Xenbase, the Xenopus model organism database (MOD), is building it’s developer team.
We a looking to hire 2 software developers and a database administrator. These positions are based at the University of Calgary, at the foot of the Rocky Mountains in Calgary, Alberta, Canada.
Ideally, we like to find people who have some biology background, or have worked in other MODs- (e.g., Zfin, Flybase, MGI) or biology databases (e.g., NCBI, UniProt, BeeGee)
Please share this job posting with anyone you think might be interested.
Xenbase has received a new grant from the NICHD/NIH to fund its expansion over the next 5 years, but due to a rule about advertising jobs beyond the current grant year, the job ads say “7 months with possibility of renewal”. Don’t let this discourage anyone from applying!
New Lectureships are available in the School of Biological Sciences, University of East Anglia, Norwich. We are looking for dynamic individuals who complement and extend the activities in BIO in both research and teaching.
BIO’s research portfolio includes: Cell and Developmental Biology, Cell signalling, Musculoskeletal Biology, Cancer, Matrix Biology, RNA Biology and more information can be found on our Research pages.
Details and links to the application forms can be found on our current vacancies page:
Developmental biologists use antibodies extensively to study the gene expression during different stages. However, there is a lack of specific antibodies against many proteins related to development. In addition, some antibodies yield unspecific and/or irreproducible results. To help alleviate this antibody quality and specificity problem, Labome sought to organize antibody applications cited in formal publication since 2008 and developed Validated Antibody Database (VAD). The most recent version, 2.2, contains manually curated 143357 entries from 38430 formal articles, covering 35146 antibody products from 110 suppliers. The suppliers include both commercial entities and non-profit organizations such as Developmental Studies Hybridoma Bank (DSHB) at University of Iowa, and Neuromab from University of California at Davis. A small number of antibodies from academic researchers are included as well, if these antibodies are validated in knockout models.
One of the side benefits of our curation effort is the identification of cross-reactive species for many antibodies. Antibodies tend to be developed for human/mouse proteins and tend to be tested by commercial suppliers for their applicability in human or mouse system. Development models often use more readily manipulatable models such as flies, worms, zebrafish, and frogs. Labome is able to obtain information about many antibodies having cross-reactivities with the model organisms from the literature.
The database is freely browsable at www.labome.com. Information about antibody validation using knockout models is also posted at Labome Facebook page www.facebook.com/LabomeNews.
Feedback and suggestions are most welcome. We hope to work with everybody to develop a useful tool.
The Max-Planck-Institute for Molecular Biomedicine in Muenster, Germany has an opening for a
Postdoctoral Scientist (position-code 07-2016)
The position is available in the DFG Emmy Noether junior group of Dr. Ivan Bedzhov that is focused on understanding the self-organization of the pluripotent lineage in mammalian embryos at the time of implantation (I. Bedzhov and M. Zernicka-Goetz, Cell, 2014). The successful candidate will investigate the mechanisms of self-organization of the pluripotent epiblast in the context of the developing embryo and in vitro using embryonic stem cell as a model system.
We are looking for a talented and highly motivated post-doctoral scientist with strong cell and molecular biology background. Previous experience with RNA-seq analysis, genome editing and embryonic stem cells will be an advantage.
The Max Planck Institute for Molecular Biomedicine offers dynamic, multidisciplinary environment with state-of-the-art transgenic, imaging, genomics and proteomics equipment and core facilities. The working language in the institute is English. The institute is located in Muenster, a vibrant city with a highly international academic environment.
The position is initially available for two years with the possibility of extension. Starting date will be as soon as possible. All conditions for the employment will be according to the regulations of the contracts for the civil service (TVöD, Tarifvertrag für den öffentlichen Dienst) level 13 TVöD.
The Max-Planck society is committed to increasing the number of individuals with disabilities in its workforce and therefore encourages applications from such qualified individuals. Furthermore, the Max Planck Society seeks to increase the number of women in those areas where they are underrepresented and therefore explicitly encourages women to apply.
Please send your application (with the position-code 07-2016), letter of motivation, CV including a complete list of publications and the contact information of 2 referees to:
career@mpi-muenster.mpg.de
or
Max Planck Institute for Molecular Biomedicine
Roentgenstrasse 20
48149 Muenster
Germany
Last week, Development announced a special issue on organoids. In vitro organogenesis is a burgeoning new field, with applications in the study of human development, drug testing and ultimately the possibility of producing functional organsin the dish that could be used for transplantation. Every new technological advance brings with it a new set of ethical issues, and this is particularly true with regards to brain organoids. Whilst there have been enormous advances in this area, brain organoids are still far from being functional ‘mini brains’. However, it is not impossible that in the near future we may be able to generate a brain organoid that has a sensory area that is able to functionally connect with a processing centre. At what point could we say that these organoids can ‘think’? And does it matter? This month we are asking:
What are the ethical issues surrounding the generation of brain organoids?
Share your thoughts by leaving a comment below! You can comment anonymously if you prefer. We are also collating answers on social media via this Storify. And if you have any ideas for future questions please drop us an email!
Below is an interview with brain organoid researcher Juergen Knoblich, which may be of interest to you:
The nematode Caenorhabidis elegans is among the most widely used and powerful model organisms for studying mechanisms underlying cellular and developmental processes. Although a variety of approaches for conditional protein expression have been developed in C. elegans, available tools for conditional protein depletion are far more limited, particularly in the germ line. We were thus motivated to develop a technique to control the abundance of proteins in living animals, which we felt would be a great addition to the toolbox available in this system.
In the Dernburg lab, we are interested in studying the molecular mechanisms underlying meiosis. In many cases we can use genetic mutations to interrogate the roles of individual proteins, but this approach does not work well if the proteins perform essential functions during mitosis, which is required for proliferation of the germ line. We therefore sought to develop a method that would enable inducible, rapid, and quantitative protein depletion in the germ line. After considering a number of possible strategies, we focused on the auxin-inducible degradation (AID) system, which has been applied in cultured cells and single-celled organisms (Nishimura et al., 2009). This approach was originally adapted from the plant auxin perception system. Inducible degradation relies on a small molecule phytohormone produced by all plants, auxin (indole-3-acetic acid). In addition, it requires TIR1, an F-box protein, which forms part of an Skp1–Cullin–F-box (SCF) E3 ubiquitin ligase complex. In the presence of auxin, TIR1 recognizes peptide sequences (degrons) that are present in a large number of target proteins, mostly transcriptional regulators, expressed by plants, and targets these proteins for polyubiquitylation and proteasome-mediated degradation. We thought this system might be transplantable to nematodes, since auxin is a very small, fairly water-soluble molecule, potentially making it easy to deliver to living animals and readily diffusible through tissues.
A rotation student, Ze Cheng, first accepted the challenge of adapting the AID system to C. elegans. He generated a number of constructs to test its feasibility. Because there is no endogenous auxin receptor in C. elegans, he made a strain stably expressing a modified Arabidopsis thaliana TIR1 from a single copy transgene using MosSCI; (Frokjaer-Jensen et al., 2008). He also made a construct to express a degron- and GFP-tagged SMU-2 from extrachromosomal arrays in the germ line. After treating worms expressing both constructs with auxin for a couple of hours, he was excited to observe a disappearance of the green fluorescent SMU-2::GFP signal, indicating the AID system might indeed be useful in C. elegans.
We then systematically expanded and tested the AID system in C. elegans. To maximize the chances of success, we incorporated two amino acid changes in AtTIR1 that were found to increase its binding affinity for degron-tagged substrates and to thereby increase its sensitivity to auxin (Yu et al., 2013). We created strains expressing TIR1 under control of various promoters and 3’ UTR sequences to drive germ line-specific or temporally regulated expression. We used a 44-amino acid minimal degron sequence derived from Arabidopsis thaliana IAA17 (Morawska and Ulrich, 2013) and tagged proteins of interest with this sequence using CRISPR/Cas9-mediated editing (Dickinson et al., 2013). Gratifyingly, we detected efficient inducible-degradation in the germ line when growing the transgenic worms with auxin-containing liquid culture or plates. Excitingly, rapid degradation was consistently achieved within a reasonable range of auxin concentrations (<1 mM), which did not seem to have any effects on wild-type worms. We also found that the position of the degron was quite flexible; it could be placed at either end of a target protein, or even sandwiched between the target and another tag, such as GFP.
Considering the potential usefulness of the AID system for the research community, we further analyzed the inducible degradation of target proteins in the soma of C. elegans in detail. To do this, we made a variety of tissue-specific TIR1 strains with tissue-specific promoters and 3’ UTR sequences. We then combined these TIR1 transgenes with different types of degron-tagged transgenic targets. We found either cytoplasmic or nuclear proteins tagged with the degron can be tissue specifically degraded at various developmental stages in an auxin concentration dependent manner. After trying a wide range of auxin, we also noticed that the degradation is reversible upon auxin removal, with lower auxin doses accelerating recovery. To our surprise, we further detected efficient inducible degradation of targets in early embryos inside the mother and in laid eggs, suggesting promising usefulness of the AID system for studying mechanisms underlying embryo development. Notably, the inducible degradation is also efficient in the absence of food, making it useful for studying starvation-induced processes, such as autophagy and larvae arrest.
When we shared these findings with some colleagues, they were extremely excited about the potential of the system. One of them, Jordan Ward at UCSF, wanted to test the ability of this system to address key questions underlying nuclear hormone receptor-mediated control of developmental gene regulatory networks. He was able to tag two essential nuclear hormone receptors, NHR-23 and NHR-25, with the degron sequence and found that each could be depleted within 40 min, enabling detailed functional dissection of these proteins during development.
Another exciting finding was that the AID system can produce more penetrant phenotypes than depletion by RNAi, not only in the worm soma but also in the germ line. Given the high efficiency of CRISPR/Cas9-mediated genome editing in C. elegans, tagging proteins of interest with the 44-amino acid degron is now quite easy and fast, further augmenting the utility of the AID system for cell and developmental studies in worms.
We have made the AID-related worm strains and plasmids available through the CGC and Addgene, respectively. A large number of worm laboratories have already tried the AID system to deplete proteins of interest in C. elegans. We are getting great feedback from our colleagues, several of whom have told us that the AID system works spectacularly in their hands. In principle, this approach may be applicable to a wide range of other organisms. We thus look forward to seeing the application of this technology in not only worm labs but also other metazoan model organism labs.
Here is some developmental biology related content from other journals published by The Company of Biologists.
Drosophila as a model to study human disease
The latest issue of Disease Models & Mechanisms highlights the translational impact of Drosophila research. In this issue, Moulton and Letsou review several Drosophila models of human inborn errors of development (read here), while the poster and review by Bellen and colleagues examine some of the tools and assays available in Drosophila to study human disease (read here).
Nanog suppresses senescence
Thummer, Edenhofer and colleagues analysed the outcomes of Nanog gain-of-function in various cell models employing a recently developed cell-permeant version of this protein. They show that Nanog blocks cellular senescence of fibroblasts through transcriptional regulation of cell cycle inhibitor p27KIP1. Read the paper here [OPEN ACCESS].
Skeletal development
Panx3 and Cx43 are two important gap junction proteins expressed in osteoblasts. Yamada and colleagues show that Panx3 and Cx43 regulate skeletal formation through their distinct expression patterns and functions. Read the paper here.
Asymmetric cell division in the worm
Phillips and colleagues show that two Dishevelled paralogs have both redundant and non-redundant roles in β-catenin regulation during asymmetric cell division in C. elegans. Read the paper here.
MOBP in oligodendrocyte differentiation
Myelin-associated oligodendrocytic basic protein (MOBP) resembles myelin basic protein (MBP), but the signals initiating its synthesis and function remain elusive. In this paper, White and colleagues show, by several approaches in cultured primary oligodendrocytes, that MOBP synthesis is stimulated by Fyn activity, and reveal a new function for MOBP in oligodendroglial morphological differentiation. Read the paper here.
How bacteria can affect development
Mushegian and colleagues examined the effect of temperature and presence of bacteria in diapausing eggs of the water flea Daphnia magna. They show that the presence of bacteria increases successful development of resting eggs at an elevated temperature. Read the paper here.
Water deprivation affects snake development
Dupoué and colleagues examined the effects of water availability on corticosterone secretion in breeding snakes. They show that water deprivation induces an increase in baseline corticosterone level in pregnant aspic vipers, which may subsequently influence offspring growth. Read the paper here.
Controlling tissue polarity
Planar cell polarity sigalling directs the polarization of cells within the plane of many epithelia. Sharp and Axelrod examine the signals that Prickle and Spiny-legs respond to and the mechanisms they use to control the direction of tissue polarity in the distal wing and the posterior abdomen of Drosophila. Read the paper here [OPEN ACCESS].
From fibroblasts to neural crest
Motohashi and colleagues identified the transcription factors specifically expressed in developing mouse neural crest cells, and showed that SOX10 and SOX9 directly converted fibroblasts into neural crest cells. Read the paper here [OPEN ACCESS].
We have PhD position available to start in October 2016 at the University Göttingen (3 yrs, 75% E 13 TV-L).
The main aim of the project will be to study intra-specific variation in compound eye size in Drosophila melanogaster. The successful PhD candidate will address this question by analyzing the genetic basis of eye size variation in various inbred strains of the Drosophila melanogaster Genetic Reference Panel (DGRP). For all DGRP lines genome sequences are available. For representative strains, developmental transcriptome data using RNAseq will be generated so that data across several scales (genome, transcriptome, phenotype) can be integrated. Similar data will be generated for artificial selection experiments based on a subset of the DGRP fly lines. The successful candidate will work in an interdisciplinary team at the Department of Animal Sciences (Prof. Dr. Henner Simianer) and the Department of Developmental Biology (Dr. Nico Posnien) as well as during an extended research stay in the group of one of the international collaborators overseas.
Please visit the website of the Research Training Group “Scaling Problems in Statistics” (http://www.uni-goettingen.de/en/156579.html) for more details about the general setting of this position.
A detailed description of the project, information about the application procedure and further requirements are available here: http://tinyurl.com/gn3az25
Please forward this job ad to all your motivated future PhD students.
I am Aaron Harnsberger, a second year Master’s degree student in the Department of Biological Sciences at Idaho State University in Pocatello, Idaho. The focus of this lab is on genetic regulatory divergence that results in the diversity of mammalian morphologies. These morphological differences can be observed at various stages of development. In this lab we use bats as our model organism. There are no other members working in this lab besides myself. There are a growing number of labs throughout the world that also use bats to study evolutionary and developmental biology. The species of bat that I use is Carollia perspicillata (C. perspicillata) or simply Carollia (Fig. 1). On average an adult Carollia bat is about 19 grams (Fig. 2). Although it is possible to maintain Carollia as a laboratory bat (Rasweiler et al., 2009), we do not have a bat colony in the lab so before I can conduct experiments I make trips to the field to collect animals from the wild.
Figure 1: Carollia perspicillata held by John Rasweiler.
At this point, you might be asking, “Why use bats?”. Bats have some very interesting adaptations that make them a great model for studying evolutionary development. They are the only mammal to achieve powered flight. The development of wings where other mammals have paws, fins, hooves or hands is one adaptation that I find fascinating. Another very interesting adaptation discovered in bats is delayed development. This is when a pregnant female delays development of the embryo. In other mammalian species this typically occurs at the blastocyst stage before implantation in the uterus. However, in Carollia delayed development occurs after implantation at the gastrulation stages (Rasweiler et al., 1997). This delay may be highly synchronous in colonies, and cues that initiate the delay and end it are still unknown. Bats can have a long life span when compared to other mammals of similar size. In addition, bats may respond differently to injury and the healing process may lack inflammation. There are more features, but as you can see bats are unique models for many biological questions.
Figure 2: C. perspicillata held by Aaron Harnsberger. Carollia is a phyllostomid (leaf-nosed) bat.
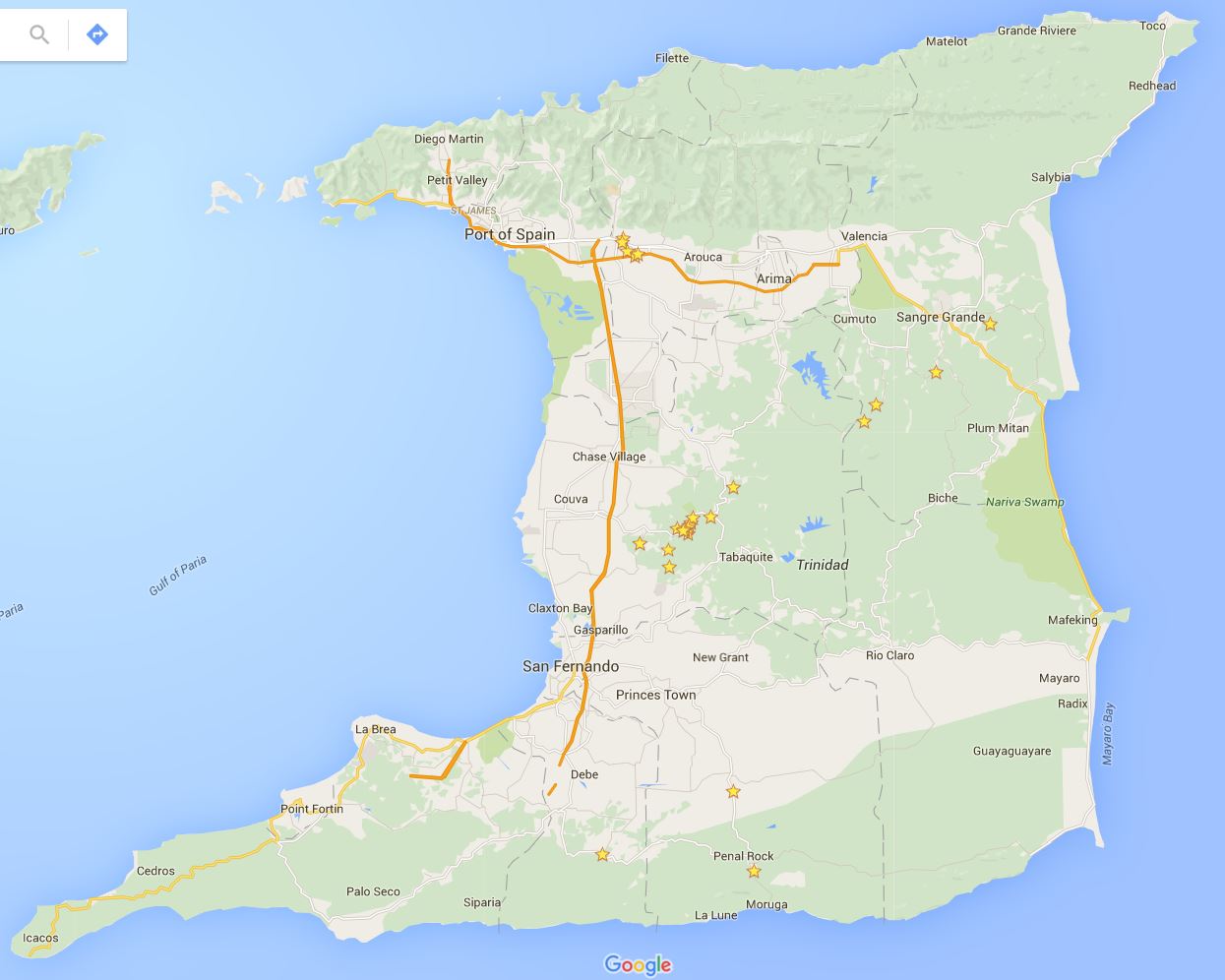
Back to the field, which in this case, is the country of Trinidad and Tobago (Fig. 3). These field collection trips to the island of Trinidad require careful planning and weeks of time to catch bats and collect embryos. I do not catch the bats on these trips by myself. The ‘Bat Team’ this year consisted of John Rasweiler, Richard Behringer, Simeon (Patsy) Williams, Joseph Truechen and myself. Patsy and Joseph are local field assistants who help us find and collect the bats. The ‘Bat Team’ has consisted of many other biologists that have made this collection trip throughout the years, starting with the initial trip in 2000.
Figure 3: Map of Trinidad taken from Google Maps. We collect Carollia from the northern and central regions of Trinidad.
First and foremost, a collection permit through the Wildlife Section of the Division of Forestry of Trinidad and Tobago is required. We give them a specific number of bats we would like to collect and plan our trip according to this number. Next we need a place to process our samples. This is the Department of Life Sciences at the University of West Indies (UWI) in St. Augustine that has lab space, microscopes and other facilities. We also need lodging since the trips take several days. This is arranged through University Housing. Finally, we rent a car to take us into the deep forest that has enough room for several passengers and our gear. Once all of this is in order we can begin catching bats.
Figure 4: Joseph, John and Patsy bringing gear up the hillside to catch bats.
To catch bats, we have to travel into the northern or central areas of the island. We look for certain habitats that can lead us to specific roosting sites. To achieve this, we conduct scouting trips and spend many hours driving, parking, looking around, taking notes, and if all goes well catching bats.
Figure 5: Field box used to store live bats for safe transport to the lab.
We use lots of different gear to catch bats. We occasionally need ladders to get into some of the places that bats roost, for example abandoned concrete water tanks (Fig. 6). On the occasions that we do need a ladder we have to carry it in, sometimes through thick forest (Fig. 4). We use nets similar to butterfly nets to catch the bats, but much longer to reach high places where they roost (Fig. 6). After catching the bats, we sort through them, releasing males and checking females for age and stage of pregnancy using external visual appearances. We keep these female bats in a field box which allows for safe transport of the bats (Fig. 5). After a morning of collecting, we return to the lab at UWI with the bats to isolate the embryos. Once embryos are dissected we process them for various types of analyses, including morphological, histological and molecular studies. To bring the bat embryos back to the lab in Idaho, we obtain an exportation permit, also from the Wildlife Section of the Division of Forestry of Trinidad and Tobago.
Figure 6: C. perspicillata roosting inside an old abandoned concrete water tank. These types of water tanks have a convenient opening on the top that we enter using a ladder. The bats are seen here cohabitating with cockroaches, just visible in the lower edge of picture.
Now that I have the bat embryos to work with I can spend time in the lab back in Idaho conducting experiments. I use whole-mount in situ hybridization (WISH) to examine the spatial distribution of transcripts for specific genes at different stages of development. Using the Carollia embryo staging techniques (Cretekos, et al. 2005), I can identify embryos at specific stages of development. This gives me specific time points to look for gene expression at critical developmental stages (Fig. 7). Many times these expression patterns are similar yet different from those in the mouse.
Figure 7: Fgf8 expression in Carollia stage 14 embryo after WISH.
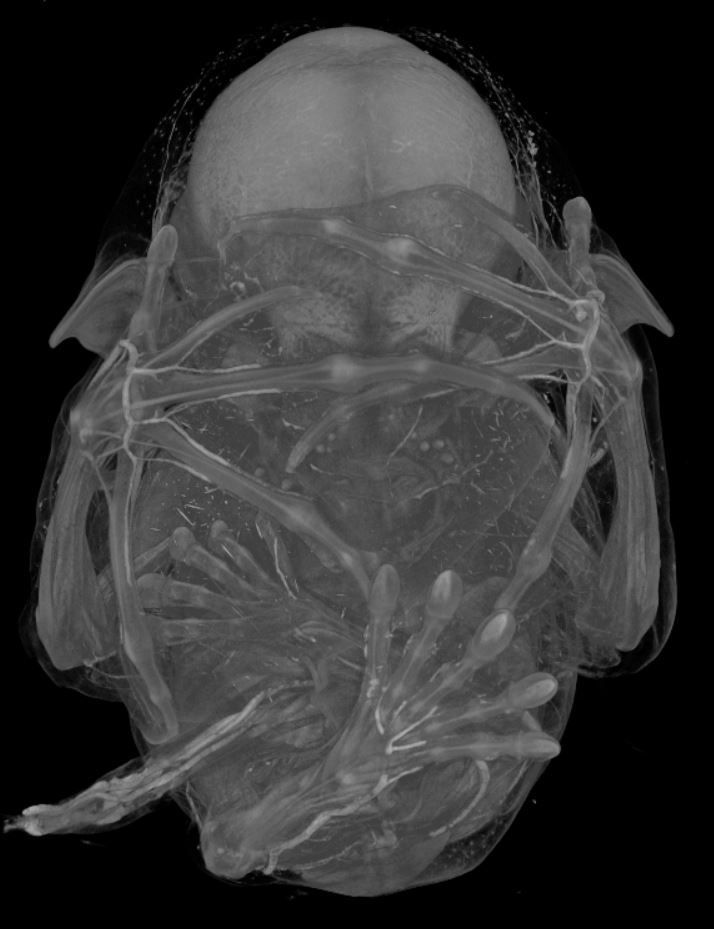
I have also used micro computed tomography (uCT) to visualize the tissue anatomy of bat embryos (Fig. 8). A uCT scan is a high resolution x-ray that is used to make a three-dimensional image. The differences in contrast allow me to identify different tissue types in the bat embryos. Again, taking advantage of the Carollia staging system, I have uCT scanned stages of development in C. perspicillata to examine particular tissues of interest for my project. The data collection was completed in a few weeks’ time. The image processing and analysis of this data has been an ongoing project in the lab.
Figure 8: Carollia stage 20 uCT image.
As you can see a day in the life of a bat lab is not usually just a day, and some of the work may not be typical in other developmental biology labs (Fig 9).
Figure 9: The 2016 ‘Bat Team’. From left; Richard Behringer, John Rasweiler, Simeon (Patsy) Williams & Aaron Harnsberger.
Tags: Bat, Carollia, WISH, in situ, uCT, microCT, development, ISU, Idaho State University
This post is part of a series on a day in the life of developmental biology labs working on different model organisms. You can read the introduction to the series here and read other posts in this series here.
One MSc or PhD student position is available for a September 2016 start date to study mechanisms determining the degeneration or the proliferation of neurons using the D. melanogaster fruit fly model. The two complementary projects will use common assays to investigate the following questions: i. How does calcium and redox signaling between the Endoplasmic Reticulum (ER) and mitochondria determine neuronal degeneration and inflammation. ii. How does the neurofibromatosis type 2 gene product Merlin determine neuronal proliferation. The project is part of a collaboration between the Simmen and Hughes laboratories that aims to further use the fruit fly model organism for the characterization of the fundamental cell biology behind neurodegeneration and childhood tumors of the nervous system. A publication list and more information about the labs can be found on these two sites
The location of the research project is at the University of Alberta, the 5th largest University in Canada with world-class biomedical research labs. The University of Alberta Cell Biology graduate program is amongst the best Cell Biology programs worldwide. Edmonton is a culturally vibrant, young city that lies in close proximity to the Rocky Mountains, offering excellent opportunities for sports enthusiasts.
Minimal requirements are competitive grades from an internationally acclaimed University. Applicants need to provide a cover letter that outlines their career objectives and why they would like to enter a PhD program, CV and scanned university grades. Outstanding applicants from Europe are encouraged to apply, but must have excellent English knowledge both spoken and written (high TOEFL/GRE scores or equivalent). Successful applicant will have to enter the University of Alberta Cell Biology graduate program. Detailed instructions for prospective applicants can be found here
 (No Ratings Yet)
(No Ratings Yet) (1 votes)
(1 votes)
 The latest issue of Disease Models & Mechanisms highlights the translational impact of Drosophila research. In this issue, Moulton and Letsou review several Drosophila models of human inborn errors of development (read
The latest issue of Disease Models & Mechanisms highlights the translational impact of Drosophila research. In this issue, Moulton and Letsou review several Drosophila models of human inborn errors of development (read 
 Thummer, Edenhofer and colleagues analysed the outcomes of Nanog gain-of-function in various cell models employing a recently developed cell-permeant version of this protein. They show that Nanog blocks cellular senescence of fibroblasts through transcriptional regulation of cell cycle inhibitor p27KIP1. Read the paper
Thummer, Edenhofer and colleagues analysed the outcomes of Nanog gain-of-function in various cell models employing a recently developed cell-permeant version of this protein. They show that Nanog blocks cellular senescence of fibroblasts through transcriptional regulation of cell cycle inhibitor p27KIP1. Read the paper  Skeletal development
Skeletal development MOBP in oligodendrocyte differentiation
MOBP in oligodendrocyte differentiation
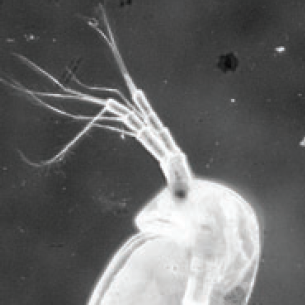 How bacteria can affect development
How bacteria can affect development
 Planar cell polarity sigalling directs the polarization of cells within the plane of many epithelia.
Planar cell polarity sigalling directs the polarization of cells within the plane of many epithelia.  From fibroblasts to neural crest
From fibroblasts to neural crest 





 This post is part of a series on a day in the life of developmental biology labs working on different model organisms. You can read the introduction to the series
This post is part of a series on a day in the life of developmental biology labs working on different model organisms. You can read the introduction to the series