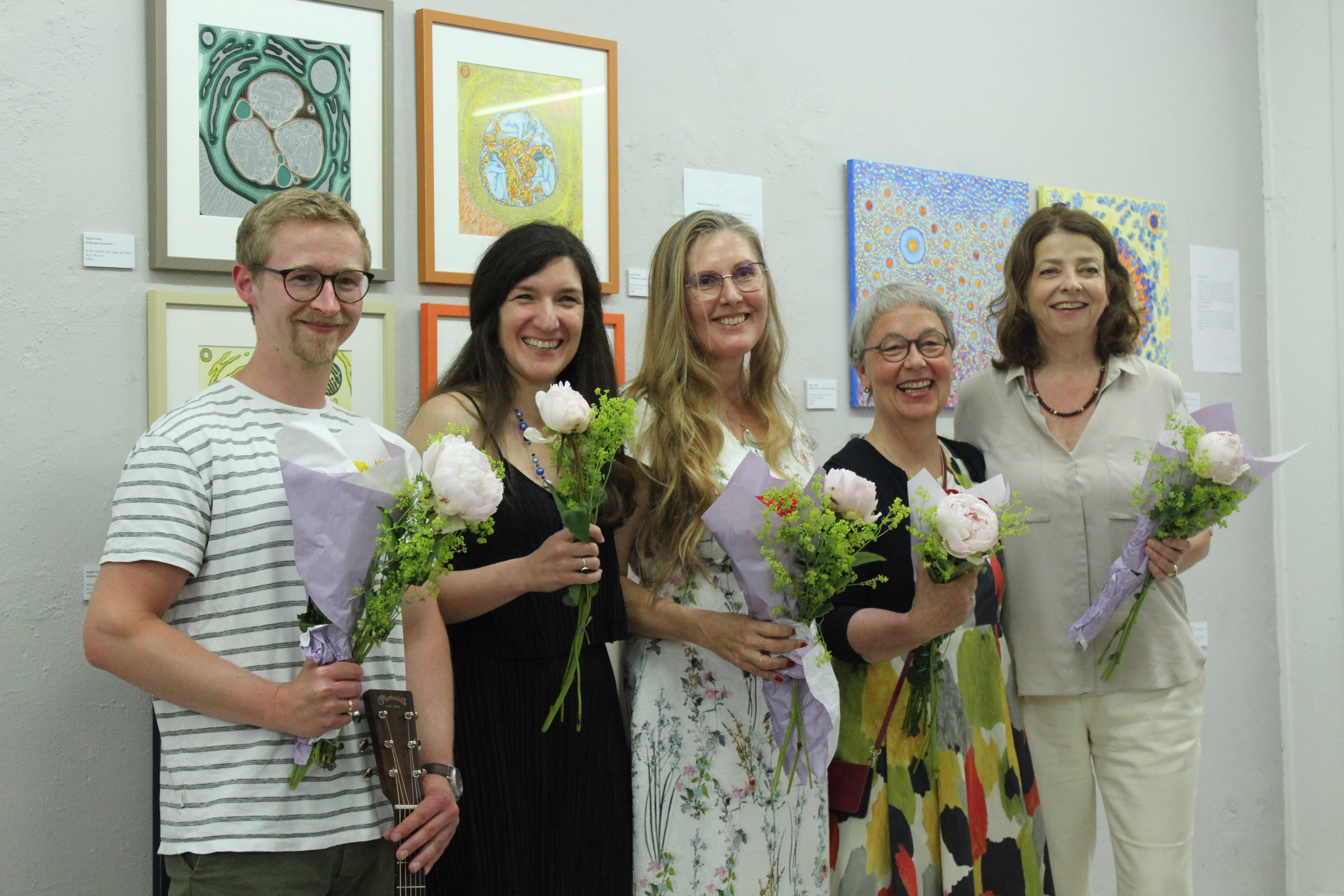The recent Special Issue from Journal of Cell Science focussed on the cell biology of mitochondria. To showcase some of the research in this issue, FocalPlane and MITOtalks have teamed up to host a webinar on Thursday 3 July at 16:00 BST (17:00 CEST, 11:00 EDT, 08:00 PDT). MITOtalks organisers Nuno Raimundo and Sjoerd Wanrooij have invited Yuli Buckley, Mireia Nager and Dikaia Tsagkari to present their research.
Prof. Caren Norden (@nordenlab.bsky.social) appointed Darwin Professor of Animal Embryology in the Department of Physiology, Development, and Neuroscience at the University of Cambridge.
Dr. Júlia Peloggia de Castro (@jupeloggia.bsky.social) awarded the Behrensen – Guzmán Palma Award for the most outstanding research paper by a predoctoral student at Stowers Institute for Medical Research.
Special thanks to Maddie Ryan, Charli Corcoran & Michaela Noskova Fairley for putting this digest together! If you would like to thank the Zebrafish Rock! team for their time & effort, you can buy us a strong cuppa at the link below. Every little bit keeps us caffeinated and motivated! We appreciate your support 🙂
Do you have news, jobs or research that you want to add to the next digest but don’t have a social media account? Use the submission form at our website below:https://linktr.ee/zebrafishrock
Our July webinar features three early-career researchers working on early embryogenesis and will be chaired by Development’s Executive Editor, Alex Eve.
André Dias (Universitat Pompeu Fabra) ‘Opposing Nodal and Wnt signalling activities govern the emergence of the mammalian body plan’
Nikhil Mishra (Institute of Science and Technology Austria) ‘Geometry-driven asymmetric cell divisions pattern cell cycles and zygotic genome activation in the zebrafish embryo’
At the speakers’ discretion, the webinar will be recorded for viewing on demand. To see the other webinars scheduled in our series, and to catch up on previous talks, please visit: thenode.biologists.com/devpres
In this SciArt profile, we find out more about Henning Falk, who did a PhD in developmental biology, and now enjoys creating science-related cartoons as a freelancer.
[Click on the images to view the full-size versions of the cartoons]
Can you tell us about your background and what you work on now?
I studied molecular biotechnology and completed my PhD at EMBL Heidelberg, where I focused on somitogenesis in mouse embryos. Microscopy has always been my favorite technology, and during my PhD project, I developed a method to live-image mouse embryo development for up to three days. After finishing my PhD, I left research and transitioned from microscope development to CNN-based image analysis, and eventually to intelligent document processing using large language models. I currently work as a project manager for a company that builds document processing solutions. Creating cartoons for science communication serves as my connection back to my former researcher self.
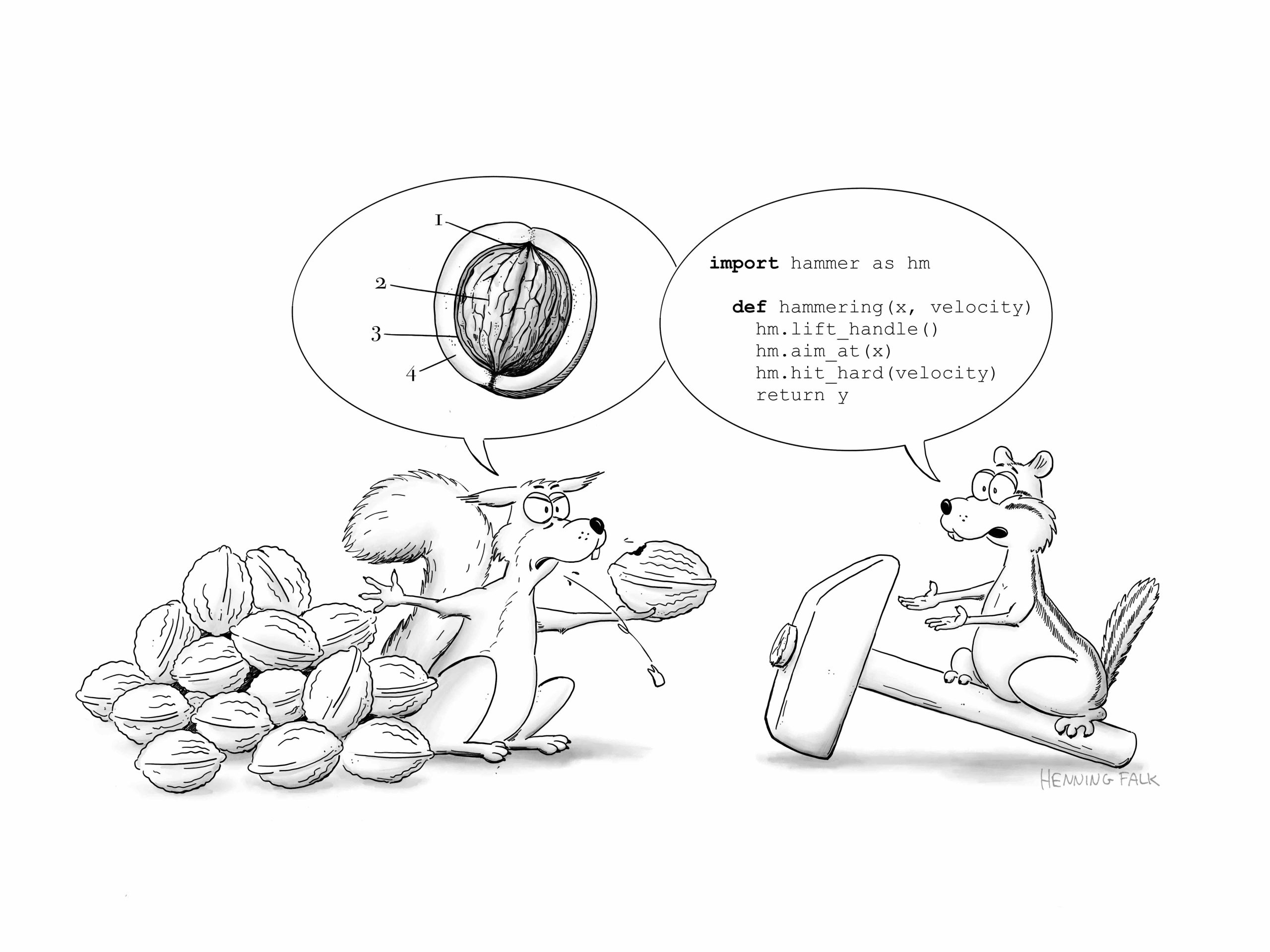
The cartoon was done for German BioImaging (GerBI-GMB) to illustrate the enabling power of research data management (RDM)
Were you always going to be a scientist?
No, my early career ideas went all over the place, though they did include both biologist and artist – sometimes leaning more toward one, other times toward the other. Now I’m a customer AI project manager – life certainly takes surprising turns sometimes.
Yes, absolutely. In kindergarten, I gave myself the self-imposed task of drawing one picture every day to bring home. During primary school, I was a regular participant in local painting competitions. Later, I focused more and more on cartoon drawing – I’m not really sure why, but somehow it stuck with me.
This cartoon is part of a series of post cards raising awareness for the work of the NFDI4BIOIMAGE consortium. Published under a CC BY 4.0 licence.
What or who are your most important artistic influences?
My biggest influences are other cartoonists who manage to convey their message – their joke – in just one simple drawing and a sentence or two. For me, that’s the real essence of good cartoons: condensing a story so drastically that one image is enough to give you the background story, introduce the characters, and deliver a punchline. It’s pretty remarkable. Ralph Ruthe and Joscha Sauer (maker of “Nichtlustig”) are two German cartoonists I find quite inspiring and funny.
Does your science influence your art at all, or are they separate worlds?
I started creating scientific cartoons for friends and colleagues when I was still working on my PhD, and I also used drawings in my own presentations to illustrate my results and theories. I received a lot of positive feedback. A few years ago, I began doing scientific cartoons and illustrations as a freelancer. So today, it’s not my science influencing my art, but rather the science of my “clients.” I find it tremendously fun to work together with researchers to figure out exactly what message they want to convey and how to transform that abstract concept or complicated experimental result into a simple drawing. It’s a tug of war between completeness and simplicity, and sometimes we go a long way together – from a page-long list of detailed sub-results to nailing down the 2-3 main takeaway points that fit into one cartoon. This is where my background in science, my love for cartoon drawing, and my project management skills come together nicely. Voilà!
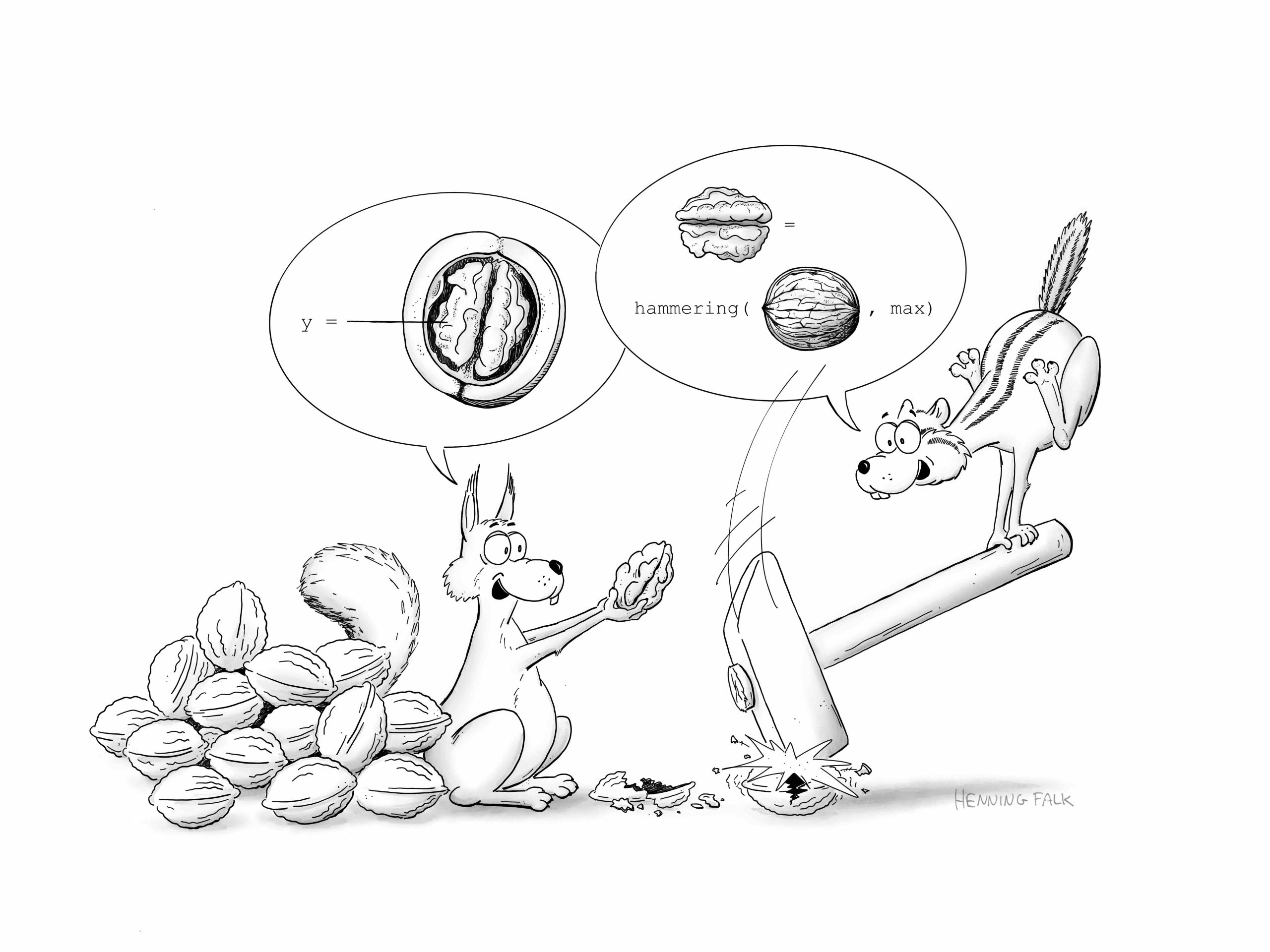
Figure for a publication depicting the challenges bio imaging scientists are faced with. Proper handling of large-scale, complex data is a challenge for researchers, data providers and users. https://f1000research.com/articles/11-638
I just finished a project for a psychologist, and before that, there were several projects in biological data management. As for what comes next, I don’t know yet.
This figure was created for a talk by Christina Derksen as basis to talk about the best practices for the development of clinical decision support systems (CDSS) in medicine by highlighting the most common pitfalls.
How/where can people find more about you?
I have a small website advertising my services and showcasing some of my recent projects. It’s falk-illustrations.de.
As mammals, we like to think of ourselves as dominant animals, and to some extent, that is true. Humans, specifically, have altered nearly every part of our planet and reshaped the climate. We’ve domesticated cattle and chickens for food and bred cats and dogs for pets. We exterminate countless other species across the tree of life. Yet, we and our mammalian relatives were not always in control, and throughout life’s history, our influence has ebbed and flowed.
Dr. Caleb Gordon, a paleontologist in the Earth and Planetary Sciences Department at Yale University, spent several years unraveling a dynamic tale that began 300 million years ago. At that time, “the biggest animals on land were our ancestors—the sprawling, sharp-toothed cousins of mammals, and the biggest animals in the oceans were giant armored or fleshy-finned fishes,” Caleb speaks enthusiastically. “The earliest reptiles appeared around this time too,” he introduces them with a smile. “Reptiles are a big, quirky family with lizards, crocodiles, turtles, dinosaurs, and birds. They include many other species, both living and extinct, that descended from an ancient common ancestor. Reptiles were diminutive little creatures in a world dominated by bigger, scarier animals who probably ate them whenever they found them.” On land, reptiles lived in the shadows of large mammal ancestors, eating insects and other small prey. They were not thriving in the oceans either, where various big fish were the top predators.
Then, some 50 million years later, a sudden surge of volcanic activity and environmental upheaval transformed the landscape and the plot. “The Great Dying, we call it, triggered the worst mass extinction in the history of animal life, around 252 million years ago,” Caleb explains in a more serious tone. “Up to 95% of species were wiped out, and almost anything bigger than a modern-day deer – gone.” But reptiles were lucky. In the aftermath, they thrived, both in aquatic and terrestrial arenas. In their “glow-up era – the Age of the Reptiles,” they became the new top predators on land and in the oceans.
Back in the present day, I sit in a local New Haven coffee shop with Caleb as he draws an evolutionary tree in his notebook. He shows me when reptiles and mammals first evolved and how they have competed throughout Earth’s history. “It’s hard for many of us, at least here in the northeast U.S., to imagine encountering big reptiles today. If we hike through a national park or drive through our local neighborhood, most of the big animals we’ll see are mammals — deer, cows, bears, dogs, wolves, coyotes, and especially humans. And if we swim in the cold coastal waters, most of the big animals we’ll see are sharks, bony fish, or whales,” he says. For his dissertation, Caleb explored a different landscape. He wanted to understand how, in the aftermath of the Great Dying, reptiles managed to win both on land and in sea as the Age of Reptiles began.
“There is an underappreciated role for the evolutionary innovations that helped reptiles win,” he explains. By evolutionary innovations Caleb means “complex features they evolved that allowed them to do new things and invade new environments after the Great Dying.” Such traits appear randomly, and on rare occasions, favoured their survival in certain circumstances. Little by little, through small changes over many years and generations, these beneficial traits became long-lasting innovations in reptiles.
To transition to water and survive the rough seas, reptiles needed special evolutionary innovations. “Marine reptiles were, over the course of millions of years, able to change the dimensions of their arms to create their own flippers. They did this by making their hands much longer while shortening their lower arm bones.” Caleb demonstrates by shrugging his shoulders close to his ear, squeezing his arms by his body, while extending and flapping his hands out. “Reptiles were able to do this quickly and easily, a bunch of times, in a bunch of different groups. But in mammals and their ancestors, flippers didn’t appear for another 200 million years.” At this broad time scale, mammals were worse at making flippers, and it rarely happened. While flippers evolved some eight to ten times in reptiles, they only evolved four times in mammals. In part because of their flippers, some aquatic reptiles evolved to become massive creatures that could now prey upon the biggest sharks.
To thrive on land, one group of reptiles managed to shape their skulls to catch big prey,” Caleb says with a smile, “imagine, a T. rex, the biggest meat-eating animal that ever walked the Earth, or its relative, the modern-day saltwater crocodile, which has one of the strongest bites among all living animals today.” To become terrifying carnivores, terrestrial reptiles needed to open their jaws wide, snap it shut quickly, and bite down hard, destroying their prey before it escaped. Caleb demonstrates this by cupping his hands open and shut like a clamshell. But nature gives and takes, and powerful jaws tend not to open as wide or bite down as fast, while nimble and lightweight jaws open wide, but usually fail to bite down as hard. Reptiles managed to overcome these limitations, Caleb explains, by “changing the architecture of their jaw muscles.” Shortly before the Great Dying, some transformed their tiny deep jaw muscles into massive ones that could open wider while maintaining a powerful bite. These reptiles were the ancestors of dinosaurs. They ate bigger prey, became fearsome land predators, and reached a new level of carnivory.
Reconstructing such details of a distant past is not easy. “Studying the evolution of ancient reptiles is tricky. We rely on scarce fossil records they left behind, and the vast majority of them left no trace at all.” To make these discoveries, Caleb does some of the classic paleontology work you may envision from the movies. He sifts through ancient bones at the Yale Peabody Museum and precisely records lengths, widths, heights, and depths. With these, he built the largest dataset of its kind: around 16,000 measurements from over 800 museum specimens, representing both living and extinct species, across hundreds of millions of years of evolution.
Caleb Gordon collecting data at the Yale Peabody Museum. Here, he measures an arm bone of an ancient marine reptile that lived during the Jurassic Period, over 140 million years ago.
“But these classic methods can only get you so far. To understand all of these fossils, and all this information, we need to use modern techniques and technologies.” He feeds his masses of data into a machine-learning algorithm to compare competing stories of whether extinct species occupied land, water, or both, and predict which scenario was most likely. Given the nature of the data, Caleb had to introduce a statistical method first devised by the US Navy, and almost never used to study fossils. “We also adapt 3D computer animation programs, the same software used by Pixar and Marvel Studios, to ‘repair’ and ‘un-crush’ fossils so that they look more like they would have when the animal was alive,” he explains. These approaches let us recraft ancient structures that are unseen in modern life. With these methods, Caleb can tell which past aquatic and terrestrial reptile stories are likely true.
3D skull segmentation in action.Caleb segments a skull bone of a Euparkeria, a close relative of crocodilians and dinosaurs.
Caleb’s work recreates scenes from prehistoric times of life forms fighting for their existence. His PhD advisor, Dr. Bhart-Anjan Bhullar, emphasizes, “Caleb’s research represents a series of fundamental advances in our understanding of the lives of ancient organisms.” These ancient organisms, the diverse family of reptiles, flew, crawled, swam, and roamed. Through luck and remarkable evolutionary innovations, they managed to conquer two vastly distinct environments and replace our ancestors at the top of the global food chain…at least for a while.
This work was supported by training from Carl Zimmer, and feedback from Zili Shen at the Yale University Graduate Writing Lab.
Recently, I attended ‘Shaping Life 3’, the quadrennial meeting of the French Society of Developmental Biology (SFBD). The first of these meetings happened in 2016: that’s where my current PI presented his work, with the excitement reaching me through my lab mates back then, who attended the meeting. Since then, attending these meetings has been on my bucket list. The second iteration took place late into the pandemic (2021), at a time when I still wasn’t feeling courageous enough to resume attending conferences: I guess, better late than never.
The meeting took place in the picturesque town of Cassis, famous also for its wine and the local vineyards. This was a bit of a homecoming meeting for me: I did my PhD in Marseille, a few kilometers from the meeting venue. My PhD institute, IBDM (Developmental Biology Institute of Marseille), being one of the organizing partners, there were many familiar faces in the meeting, showcasing their science.
Being a developmental biology conference, I felt more at home than ever. The international membership of SFBD was also reflected in this conference, with many participants coming from abroad, including outside Europe. The conference showcased the-latest-and-greatest in classical model systems as well as emerging ones, and covered topics from the origin of multicellularity to the development of the distributed central nervous system in Octopus. The diversity of the topics covered in the meeting was only rivalled by the diversity of the organisms, with 2 full sessions on ‘Evo-Devo and Emerging models’. In my opinion, this also mirrors a shift in the field of developmental biology, where doing experiments in non-model systems is becoming mainstream, with comparative analysis across multiple species gaining traction. It felt like the future – where such studies are included in more classical sessions like ‘morphogenesis’ – is closer than ever.
Exceptionally, only a quarter of the talks were from invited speakers; the rest were selected from submitted abstracts, with many of the speakers traveling from abroad, like me. This is the largest proportion of ‘selected talks’ I have seen. Naturally, this means more effort on the organizers’ side, going out of their way to do the difficult job of rejecting abstracts for oral presentation. Personally, I also appreciated that many speakers showed a lot of unpublished results; posters also had a similar ubiquity of unpublished/preprinted results: a welcoming sign to see preprinting becoming commonplace.
A highlight of the conference was the SFBD PhD prize seminar, by Gabriela Poliacikova, who walked us through the unusual function of Hox genes and their cofactor M1BP in Drosophila flight muscle development, during adult stages (see her publication here). She masterfully conveyed the excitement in her PhD project: reflected in the roaring applause that followed her talk.
In conjunction with all the science, we also had a Career Development session, especially for Early Career Researchers, with a lot of discussions on the uncertainties and anxieties associated with the current job market and how to navigate them. Our group decided to hold this session on the beach, chatting with Eve Seuntjens and Wolfgang Keil. Both of them have had exceptional experiences, with Wolfgang emphasizing the importance of widening our areas of expertise, and Eve advising us not to give up on our dreams (see also her interview on the Node).
Being a recent member of SFBD, I was also looking forward to the ‘General Assembly’ meeting during the conference, though with a bit of apprehension: I only know ‘survival-French’ – fading progressively in the backdrop of exposure to German – and wasn’t sure if that would be enough to follow discussions without bothering someone for help with translation. To my surprise, this session was in English, and to my ignorance, it has been so already since the previous few iterations (including the presentation slides): a clear sign that (at least) the community of French Developmental Biologists is steadfast in welcoming international participants, especially in the current turbulent times.
The meeting also struck a nice balance between intense scientific sessions and free time to digest it all. By the end of the meeting, many participants, including me, felt overwhelmed by the flood of information, rescued only by the walks on the beach and small excursions to the Calanques nearby. With minor exceptions, the weather was on our side, with plenty of sunshine and fresh Mediterranean breeze. The Welcome cocktails and Gala dinner on the rooftop of the conference venue were a welcome nicety enjoyed by everyone.
In case you missed it, the building on the left (the one with all the flags) was the conference venue. (5 votes) Loading...
From Saturday 31.05.2025 to Friday 27.6.2025, an art gallery in Heidelberg, Germany, presents an interdisciplinary exhibition combining art and science. The exhibition features the work of Dr. Ayelen Valko, a visual artist and cell biologist who explores the cellular universe to express its beauty and mystery to the general public. Through a blend of artistic techniques, metaphor and symbolism, Ayelen makes complex biological concepts accessible and engaging. The Node previously featured Ayelen in our SciArt profile. Here, we caught up with Ayelen again to find out more about her art, motivations and experiences as a scientist in the art world.
Flyer for the exhibition at GEDOK Heidelberg, showing one of the works from the ‘Anthropo-Lysosomes’ series.
How did your interest in combining cell biology and visual art begin?
I have been painting and drawing my whole life. The natural world has always been my inspiration. My family often tells me that, as a child, I would spend hours drawing leaves and insects in my grandmother’s garden. However, it wasn’t until I began my Ph.D. program and was trained in various microscopy techniques that I realized how much artistic inspiration I could draw from microscopy images.
I have a dual background in art and science. I took several courses at the National University of Arts in Argentina, focusing on portrait and abstract painting and exploring traditional media, such as oil on canvas and acrylics. I then specialized in scientific and naturalistic illustration, developing skills in watercolor, scratchboard, graphite, and ink through courses and workshops at institutions such as the Ernst Haeckel Scientific Painting Lab, the Adumbratio Scientific Illustration Center, and Aves Argentinas.
Today, I use these and other techniques in mixed-media collages and paintings, which are mostly inspired by cell biology.
Vernissage of the exhibition.
Ayelen giving the opening speech.
Why is the microscopic universe an interesting subject for an art exhibition?
To me, understanding the biological processes that sustain life can help us to appreciate it more fully. The microscopic world holds a quiet, yet powerful kind of beauty—both artistic and philosophical. Although it is not a common theme in the arts, the complex and fragile systems that keep cells alive offer endless inspiration. This makes the microscopic universe a fascinating subject for an exhibition: it invites viewers to look closer, to discover an unseen dimension of life that is usually reserved for scientists. Through my work, I aim to capture the essence of these biological processes and bring them to life on canvas in a way that makes them more accessible to everyone. I try to show the hidden beauty of cellular landscapes as I see them, filtered through my own experiences and imagination. My paintings and collages blend scientific ideas with artistic expression, shaped by memories, emotions, and the subconscious. In essence, my work brings together biology, art, and psychology.
… it invites viewers to look closer, to discover an unseen dimension of life that is usually reserved for scientists.
Works depicting various organs, tissues, and biological processes.
What is the idea behind the exhibition?
The concept behind this exhibition is to take the viewer on a visual journey that begins with whole organisms and gradually descends into the inner world of organs, tissues, individual cells, and intracellular compartments—revealing the wonders hidden within a single cell.
To achieve this, I used a wide range of materials and techniques, including inks, colored pencils, fineliners, watercolors, oils, acrylics, and markers, working on paper, canvas, and fabric. The exhibition also features collages made from diverse materials such as thread, fabric, newspaper, tissue paper, cardboard, plastic beads, and sand—or combinations of these.
One section of the exhibition is dedicated to scientific journal covers. There, viewers can see how some of my artworks have been adapted into journal cover illustrations, and read a selection of art-and-science articles I have written for some of those publications.
As the exhibition moves into the realm of subcellular landscapes, I present two ongoing series: Anthropo-Lysosomes and Origins in Blue. These explore opposing biological phenomena—destruction and creation, respectively—through metaphor and symbolism. In both series, I incorporate anthropomorphic elements into the cellular environments. I believe this helps make complex biological concepts more accessible than they might be in more purely abstract representations.
In Anthropo-Lysosomes, I focus on the cell’s digestive compartment—the lysosome. Across the series, three anthropomorphic figures appear inside the lysosome and undergo a progressive visual degradation, symbolizing biological breakdown and psychological vulnerability. I use a variety of techniques to emphasize different facets of destruction in each piece.
The Anthropo-Lysosomes series.
In contrast, Origins in Blue zooms into the mammalian ovary, starting from the organ level and moving toward a single oocyte. Along the way, the imagery becomes increasingly fantastical, blending imagined cellular structures with emotional and symbolic layers. This series explores not only anabolism—biological synthesis—but also interpersonal relationships, particularly themes of otherness, judgment, and inherited conflict. It touches on early childhood and the entry of a new individual into a world shaped by unresolved tensions.
The Origins in Blue series.
How does your background in biology shape your artistic expression, and why do you bring personal experience into that mix?
While it is common to see art used as a tool for science outreach, such as on journal covers or in graphical abstracts, it is much less common to see the relationship reversed, with science becoming a source of inspiration for artistic creation. This inverse perspective is precisely where my work is rooted. As both an artist and a biologist, I strive to depict the beauty and mystery of the cellular universe through my own subjective lens and lived experience. Consider the Surrealists, such as Remedios Varo and Frida Kahlo: They drew inspiration from psychology and the unconscious, but they weren’t trying to educate their audience about Freud or Jung. I asked myself, “Why not use the cellular and subcellular worlds as artistic inspiration without always aiming to explain or teach science?” I believe there’s real artistic value in that approach—finding inspiration in science for expression, not just communication.
As both an artist and a biologist, I strive to depict the beauty and mystery of the cellular universe through my own subjective lens and lived experience.
What challenges have you encountered moving between the scientific and artistic worlds?
Overall, it has been a very positive experience for me. Creating cover illustrations for scientific journals and participating in science communication activities, such as giving talks in schools and running art-and-science workshops, have been deeply rewarding. I find that these kinds of projects offer clearer pathways: you can submit a cover proposal or apply to speak at an outreach event. They’re more structured and predictable ways of combining art with science.
However, finding my place in the art world has been more challenging, especially when trying to exhibit science-inspired work in galleries and museums. This path—less clearly defined—has brought me a great deal of satisfaction, but also some real obstacles.
While most people are curious and supportive, there’s still some confusion about my work. The scientific community sometimes asks me why I “paint” science instead of conducting bench research. In the art world, science is often seen as too rigid, even contradictory to artistic freedom. There’s a lingering assumption that scientists might not be flexible or imaginative enough to truly belong in the arts. The tension of working between disciplines is very real. I often feel like I belong to both artistic and scientific worlds, but also to neither.
Another challenge I face is my preference for creating handmade works rather than digital ones. This can be limiting when approaching certain institutions that focus on “sciart” but lean heavily toward digital aesthetics. They often seek pieces resembling glowing, rotating DNA holograms seen in science fiction films, not for their meaning, but for their visual effect. In those spaces, form tends to outweigh substance, and my more intimate, tactile approach doesn’t always align with the curatorial vision.
Additionally, most art residencies and exhibition programs at scientific or artistic institutions that intertwine art and science are designed for artists or scientists who want to collaborate with the other discipline, but rarely for individuals who belong to both fields.
The tension of working between disciplines is very real. I often feel like I belong to both artistic and scientific worlds, but also to neither.
The exhibition seen from the front of the gallery during a reading event by other GEDOK artists (and guest artists), as part of the organization’s ongoing cultural activities.
How did you find this art gallery and why you thought it was a good venue for your work?
I first came across the GEDOK Heidelberg gallery while walking through the city. I saw the exhibition they were showing at the time and thought it could be a good fit for my own work. So I waited for the next open call for new members, applied, and was accepted.
GEDOK is an artistic community in Germany — in fact, it’s the oldest and largest interdisciplinary network for female artists and art supporters in Europe. It brings together creators from the visual arts, music, literature, and beyond. Next year, the organization will celebrate its 100th anniversary, which speaks to its truly rich history. It has branches in 23 cities across the country, and I’m a member of GEDOK Heidelberg.
After becoming a member, I also applied for the opportunity to present a solo exhibition. A jury of artists and specialists accepted my proposal and gave me very encouraging feedback.
The exhibition was curated by Angelika Wild-Wagner and Anne Arend-Schulten, curators and artists from GEDOK, who accompanied me throughout the entire process. They helped me shape the exhibition concept and supported me during the installation. They also encouraged me to create a two-meter-long DNA sculpture that explores the theme of nature versus nurture, which is now part of the exhibition. It was a very enriching collaboration.
I also received support from Sabine Friebe-Minden, a graphic designer and fellow GEDOK artist. She developed the entire visual concept for the exhibition advertisement.
From left to right: Dr. Oliver Pajonk (a scientist and musician who provided the evening’s music), Dr. Ayelen Valko (the artist behind the exhibition), Sofie Morin (a GEDOK writer who also performed at the opening), and the curators Anne Arend-Schultenand Angelika Wild-Wagner (also GEDOK artists).
Oliver performing some of his original guitar compositions.
How has the public reacted to your work?
So far, the response has been very encouraging. The opening was well attended by a diverse audience.
Those with a scientific background often enjoy recognizing familiar cellular elements and biological processes. Some are drawn to scientific illustrations depicting different model organisms and organs, while others prefer more abstract cellular and molecular landscapes. Ultimately, it depends on each person’s interests and tastes.
Those without a scientific background tend to appreciate the works for their shapes, colors, and composition. Many engage with the idea of imagining the universe inside a cell, which opens the door to science communication and popularization.
Some visitors are also especially moved by the metaphors and symbolism. They connect with the psychological or emotional aspects of the work, demonstrating how the pieces can resonate on multiple levels.
Visitors discussing the artworks.
What’s next for you?
I’m currently looking for new venues to exhibit my work—such as scientific institutions, museums, and art galleries—and I hope to collaborate more closely with both artists and scientists.
I also plan to finish the two series I mentioned earlier and to continue exploring new biological themes in my artwork.
In addition, I’d like to engage more deeply in science outreach activities. I particularly enjoy working with schools: I’ve given virtual talks to students in Argentina, my home country, and those experiences have been especially rewarding.
As part of this outreach effort, Lis Albert, a science communicator and PhD student at Heidelberg University, and I will lead a science communication activity for high school students at the gallery. On June 23, ninth-grade students from Helmholtz-Gymnasium Heidelberg will participate in the session with their teacher, Silke Reinhardt. We will use my science-inspired art as a starting point to talk about cell biology and laboratory work.
At some point, I’d also love to connect with publishers to develop a book on art and science. It’s an idea I’ve been thinking about for a while, and one I’m eager to explore further.
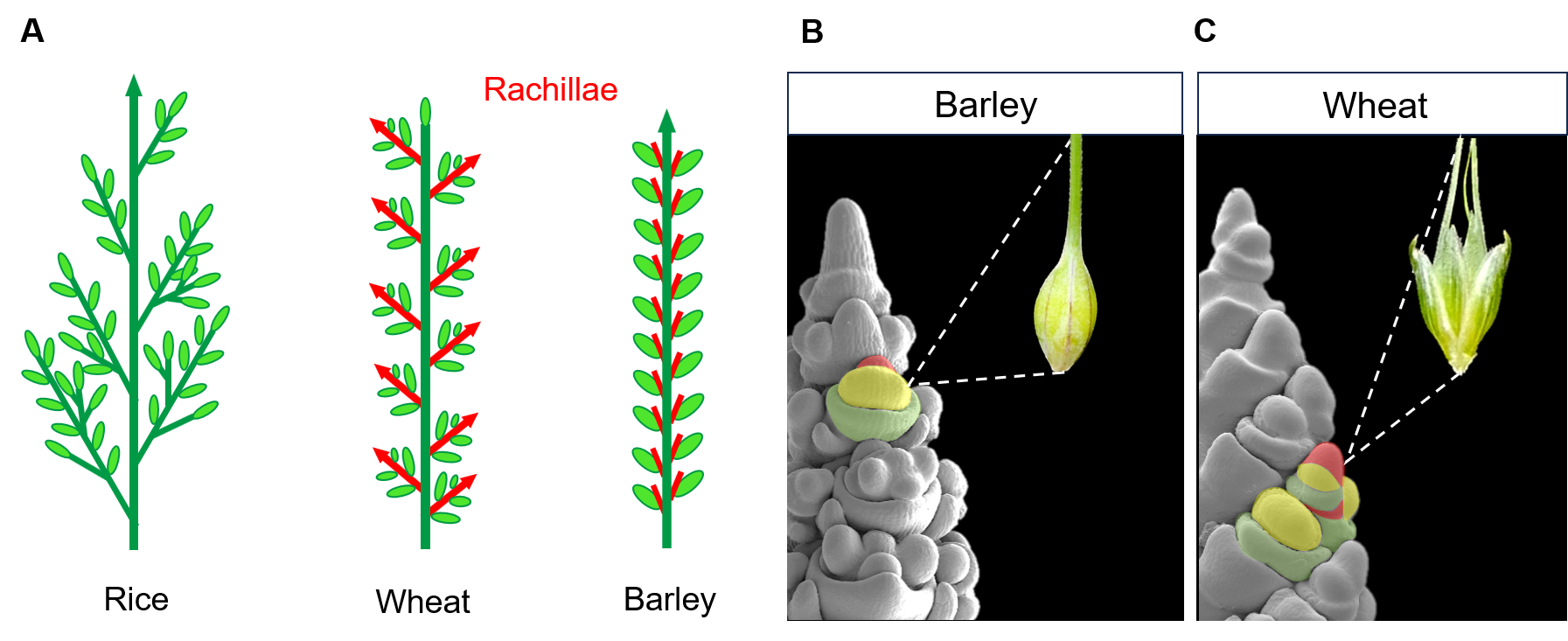
Grasses play a crucial role in human food production and are primarily cultivated for their edible grains. The quantity of grain a grass species can produce is closely linked to the architecture of its inflorescence. Over the course of evolution, grasses have developed a wide variety of inflorescence architectures. From the complex branched inflorescences of the Oryzae tribe (e.g. rice), where multiple grains develop on primary and secondary branches, to the simple spike-type inflorescence of the Triticeae tribe (e.g barley and wheat), where single grains develop on short vestigial axes called rachillae1 (Fig.1 A).
These architectural differences are established during the early stages of the plant’s development by the activity of meristems, specialised structures leading organ growth. The size, position, and lifespan of these meristems determine the eventual shape of the inflorescence2. An example of how small differences in meristem activity can significantly impact the final inflorescence architecture is found within the Triticeae tribe. In barley, the rachilla primordium grows just enough to form a single floret and grain. In contrast, in wheat, its prolonged activity allows for the formation of multiple florets per spikelet, ultimately resulting in its characteristic multi-grain spikelet1 (Fig.1 B,C).
Figure 1: (A) Schematic representation of different grass inflorescence architectures. Main stem and branches in dark green, grains in light green and rachillae in red. (B,C) Scanning electron microscope images of barley and wheat inflorescences at early stages of development. Different organs comprising a spikelet are coloured (rachilla primordium in red, floret meristem in yellow and lemma primordium in green). Each spikelet develops into a single grain in barley and multiple grains in wheat.
When I joined Rüdiger Simon’s lab to begin my PhD, the group was primarily focused on understanding the role of CLE signalling pathways in regulating shape, size, and maintenance of shoot and root apical meristems in Arabidopsis thaliana. At that time, they had begun extending their research to the cereal plant barley. Gwendolyn Kirschner, a former PhD student in the lab, had started investigating the role of CLE-peptide signalling in barley by generating fluorescent reporter lines, including the barley orthologs of the Arabidopsis CLE40 peptide (HvFCP1) and CLV1 receptor (HvCLV1), which had previously been shown to regulate stem cell fate in Arabidopsis meristems3,4. While Gwendolyn primarily analysed barley roots, my project focused more on shoot apical meristem and inflorescence development.
In comparison to the simple inflorescence of Arabidopsis, grasses evolved a more complex organisation, with different meristem types leading to the formation of various organs comprising the spikelet, the basic unit responsible for the development of grains in cereals. This observation led us to the questions: “How is the shape and activity of all these different meristems regulated and coordinated to generate specific inflorescences in grasses? Did barley evolve specific CLE/CLAVATA signalling pathways to regulate the activity of different meristem types?”
HvCLV1 regulates meristem activity along the vertical and lateral axes of the barley inflorescence.
I began by mutating the closest ortholog of CLV1 in barley (HvCLV1) as well as other closely related receptors, which we are still investigating. In maize, as in Arabidopsis, mutation of CLV1 or its ortholog leads to a drastically enlarged inflorescence meristem, resulting in the disorganised formation of additional primordia4,5. In the barley Hvclv1 mutant, I initially observed occasional formation of extra grains within the inflorescence. However, detailed analysis by scanning electron microscopy revealed that an enlarged inflorescence meristem generated an additional row of spikelet primordia, organised in a spiral phyllotaxis rather than in a disorganised manner (Fig. 2A, B).
Moreover, I noticed that Hvclv1 inflorescences developed an elongated rachilla primordium, which produced additional florets per spikelet, an effect previously observed in barley mutants as multiflorus2.b and INTERMEDIUM-m6,7. These results led me to conclude that the ectopic grains generated by the Hvclv1 mutant were due to increased activity of the inflorescence meristem along the vertical axis and of the rachilla primordium along the lateral axis. To further support this, I imaged mature rachillae from WT and Hvclv1 plants and observed the formation of a meristem-like structure at the tip of the Hvclv1 rachilla, which developed into an actively growing small branch instead of the vestigial hairy structure seen in WT (Fig. 2C, D).
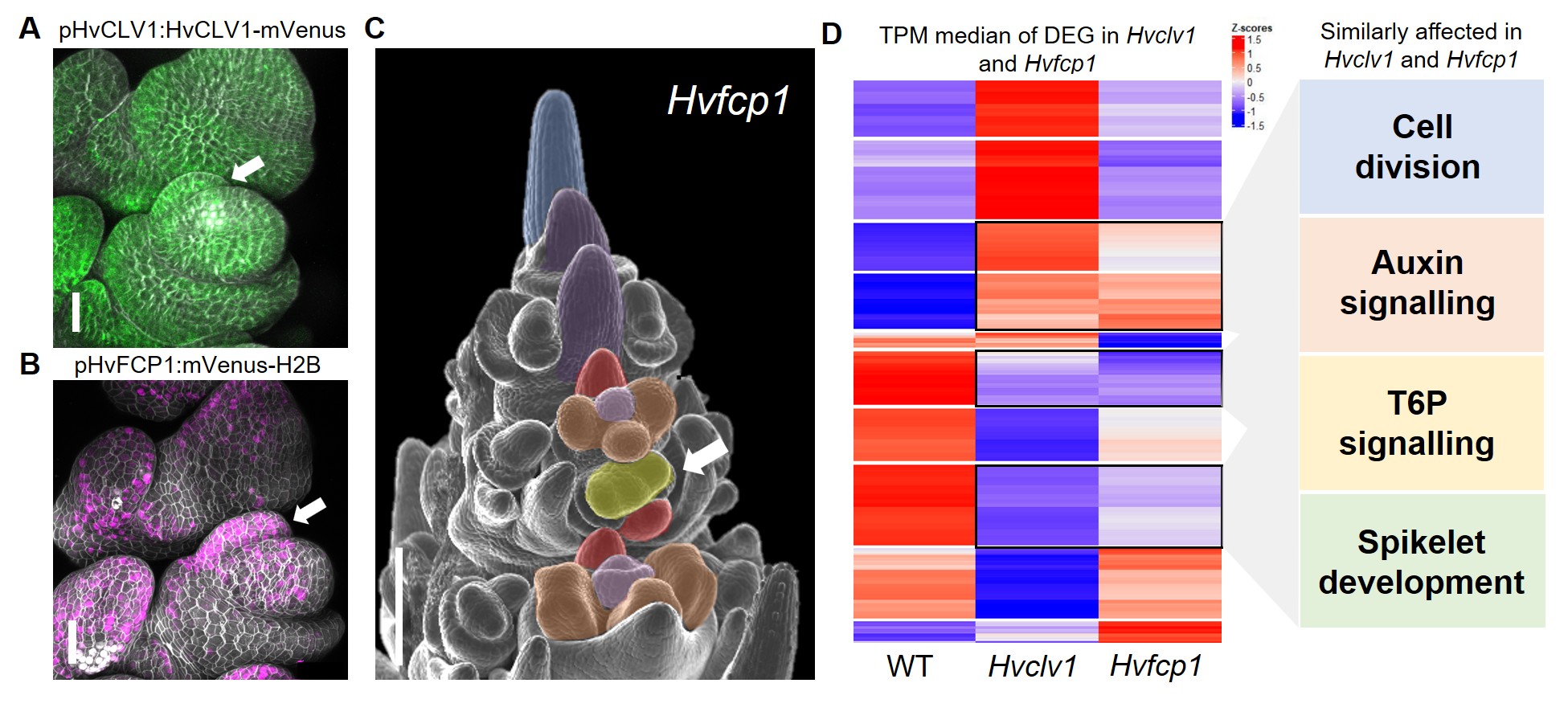
Figure 2: (A,B) Scanning electron microscope images displaying spikelet phyllotaxis (dashed lines) in WT (A) and Hvclv1 (B) early inflorescence tips, combined with photos of the respective final inflorescences. (C,D) Scanning electron microscope images of WT and Hvclv1 early inflorescences. Colours were used to highlight different meristems and primordia (inflorescence meristem in blue, spikelet meristem in violet, rachilla primordium in red, floret meristem in yellow, anther and carpel primordia in brown and pink). Red dashed lines indicate the mature rachilla, combined with zoom-in images where the white arrow indicates the meristem-like structure identified in Hvclv1. Figure modified from Vardanega et. al 2025.
The CLE peptide HvFCP1 acts with HvCLV1 to restrict rachilla primordium activity to the formation of a single floret.
Once the function of the HvCLV1 receptor was characterised, I wondered whether the regulation of rachilla growth was determined by the binding of a specific CLE peptide. Barley possesses 28 different CLE peptides, but I was specifically seeking one that is strongly conserved among grasses and may have contributed to the drastic reduction of branch size in Triticeae. Upon reviewing the literature, I realised that only one peptide retained the same protein sequence across all studied grass species: FCP18. HvFCP1 is the closest ortholog to the Arabidopsis CLE40, for which we already had a fluorescent reporter line. When I examined the expression of HvFCP1 during barley inflorescence development, I noticed that it was not only co-expressed with HvCLV1 but also specifically expressed in the rachilla primordium (Fig. 3 A,B). The insensitivity of the Hvclv1 shoot apical meristem to HvFCP1 peptide treatment, along with the formation of an elongated rachilla producing additional florets even in Hvfcp1 mutants (Fig. 3 C), ultimately demonstrated that HvFCP1 interacts with HvCLV1 to regulate rachilla activity in barley, thereby determining its specific inflorescence architecture.
Figure 3: (A,B) Confocal microscope pictures of barley spikelets displaying HvCLV1 protein localisation (green) and HvFCP1 promoter activity (magenta). The white arrow indicates the rachilla primordium. (C) Hvfcp1 inflorescence displaying double florets (white arrow). (D) RNA seq results. The heatmap shows the z-score of median Transcripts Per Million (TPM) values for each of the Differently Expressed Genes (DEG) in Hvclv1 vs WT and Hvfcp1 vs WT. Black rectangles indicate similarly differentially expressed genes between Hvclv1 vs WT and Hvfcp1 vs WT, with the affected biological processes on the side. Figure modified from Vardanega et. al 2025.
I then investigated which genes are directly or indirectly regulated by HvFCP1/HvCLV1 by performing RNA sequencing on mutant inflorescences compared to wild type (WT). The transcriptome analysis revealed several similarly differentially expressed genes (DEG) between Hvclv1 vs WT and Hvfcp1 vs WT, involved in processes such as cell division, auxin signalling, and trehalose-6-phosphate signalling, providing possible target genes that we are currently investigating (Fig.3 D).
Translating knowledge and techniques from model plants to crops.
In addition to its biological novelty, this paper represents a successful example of translational research, bridging techniques and knowledge from model species to agronomically significant crops. We applied various microscopy techniques more commonly used in model plants, such as Arabidopsis thaliana, but less frequently employed in cereal biology. We developed reporter and complementation lines and quantified receptor cytoplasmic internalisation in rachilla and floret meristems. Furthermore, we utilised methods such as 3D reconstruction and smRNA-FISH for detailed phenotypic analysis of the inflorescence meristem and its expression patterns.
To build on this approach, we generated BARVISTA (http://purl.org/barvista/home), a dataset providing transcriptional information for each cell within the barley inflorescence by integrating single-cell RNA sequencing data with spatial transcriptomics results. Using this resource, we identified transcription factors involved in establishing the specific patterns of meristem ontogenesis necessary to shape the characteristic morphology of the barley spike9.
In conclusion, our work shed light on the signalling pathways that regulate the shape and behaviour of individual meristem types within the inflorescence, paving the way for future efforts to engineer inflorescence architecture through targeted regulation of distinct meristem activities.
REFERENCES:
Koppolu, R. & Schnurbusch, T. Developmental pathways for shaping spike inflorescence architecture in barley and wheat. Journal of Integrative Plant Biology61, 278–295 (2019).
Kyozuka, J., Tokunaga, H. & Yoshida, A. Control of grass inflorescence form by the fine-tuning of meristem phase change. Current Opinion in Plant Biology17, 110–115 (2014).
Schlegel, J. et al. Control of Arabidopsis shoot stem cell homeostasis by two antagonistic CLE peptide signalling pathways. eLife10, e70934 (2021).
Bommert, P. et al. thick tassel dwarf1 encodes a putative maize ortholog of the Arabidopsis CLAVATA1 leucine-rich repeat receptor-like kinase. Development132, 1235–1245 (2005).
Koppolu, R. et al. The barley mutant multiflorus2.b reveals quantitative genetic variation for new spikelet architecture. Theor Appl Genet135, 571–590 (2022).
Zhong, J. et al. INTERMEDIUM-M encodes an HvAP2L-H5 ortholog and is required for inflorescence indeterminacy and spikelet determinacy in barley. Proceedings of the National Academy of Sciences118, e2011779118 (2021).
Goad, D. M., Zhu, C. & Kellogg, E. A. Comprehensive identification and clustering of CLV3/ESR-related (CLE) genes in plants finds groups with potentially shared function. New Phytologist216, 605–616 (2017).
Demesa-Arevalo, E. et al. Imputation integrates single-cell and spatial gene expression data to resolve transcriptional networks in barley shoot meristem development. 2025.05.09.653223 Preprint at Biorxiv https://doi.org/10.1101/2025.05.09.653223 (2025).
We are pleased to support The Pavilion for People, running alongside the 2025 UN Ocean Conference. Explore the space to learn more and share your thoughts on saving our oceans.
As part of The Pavilion for People, we will host a webinar on Thursday 12 June at 11am (BST) on ‘How to organise events more sustainably’. The webinar is free to join. Access the schedule a few minutes before the event and follow the instructions on how to join it.
During the webinar, we will detail the latest best practice advice, as well as providing information on our available resources for creating sustainable events including our new carbon event calculator, coming soon. We look forward to meeting you.
The 28 May 2025 webinar was chaired by Development’s Executive Editor, Alex Eve and featured the first authors of the finalists of Development’s 2024 outstanding paper prize.

 (No Ratings Yet)
(No Ratings Yet)


 (2 votes)
(2 votes)