the trends in body size between mammals and flies are remarkably similar.
At present, it remains unclear how sex chromosomes affect body size and adiposity in mice. Flies may therefore provide important insight into the molecular mechanisms underlying these sex hormone-independent effects on body size and adiposity. For example, an important finding from Rideout et al. (2015) is that females achieve a larger body size at least in part due to increased secretion of Drosophila insulin-like peptide 2. Intriguingly, XX FCG mice have higher fasting levels of insulin26. Since the insulin pathway is an important regulator of body weight and adiposity, similar pathways may be involved in mediating the effects of sex on these traits in both flies and mice.
Over the past two decades, Drosophila has provided vital information on the molecular and physiological mechanisms responsible for determining growth during development and final body size31. Drosophila ISCs have more recently emerged as a good model to investigate adult somatic stem cell functions and regulation in vivo32,33. Recent studies using the fly model have identified many conserved intrinsic factors and extrinsic niche signals that drive the proliferation, self-renewal, and differentiation of ISCs34. In addition, a recent study demonstrated that the sex of the gut mediates the sex-biased responses of males and females to lifespan extension by dietary restriction35. We therefore suggest that flies present an excellent model to dissect the molecular mechanisms underlying sex differences in diverse aspects of development and physiology.
References
Bridges, C.B. Triploid Intersexes in Drosophila Melanogaster. Science54, 252-4 (1921).
Erickson, J.W. & Quintero, J.J. Indirect effects of ploidy suggest X chromosome dose, not the X:A ratio, signals sex in Drosophila. PLoS Biol5, e332 (2007).
Cline, T.W. & Meyer, B.J. Vive la difference: males vs females in flies vs worms. Annu Rev Genet30, 637-702 (1996).
Butler, B., Pirrotta, V., Irminger-Finger, I. & Nothiger, R. The sex-determining gene tra of Drosophila: molecular cloning and transformation studies. EMBO J5, 3607-13 (1986).
Boggs, R.T., Gregor, P., Idriss, S., Belote, J.M. & McKeown, M. Regulation of sexual differentiation in D. melanogaster via alternative splicing of RNA from the transformer gene. Cell50, 739-47 (1987).
Inoue, K., Hoshijima, K., Sakamoto, H. & Shimura, Y. Binding of the Drosophila sex-lethal gene product to the alternative splice site of transformer primary transcript. Nature344, 461-3 (1990).
Ryner, L.C. et al. Control of male sexual behavior and sexual orientation in Drosophila by the fruitless gene. Cell87, 1079-89 (1996).
Heinrichs, V., Ryner, L.C. & Baker, B.S. Regulation of sex-specific selection of fruitless 5′ splice sites by transformer and transformer-2. Mol Cell Biol18, 450-8 (1998).
Inoue, K., Hoshijima, K., Higuchi, I., Sakamoto, H. & Shimura, Y. Binding of the Drosophila transformer and transformer-2 proteins to the regulatory elements of doublesex primary transcript for sex-specific RNA processing. Proc Natl Acad Sci U S A89, 8092-6 (1992).
Hoshijima, K., Inoue, K., Higuchi, I., Sakamoto, H. & Shimura, Y. Control of doublesex alternative splicing by transformer and transformer-2 in Drosophila. Science252, 833-6 (1991).
Alpatov, W.W. Phenotypical variation in body and cell size of Drosophila melanogaster. Biological Bulletin58, 85-103 (1930).
Taylor, K. & Kimbrell, D.A. Host immune response and differential survival of the sexes in Drosophila. Fly (Austin)1, 197-204 (2007).
Neckameyer, W.S. & Matsuo, H. Distinct neural circuits reflect sex, sexual maturity, and reproductive status in response to stress in Drosophila melanogaster. Neuroscience156, 841-56 (2008).
Magwere, T., Chapman, T. & Partridge, L. Sex differences in the effect of dietary restriction on life span and mortality rates in female and male Drosophila melanogaster. J Gerontol A Biol Sci Med Sci59, 3-9 (2004).
Rideout, E.J., Narsaiya, M.S. & Grewal, S.S. The Sex Determination Gene transformer Regulates Male-Female Differences in Drosophila Body Size. PLoS Genet11, e1005683 (2015).
Hudry, B., Khadayate, S. & Miguel-Aliaga, I. The sexual identity of adult intestinal stem cells controls organ size and plasticity. Nature530, 344-8 (2016).
Billeter, J.C., Rideout, E.J., Dornan, A.J. & Goodwin, S.F. Control of male sexual behavior in Drosophila by the sex determination pathway. Curr Biol16, R766-76 (2006).
Evans, D.S. & Cline, T.W. Drosophila switch gene Sex-lethal can bypass its switch-gene target transformer to regulate aspects of female behavior. Proc Natl Acad Sci U S A110, E4474-81 (2013).
Finley, K.D. et al. Dissatisfaction encodes a tailless-like nuclear receptor expressed in a subset of CNS neurons controlling Drosophila sexual behavior. Neuron21, 1363-74 (1998).
Shirangi, T.R. & McKeown, M. Sex in flies: what ‘body–mind’ dichotomy? Dev Biol306, 10-9 (2007).
Hartmann, B. et al. Distinct regulatory programs establish widespread sex-specific alternative splicing in Drosophila melanogaster. RNA17, 453-68 (2011).
Shepard, P.J. & Hertel, K.J. The SR protein family. Genome Biol10, 242 (2009).
Gambacciani, M. et al. Body weight, body fat distribution, and hormonal replacement therapy in early postmenopausal women. J Clin Endocrinol Metab82, 414-7 (1997).
Grove, K.L., Fried, S.K., Greenberg, A.S., Xiao, X.Q. & Clegg, D.J. A microarray analysis of sexual dimorphism of adipose tissues in high-fat-diet-induced obese mice. Int J Obes (Lond)34, 989-1000 (2010).
Nakada, D. et al. Oestrogen increases haematopoietic stem-cell self-renewal in females and during pregnancy. Nature505, 555-8 (2014).
Chen, X. et al. The number of x chromosomes causes sex differences in adiposity in mice. PLoS Genet8, e1002709 (2012).
Markle, J.G. et al. Sex differences in the gut microbiome drive hormone-dependent regulation of autoimmunity. Science339, 1084-8 (2013).
Steegenga, W.T. et al. Sexually dimorphic characteristics of the small intestine and colon of prepubescent C57BL/6 mice. Biol Sex Differ5, 11 (2014).
Cook, M.B. et al. Sex disparities in cancer incidence by period and age. Cancer Epidemiol Biomarkers Prev18, 1174-82 (2009).
El Ouaamari, A. et al. SerpinB1 Promotes Pancreatic beta Cell Proliferation. Cell Metab23, 194-205 (2016).
Edgar, B.A. How flies get their size: genetics meets physiology. Nat Rev Genet7, 907-16 (2006).
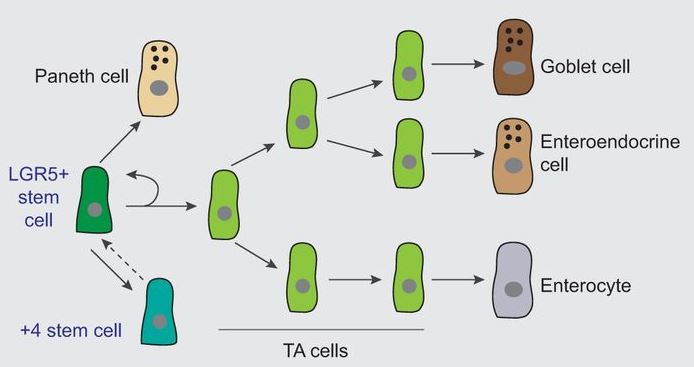
Micchelli, C.A. & Perrimon, N. Evidence that stem cells reside in the adult Drosophila midgut epithelium. Nature439, 475-9 (2006).
Ohlstein, B. & Spradling, A. The adult Drosophila posterior midgut is maintained by pluripotent stem cells. Nature439, 470-4 (2006).
Li, H. & Jasper, H. Gastrointestinal stem cells in health and disease: from flies to humans. Dis Model Mech9, 487-99 (2016).
Regan, J.C. et al. Sex difference in pathology of the ageing gut mediates the greater response of female lifespan to dietary restriction. Elife5, (2016).
Here is some developmental biology related content from other journals published by The Company of Biologists.
Gastrointestinal stem cells in health and disease
This Review from Li and Jasper highlights recent findings on intestinal stem cell (ISC) diversity in the GI tract of Drosophila, focusing on the role of ISCs in healthy and diseased conditions, and drawing parallels with vertebrate GI stem cells. Read the paper here [OPEN ACCESS].
TET-1 in neural tube closure
Lozanoff and colleagues propose an epigenetic mechanism establishing the regulation of genes that are crucial for neural tube closure. This mechanism could be a novel target for resolving such birth defects and associated disorders. Read the paper here [OPEN ACCESS].
Genetic variants of zebrafish muscle myosin
Pack and co-workers report two newly identified Myh11 gene missense mutations discovered in a zebrafish enhancer-suppressor mutagenesis screen. The mutations disrupt myosin regulation and ATPase activity in a graded fashion, and this correlated with their effects on intestinal and vascular physiology. Read the paper here [OPEN ACCESS].
Nuclear migration events in development
Nuclear migration is a central part of many cell and developmental processes. Bone and Starr review the large number of different molecular mechanisms that exist to move nuclei to specific, intracellular locations. Read the paper here.
TrkA mediates Sema3A signalling
Goshima and colleagues show that, by interacting with PlexA4, TrkA plays a crucial role in redirecting local Sema3A signaling to retrograde axonal transport, thereby regulating dendritic GluA2 localization and patterning. Read the paper here.
Epiboly expands the mammalian epidermis
Formstone and co-workers find that the nascent mammalian epidermis spreads to enclose the embryo trunk through a process akin to epiboly, which has important implications for human birth defects of the abdominal wall. Read the paper here [OPEN ACCESS].
Tadpole eyes swivel and cross during metamorphosis
Combes and colleagues analyse adaptive neural plasticity of spinal locomotor-extraocular motor circuit coupling that enables Xenopus frogs to continuously generate effective retinal image-stabilizing eye movements throughout the metamorphic transition from fish-like tadpole to quadrupedal adult. Read the paper here.
Extra arms destabilise sea urchin larvae
Chan and co-workers show that pre-competent, 6-armed larval urchins swim faster and are less stable in experimental turbulent flow than younger 4-armed larvae, suggesting a potential age/morphology-driven differential transport mechanism in ambient flow conditions. Read the paper here [OPEN ACCESS].
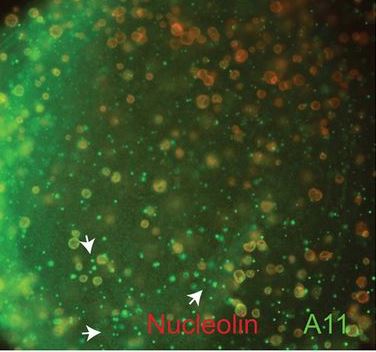
Amyloid assembley in Xenopus oogenesis
Hayes and Weeks find that non-membrane-bound nuclear particles in Xenopus oocytes responsible for RNA transcription, modification and processing contain proteins assembled into amyloids as part of normal development. Read the paper here [OPEN ACCESS].
Most scientific conferences have become large gatherings where little exchange of ideas can actually occur. There are way too many people to start socializing with folks you don’t know and there are so many talks that the idea of sitting through one on a topic on which you are not an expert sounds suicidal. At the other extreme was The Company of Biologists’ “Metabolism in Development and Disease” Workshop. Within a Downtown Abbey-like XVI century house, a few dozen scientists, including early career ones like us, met to discuss the implications of cell metabolism in embryonic development, immune response and disease – particularly cancer.
It was excellent. And not only because of the impressive Wiston House and its surroundings or the great list of invited speakers. It was excellent because this was a true instance where people from disparate fields met, exchanged ideas and learnt from each other. We were no exception -as our backgrounds testify. Patricia is a biochemist that has always worked on metabolism while Carlos is a developmental biologist by training. As early career scientists, it was incredibly exciting to meet and discuss with more seasoned researchers in an open and intimate environment. The pastoral fields that surrounded Wiston House and the only bar of the neighborhood, conveniently located in the same house, also helped to foster friendship and possible collaborations with fellow junior and senior biologists alike.
Wiston House. Photo: Carlos Carmona-Fontaine
The talks were excellent too. There will be an upcoming meeting report to be published in Development (by Aurelio Teleman) that will summarize some of the scientific content of the meeting, so we will focus more on our personal take on the lessons from the meeting. Molecular biologists have pushed for an understanding of cellular functions based on signal transduction and codes – a visionary idea. For example, the conceptualization of the Central Dogma powered by people like Francis Crick and Sydney Brenner was a milestone in the history of molecular biology. It allowed us to simplify the complexity of cellular activities and think them as signaling problems. This concept helped us greatly to advance our understanding of how cells function and interact with each other. This is similar to how graphical interfaces and operating systems have eased the use of computers, as we do not need to deal with abstract programs and complicated coding anymore. There is a downside to this approach however: we could forget how computers actually work. Similarly, we sometimes forget that “cell markers” are enzymes, or nutrient transporters or pH and oxygen sensors. Transcription factors are critical for the development of the embryo (or a tumor) but they are that way because they ultimately encode for functional proteins that direct specific chemical reactions – such as RNA synthesis – in the cell. Why don’t we think where the methyl or acetyl groups are coming from when we talk about histone modifications and the “epigenetic code”? We risk biasing our understanding of cell biology as a collection of codes and signals, rather than actual chemical reactions.
The painting “Metabolism” by Edvard Munch depicts Adam and Eve, a metaphor for the initiators of human’s development. Photo: https://commons.wikimedia.org
The field of cell metabolism is inherently linked with a more biochemical understanding of biological functions. For the last two decades, this field has been reinvigorated by advances in molecular biology and modern genetics.
This synergy has been adopted by a vast number of scientists working in immunology, cancer and many other research fields. Although modern metabolism has also begun to permeate developmental biology, we think that this workshop was absolutely unique and marks the milestone of a hopefully long-lasting interaction between embryology and metabolism. In this meeting, it was clear that the field of development reciprocated with its emphasis on spatial compartmentalization and temporal dynamics of cell metabolism. For example, it was exciting to learn about the efforts to develop and improve machines that allow the detection of metabolites and stable isotopes with a 50nm resolution! We also discussed how nutrients can mediate cell-cell interactions and allow for mutualistic or competitive cell relationships. There is still a long road ahead and these research fields need to cooperate to develop new and better tools to probe complex metabolic systems. For example, promising technologies such as optogenetics will clearly help us to “shine light” onto as yet unknown metabolic networks within and between cells. Although regulation of metabolic networks in certain diseases may sound futuristic, it may well be a crucial step to develop successful personalized therapies – though as we said it’s a long road!
Detail of “Embryology” by Magdalena Abakanowicz is an abstract large installation where shapes that can resemble eggs or early embryos that have achieved different sizes. One could imagine that their differential growth is due to differences in their metabolism. Photo: Carlos Carmona-Fontaine
At the end of the workshop we left with a feeling that cell signals are a key way to understand cell biology, but we cannot forget that ultimately life is about growing and growth requires a exquisite regulation of its basic components: metabolites.
We thank Ben Steventon and Katherine Brown for comments on the text.
Professors Graham Williams and Duncan Bassett have 3 full-time posts (5 years each) available. One technical post in skeletal biology is funded by the Horizon 2020 grant: Resetting the THYRoid axis for prevention of AGE-related diseases and co-morbidities. One post-doc and one technician post in molecular and cellular biology are funded by the Wellcome Trust: Cellular thyroid hormone availability: regulation of development and tissue repair, and pathogenesis of degenerative disease.
Please see our website for this exciting opportunity to join our cutting-edge research program based at Imperial College London.

 (1 votes)
(1 votes)

 In the first study, Andy Fischer and colleagues investigate the role of mTor signalling in the formation of Müller glia-derived progenitor cells (MGPCs) in the chick retina. The authors use NMDA to induce a cytotoxic response, and observe that mTor signalling is transiently activated upon activation of Müller glia cells. Inhibition of mTor signalling in vivo prevents the proliferation of the Müller glia cells and blocks the regenerative response. Using a range of inhibitors and readouts, the authors show that mTor signalling is required for the proliferation of MGPCs. The authors further show that mTor signalling is activated in response to insulin, IGF1 and FGF2, and that this response is most likely independent of the MAPK pathway.
In the first study, Andy Fischer and colleagues investigate the role of mTor signalling in the formation of Müller glia-derived progenitor cells (MGPCs) in the chick retina. The authors use NMDA to induce a cytotoxic response, and observe that mTor signalling is transiently activated upon activation of Müller glia cells. Inhibition of mTor signalling in vivo prevents the proliferation of the Müller glia cells and blocks the regenerative response. Using a range of inhibitors and readouts, the authors show that mTor signalling is required for the proliferation of MGPCs. The authors further show that mTor signalling is activated in response to insulin, IGF1 and FGF2, and that this response is most likely independent of the MAPK pathway. In the second study, Joachim Wittbrodt and colleagues look at the role of a single factor, Atoh7, in directing Müller glia cells to proliferate and differentiate in the absence of an injury. The authors use a fluorescent transcriptional reporter of atoh7 to demonstrate atoh7 expression in proliferating Müller glia cells after retinal injury in medaka. The authors then use an inducible system to activate expression of atoh7 in vivo in the Müller glia cells, and find that this is sufficient to drive the cells to re-enter the cell cycle and undergo proliferation. Forced expression of atoh7 in these cells activates Notch signalling, and indeed the authors show that overexpression of the Notch intracellular domain can recapitulate the effects seen by atoh7 overexpression. Importantly, not only did atoh7 overexpression in Müller glia lead to cell cycle re-entry and proliferation, but the authors also observed the formation of neurogenic clusters and subsequent de novo neurogenesis following atoh7 overexpression in these cells. Together, these two studies bring together novel and exciting findings regarding the regulation of Müller glia proliferation following injury and the subsequent regenerative response.
In the second study, Joachim Wittbrodt and colleagues look at the role of a single factor, Atoh7, in directing Müller glia cells to proliferate and differentiate in the absence of an injury. The authors use a fluorescent transcriptional reporter of atoh7 to demonstrate atoh7 expression in proliferating Müller glia cells after retinal injury in medaka. The authors then use an inducible system to activate expression of atoh7 in vivo in the Müller glia cells, and find that this is sufficient to drive the cells to re-enter the cell cycle and undergo proliferation. Forced expression of atoh7 in these cells activates Notch signalling, and indeed the authors show that overexpression of the Notch intracellular domain can recapitulate the effects seen by atoh7 overexpression. Importantly, not only did atoh7 overexpression in Müller glia lead to cell cycle re-entry and proliferation, but the authors also observed the formation of neurogenic clusters and subsequent de novo neurogenesis following atoh7 overexpression in these cells. Together, these two studies bring together novel and exciting findings regarding the regulation of Müller glia proliferation following injury and the subsequent regenerative response.
 In early 2012, circular RNA (circRNA) was shown to be a transcriptional product in thousands of human and mouse genes and in hundreds of cases constituted the dominant RNA isoform. Subsequent studies revealed that the expression of circRNAs is developmentally regulated, tissue and cell-type specific, and shared across the eukaryotic tree of life, suggesting important functions for these molecules. Here,
In early 2012, circular RNA (circRNA) was shown to be a transcriptional product in thousands of human and mouse genes and in hundreds of cases constituted the dominant RNA isoform. Subsequent studies revealed that the expression of circRNAs is developmentally regulated, tissue and cell-type specific, and shared across the eukaryotic tree of life, suggesting important functions for these molecules. Here,  Root hairs are highly specialized cells found in the epidermis of plant roots that play a key role in providing the plant with water and mineral nutrients. Many studies have shown that the fate of root epidermal cells, which differentiate into either root hair or non-hair cells, is determined by a complex interplay of intrinsic and extrinsic cues. Here, Wolfgang Schmidt and colleagues review these studies and discuss recent evidence suggesting that environmental information can be integrated at multiple points in the root hair morphogenetic pathway and affects multifaceted processes at the chromatin, transcriptional and post-transcriptional levels. See the Review on p.
Root hairs are highly specialized cells found in the epidermis of plant roots that play a key role in providing the plant with water and mineral nutrients. Many studies have shown that the fate of root epidermal cells, which differentiate into either root hair or non-hair cells, is determined by a complex interplay of intrinsic and extrinsic cues. Here, Wolfgang Schmidt and colleagues review these studies and discuss recent evidence suggesting that environmental information can be integrated at multiple points in the root hair morphogenetic pathway and affects multifaceted processes at the chromatin, transcriptional and post-transcriptional levels. See the Review on p.  (No Ratings Yet)
(No Ratings Yet)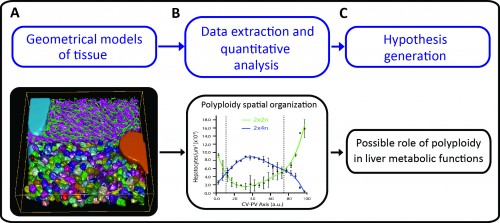

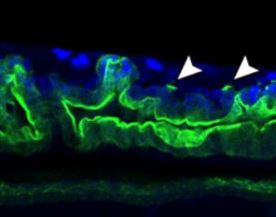 Genetic variants of zebrafish muscle myosin
Genetic variants of zebrafish muscle myosin
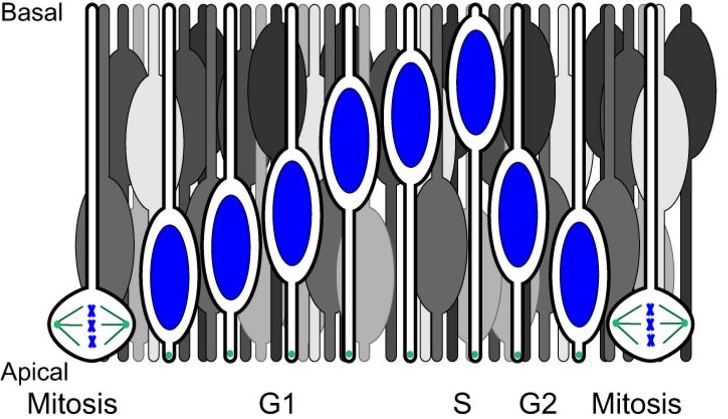 Nuclear migration events in development
Nuclear migration events in development 

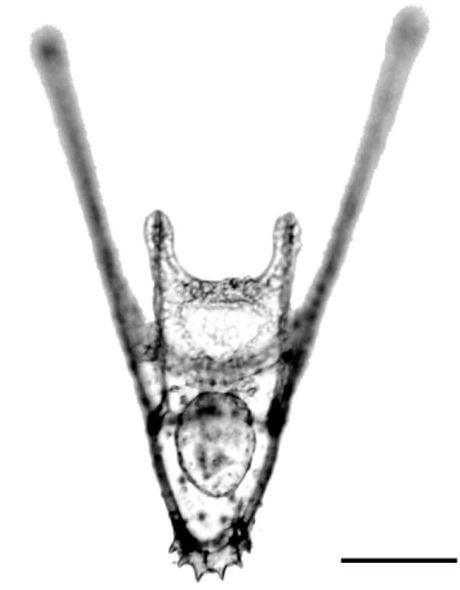 Extra arms destabilise sea urchin larvae
Extra arms destabilise sea urchin larvae
 Amyloid assembley in Xenopus oogenesis
Amyloid assembley in Xenopus oogenesis

