Post-doc position on evolution of non-embryonic developments in urochordates
Posted by Stefano Tiozzo, on 22 May 2016
Closing Date: 15 March 2021
A 15 months postdoctoral position is available at the Observatoire Océanologique de Villefranche-sur-Mer -LBDV (UPMC – Sorbonne Universités). We are seeking a highly motivated and passionate researcher to join a 5-years international project, DEVODIVERSITY, funded by the French Agence Nationale de la Recherché (ANR) and the São Paulo Research Foundation (FAPESP).
By using molecular and cell biology, NGS transcriptomic, genomic, and ecological approaches, a multidisciplinary consortium led by two teams, the Tiozzo Lab at the Villefranche-sur-Mer Developmental Biology Laboratory in France (CNRS-UPMC), and the Brown Lab at the Istituto de Biociências in Brazil (USP), will study the evolution of regeneration, asexual reproduction, and clonality in several species of ascidians (Urochordata), and examine how ecological factors affect distribution ranges, evolution of life cycles and developmental strategies. DEVODIVERSITY has the following main aims:
- To resolve the phylogenetic relationships and evolutionary transitions between strict sexual reproduction to budding and high regenerative abilities among Styelidae (Ascidiacea).
- To provide a coupled morphological and ecological understanding of asexual propagation (budding). We will generate detailed anatomical and developmental descriptions of budding processes, and explore if and which environmental conditions are associated with the use of particular budding modes.
- To compare gene pathways involved in stem cell functions or trans/de-differentiation of budding and regenerative processes by the generation and in silico analyses of transcriptomic data.
- To launch a comparative genomic approach study in order to better understand the evolution of a major life history transition in marine chordates, in particular the evolutionary transition from sexual to asexual propagation.
The candidate will be mainly based at the LBDV (Villefranche sur Mer, France) and may have the opportunity to do laboratory and fieldwork in both countries.
It is ESSENTIAL:
- A doctoral degree in animal biology, evolutionary biology or related disciplines.
- Demonstrated experience in molecular biology, organismal biology and at least bases of bioinformatics and biostatistics.
- Enthusiasm and genuine commitment to research work.
- Demonstrated ability to undertake independent research, flexibility and problem solving skills.
- Excellent communication skills in English spoken/written.
- Ability to collaboratively work on a team.
Expertise in phylogenomics and transcriptomics is highly desirable but not mandatory. The LBDV has an international working environment, speaking French is not required but it would be a plus.
The applicant should contact directly Dr. Stefano Tiozzo (tiozzo@obs-vlfr.fr) providing a letter of interest, a CV and the contact of three potential referees. Please, add “DD_postdoc” on the subject of your email. Applications review will continue until the position is filled. Starting data could be any time from September 2016.
Contacts
Stefano Tiozzo (tiozzo@obs-vlfr.fr): http://biodev.obs-vlfr.fr/~tiozzo/tiozzo-lab.
LBDV: http://biodev.obs-vlfr.fr/en/index.html
 (No Ratings Yet)
(No Ratings Yet)
 (1 votes)
(1 votes)
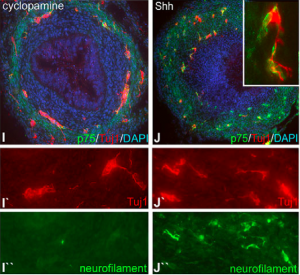





 On Thursday 9 May 1991, the world awoke to front-page news of a breakthrough in biological research. From Washington to Wollongong, newspapers, radio and TV were abuzz with the story of a transgenic mouse in London called Randy. Why was this mouse so special? The mouse in question was a chromosomal female (XX) made male by the presence of a transgene containing the Y chromosome gene Sry. This sex-reversal provided clear experimental proof that Sry was the elusive mammalian sex-determining gene. Twenty-five years on, Koopman, Sinclair and Lovell-Badge reflect on what this discovery meant for our understanding of how males and females arise and what remains to be understood. Read the Spotlight on p.
On Thursday 9 May 1991, the world awoke to front-page news of a breakthrough in biological research. From Washington to Wollongong, newspapers, radio and TV were abuzz with the story of a transgenic mouse in London called Randy. Why was this mouse so special? The mouse in question was a chromosomal female (XX) made male by the presence of a transgene containing the Y chromosome gene Sry. This sex-reversal provided clear experimental proof that Sry was the elusive mammalian sex-determining gene. Twenty-five years on, Koopman, Sinclair and Lovell-Badge reflect on what this discovery meant for our understanding of how males and females arise and what remains to be understood. Read the Spotlight on p.  Mammalian pluripotency is the ability to give rise to all somatic cells as well as the germ cells of an adult mammal. Here, Juan Carlos Izpisua Belmonte and colleagues provide a summary of mammalian pluripotency both in vivoand in vitro, and highlight recent and future applications of pluripotent stem cells for basic and translational research. Read the poster article on p.
Mammalian pluripotency is the ability to give rise to all somatic cells as well as the germ cells of an adult mammal. Here, Juan Carlos Izpisua Belmonte and colleagues provide a summary of mammalian pluripotency both in vivoand in vitro, and highlight recent and future applications of pluripotent stem cells for basic and translational research. Read the poster article on p.  In March 2016, meeting organizers Sebastian Jessberger and Hongjun Song brought together over 100 scientists from around the world to Cancun, Mexico to present the latest research on neurogenesis. Here,
In March 2016, meeting organizers Sebastian Jessberger and Hongjun Song brought together over 100 scientists from around the world to Cancun, Mexico to present the latest research on neurogenesis. Here,