This week we’ll meet Dr Karin Van der Burg, a new faculty at Clemson University. Karin’s introduction to seasonal adaptation came during her undergrad first-year biology lecture, when she first heard about butterflies changing wing colors depending on temperature. “I thought that was SO COOL!” she recalls that moment. Her first encounter with phenotypic plasticity shifted how she saw development. Since then, Karin’s curiosity has led her deep into the physiological and genetic mechanisms that allow insects to tune their development to the seasons. From hormones and gene expression to chromatin and cold tolerance, her work spans multiple levels of biological organization. She’s studied butterflies that color-shift with the calendar, and budworms that shut down development to survive freezing winters. Now at her own lab, she’s combining genetics and physiology to explore how environmental cues like day length, temperature and seasons get integrated by hormonal systems to shape development using the Buckeye butterfly, Junonia coenia, and other incredible insect models. At heart, her science is driven by one thing: a need to know how it all works. Check out her lab page here. Give her a follow over Twitter and Bluesky. Keep an eye for announcements because she will soon be hiring postdocs and students in her lab !
What was your first introduction to the study of seasonal adaptation in insects? Tell us about that moment and how it shaped your scientific path?
I believe the first time I heard about this was in first year of college, in a lecture from Dr. Paul Brakefield. He told us about Bicyclus anynana, a small brown African butterfly species that changes its phenotype depending on rearing conditions. Exposure to cooler temperatures leads to a dull brown phenotype, which is adaptive in the dry season, while exposure to warmer temperatures results in the development of colorful wing eyespots, which are more adaptive in the wet season. At the time I thought that was SO COOL!
This was basically my first introduction to phenotypic plasticity: the phenomenon where multiple phenotypes could be created from one genotype. It really changed my perspective on how organisms develop and grow; I used to believe it was a very deterministic process, but instead it turned out to be very adaptable.
The Common Buckeye butterfly, Junonia coenia
Tell us what sparked your interest in the connection between endocrine signaling and seasonal adaptation? Walk us through your journey into studying the genetic and physiological basis of seasonal adaptation and introduce us to the field.
What really drew me in was the idea that phenotypes are plastic. At the time, work done in the evolution group at Leiden University did a lot of research in hormonal signaling underlying this plasticity, and I was fortunate enough to be able to do my undergraduate research project in that group. My project focused on genes associated with DNA methylation. It didn’t really go anywhere, but it was my first introduction to hormonal signaling and epigenetic signaling possibly working in concert.
As I continued my education, into my master’s and my PhD, I really started to think about how endocrine signaling affects epigenetic changes, which in turn results in changes in developmental pathways, resulting in different phenotypes. I realized to truly understand how organisms respond to seasonal changes, we need to look at the complete picture, and not one small aspect.
During my PhD I did mostly genetics research (with Dr. Reed at Cornell University), looking at genetic changes involved with changes in butterfly wing color plasticity. I also looked at changes to gene expression and chromatin accessibility, with a little bit of endocrine signaling, all to understand how seasonal plasticity in butterfly wing colors can evolve. While it was really interesting, I did feel like I was too focused on one aspect of a phenotype (in my case, butterfly wing color), and not the organism as a whole. I felt like there were many more changes involved in seasonal plasticity. Thus, for my postdoc I switched to a much more physiology focused lab (Dr. Marshall at UBC) to really get that more holistic perspective. To really understand how insects adapt to seasonal conditions, I believe we need to look at insect holistically, and not just one small aspect.
You have worked with different model systems including budworm Choristoneura fumiferana, and the common buckeye butterfly Junonia coenia. Tell us about your experience of working with these unique systems. What advantages do they offer in studying seasonal adaptations?
The biggest advantage is that these two organisms allow me to investigate two different types of seasonal adaptation; J. coenia has two flight seasons a year, an early and a late summer season. Each season has different conditions, such as temperature, rainfall, and food availability. We can use that to investigate how insects survive and reproduce in different conditions. Most notable about this butterfly is its change in wing coloration: butterflies emerge with a pale tan color when reared under warm, long-day conditions, and a dark red color when reared under cold, short-day conditions. Very likely there are other seasonal adaptations too, although they are not well known.
C. fumiferana, or the eastern spruce budworm, is really interesting in that it only has one lifecycle per year, and it is also stationary in the northern boreal forest. That means it needs to survive winter, with truly harsh conditions. Some very extreme changes in phenotype are necessary to survive under those conditions. My main interest is in the diapause phenotype, a prolonged period of arrested development during the early larval stages that allows for survival during harsh winters.
I like the combination of the two, because it allows me to study survival and reproduction in multiple different seasons, with very different survival strategies!
Tell us about your work on butterfly wing color plasticity. What were some of your key findings regarding the genetic factors controlling changes in wing color plasticity? Specifically, how does the upregulation of metabolites like trehalase contribute to the environmentally induced and genetically assimilated red phenotype?
During my PhD, I did a big research project on seasonal wing color plasticity in Junonia coenia. As I mentioned, in the wild this butterfly displays seasonal wing color plasticity, but we found we can easily manipulate this through artificial selection, such that plasticity was lost in only a few generations. We found three genes to be involved in this loss of plasticity, trehalase, herfst, and cortex. Later, my colleagues found that it probably wasn’t cortex regulating plasticity, but ivory, a long non-coding RNA. (Fandino et al., 2024; Livraghi et al., 2024; Tian et al., 2024).
Trehalase was indeed a very interesting find! The gene is upregulated under cold conditions in Junonia, and in other insects it is involved in cold-hardening. Trehalose (the sugar) can act as a cryoprotectant (prevents hemolymph freezing), and it is involved in metabolism.
We hypothesized that trehalase may be involved with the production of red pigments as well, because the ommochrome pigment that produces the red color contains a sugar molecule.
It’s really interesting to hypothesize on the multiple roles trehalase might play in seasonal plasticity. For example, it could be that the involvement in red pigmentation is a secondary effect; where trehalase was upregulated at first to manage cold conditions, and later was co-opted to produce red pigmentation as well! I will say that this is mostly speculation at this point, but it is definitely a research avenue worth pursuing.
Butterflies rely on endocrine cues to regulate metabolism and developmental timing. How does the ecdysone signaling pathway integrate environmental information to drive seasonal phenotypes?
These are really, really good questions! There is a lot of evidence that ecdysone signaling in butterflies is a universal regulator of seasonal plasticity (Bhardwaj et al., 2020). Given that ecdysone signaling is responsive to changes in day length, it is likely that circadian genes are involved, although that mechanism is not well worked out in insects.
Can small changes in endocrine signaling lead to tissue-specific adaptations without widespread disruptions. How does this level of regulation evolve, and what makes it so flexible?
Again, very good question. There are two things at play here: the seasonal responsiveness of ecdysone signaling, and the ability of seasonal plasticity to rapidly evolve. I believe overall development is very robust against predictable seasonal changes in ecdysone signaling. Tissue specific adaptations, such as wing color and wing shape in Junonia, or eyespot size in other nymphalid butterflies, can evolve rapidly to become more or less responsive to fluctuations in ecdysone. I suspect that outside of predictable seasonal changes, fluctuations in ecdysone signaling would be very problematic for normal development. I think (I may be wrong) the reason why plasticity can evolve so rapidly is because the seasonal responsiveness system (ecdysone signaling) is so robust and thus predictable, and so it makes for a reliable internal cue to adjust tissue specific developmental programs.
How does natural selection shape the regulation of hormonal pathways involved in adaptation? Are there evolutionary constraints that limit how endocrine signaling can be modified?
I think we know very little about the evolution of hormonal pathways itself! There are almost certainly many constraints on how endocrine signaling itself can evolve, because hormonal signaling is involved in so many different things. I believe that natural selection can readily act on downstream receivers of endocrine signals, but maybe not so much on the ecdysone signal itself. I’m happy to be proven wrong here though!
Speaking of your experimental approach – What have been the biggest challenges in studying endocrine regulation of seasonal adaptation? Are there specific experiments that were particularly difficult to execute? Did you have to deal with midnight timepoints or require an army of undergrads/ long hours etc.
Ecdysone measurements are a huge pain… Much harder than any of the genetic analysis I’ve done; ATAC-seq /RNA-seq / CRISPR/Cas9 were all a breeze compared to ecdysone measurements. It appears such a straightforward experiment, but I was constantly dealing with broken HPLC machines, degradation of ecdysone samples in the freezer… And yes, many late night or early morning sampling time points. I did all the sampling myself, but I did have an army of undergraduate students to help rear all the caterpillars!
Looking ahead, what are the next big questions in understanding how endocrine signaling and metabolism intersect with seasonal adaptation?
I’m very interested in exploring the interplay between circadian rhythms and ecdysone signaling! How/why is ecdysone signaling so responsive to external cues?
What role does curiosity play in your life, both within and outside of science? Why do you choose to work on insects?
Ultimately, I just want to know how stuff works… I enjoy thinking about biological research questions that integrate external and internal factors. I landed on gene regulatory mechanisms because for me, that’s where the bridge is between hard-coded DNA and the final phenotype which is very dependent on external conditions. I mostly work on insects because they are easier to work with than mammals, and because many insect species are very important to us.
Have you noticed a shift in how researchers approach insect biology?
Definitely have seen a shift to consider organisms more holistically, integrating factors at multiple levels of biological organization. I also feel like the scientific community is a lot more aware how important external conditions such as seasons are! So yes, I definitely think so.
Tell us about how you see the future of adaptative metabolism evolve with the new upcoming tools – what techniques have you used and which tools are you most excited about?
Single-cell RNA/ATAC-seq, and new CRISPR/Cas9 applications…
I haven’t worked with single cells yet, but I’d like to in the future! I’ve worked a lot with gene-editing through CRISPR/Cas9, but new techniques using CRISPR are coming out regularly and it is very exciting.
Were there any pivotal moments that shaped your career path? What’s an unexpected place you’ve found inspiration for your work? What advice would you offer to students and early-career scientists?
I was in a very genetics driven lab during my PhD, working with Bob Reed at Cornell. During my PostDoc, I shifted to a much more physiology-oriented lab, working with Katie Marshall at UBC. That shift really cemented that I wanted to integrate physiology and genetics research. I’m not sure this is ‘unexpected’ but I get most inspired from conversations with other folks, especially new students. The questions I get asked, especially ones that I don’t know the answer to, are really inspiring!
For advice, always be willing to consider new perspectives, and never be afraid to share your own thoughts, even if you’re not super confident. Science is a group effort!
How do you maintain a balance between your rigorous research activities and personal life? Are there hobbies or practices you find particularly rejuvenating?
These days I have a child, and that forces me to put down work when I’m at home! Otherwise, I love crafting! For example, making quilts or knitting.
If you hadn’t embarked on a career in biological research, what other profession might you have pursued, and why?
I truly don’t know – Before I found biology I tried engineering for a year and failed miserably. I feel like biology is the alternative career path for me and it has worked out well so far!
Anything you’d want to highlight for the future?
I’ll be looking for a postdoc this coming year! I’ll make a more formal announcement soon. I’m also rounding up a big project that I started in my postdoc on local adaptation in spruce budworm, I’m very excited for that!
Last week we learnt about how cancer cells rewire their metabolism to alter their cell fate and proliferate, check out –Switching Gears – Metabolic rewiring in cancer (Luis Cedeno-Rosario).
Spotted a preprint in this list that you love? If you’re keen to gain some science writing experience and be part of a friendly, diverse and international community, consider joining preLights and writing a preprint highlight article.
Hannah N. Gruner, C. J. Pickett, Jasmine Yimeng Bao, Richard Garcia, Akiko Hozumi, Tal Scully, Shaoyang Ning, Mavis Gao, Gia Bautista, Keren Maze, Karissa Lim, Tomohiro Osugi, Mae Collins-Doijode, Ofubofu Cairns, Gabriel Levis, Shu Yi Chen, TaiXi Gong, Honoo Satake, Allon Moshe-Klein, Eduardo D. Gigante, Yasunori Sasakura, Bradley Davidson
Alexandra E. Wehmeyer, Johanna K. Schmitt, Felix Eggersdorfer, Lea Zissel, Chiara M. Schröder, Mehmet Tekman, André Dias, Katrin M. Schüle, Simone Probst, Alfonso Martinez-Arias, Katie McDole, Sebastian J. Arnold
Wen Tian, Timothy Ting-Hsuan Wu, Shenbiao Gu, Jason L. Chang, Cerianne Huang, Ryan Vinh, Adam M. Andruska, Kyle K. Song, Dongeon Kim, Yu Zhu, Seunghee Lee, Junliang Pan, Peter N. Kao, Tushar Desai, Lawrence S. Prince, Lindsay D. Butcher, Xinguo Jiang, Marlene Rabinovitch, Kristy Red-Horse, Mark R. Nicolls
Alicia Lardennois, Chaitanya Dingare, Veronika Duda, Petra A Klemmt, Constanze Heinzen, David Kleinhans, Thorsten Falk, Carsten Schelmbauer, Olivia Mozolewska, Sofia Papadopoulou, Jason J K Lai, Didier Y R Stainier, Virginie Lecaudey
Chih-Wen Chu, Satheeja Velayudhan, Jakob H. Schauser, Sapna Krishnakumar, Stephanie Yang, Keiji Itoh, Dominique Alfandari, Ala Trusina, Sergei Y. Sokol
Soham Basu, Petrus Steenbergen, Florian Gabler, Alexandre Paix, Paolo Ronchi, Gleb Bourenkov, Thomas Schneider, Jonas Hellgoth, Anna Kreshuk, Suat Özbek, Aissam Ikmi
Konrad Chudzik, Isabel Guerreiro, Samy Kefalopoulou, Alex Abraham, Magdalena Schindler, Alessa Ringel, Mario Nicodemi, Irina Solovei, Andrea M. Chiariello, Stefan Mundlos, Jop Kind, Michael I. Robson
Alexandre Jourdon, Jessica Mariani, Abhiram Natu, Feinan Wu, Boxun Li, Davide Capauto, Kevin T. Hagy, Scott Norton, Livia Tomasini, Alexias Safi, Anahita Amiri, Jeremy Schreiner, Cindy Khanh Nguyen, Neal Nolan, Matthew P. Nelson, Daniel M. Ramos, Michael E. E. Ward, Anna Szekely, James C. McPartland, Kevin Pelphrey, Pamela Ventola, Katarzyna Chawarska, Charles A. Gersbach, Gregory E Crawford, Alexej Abyzov, Flora M. Vaccarino
Jason F. Cooper, Kim Nguyen, Darrick Gates, Emily Wolfrum, Colt Capan, Hyoungjoo Lee, Devia Williams, Chidozie Okoye, Kelsie Nauta, Ximena Sanchez-Avila, Ryan T. Kelly, Ryan Sheldon, Andrew P. Wojtovich, Nicholas O. Burton
Bernard K. van der Veer, Colin Custers, Wannes Brangers, Riet Cornelis, Spyridon Champeris Tsaniras, Kobe De Ridder, Bernard Thienpont, Huiyong Cheng, Qiuying Chen, Daniel Kraushaar, Richard H. Finnell, Steven S. Gross, Kian Peng Koh
Takashi Ishida, Juliane Mercoli, Adam M. Heck, Ian Phelps, Barbara Varnum-Finney, Stacey Dozono, Cynthia Nourigat-McKay, Katie Kraskouskas, Rachel Wellington, Olivia Waltner, Dana L Jackson, Colleen Delaney, Shahin Rafii, Irwin D. Bernstein, Kimberly A. Aldinger, Birth Defects Research Laboratory (BDRL), Cole Trapnell, Helong G. Zhao, Brandon Hadland
Cesi Deng (邓策思), Adedamola Elujoba-Bridenstine, Ai Tang Song, Rylie M. Ceplina, Casey J. Ostheimer, Molly C. Pellitteri Hahn, Cameron O. Scarlett, Owen J. Tamplin
Beverly A Rothermel, Malay Chaklader, Guilherme H Souza Bomfim, Neil Jeju, Yingli Duan, Brian F Niemeyer, Joaquín Maximiliano Espinosa, Edwin Rosado-Olivieri, Weichun Lin, Rodrigo S Lacruz
Akaljot Singh, Holly M. Poling, Jennifer Foulke-Abel, Nambirajan Sundaram, Abid A. Reza, Sarah Joseph, Abrahim ElSeht, Kalpana Srivasta, Maksym Krutko, Christopher N. Mayhew, David T. Breault, James M. Wells, Jay Thiagarajah, Amy E. O’Connell, Olga Kovbasnjuk, Michael A. Helmrath
Seungmi Ryu, Jason Inman, Hyenjong Hong, Vukasin M Jovanovic, Yeliz Gedik, Yogita Jethmalani, Inae Hur, Ty Voss, Justin Lack, Jack Collins, Pinar Ormanoglu, Anton Simeonov, Carlos A. Tristan, Ilyas Singeç
From Ryu et al. This image is made available for use under a CC0 license.
Anahí Binagui-Casas, Anna Granés, Alberto Ceccarelli, Eleni Karagianni, Daniel Lopez Ramajo, Rosa Portero, Matthew French, Jen Annoh, Frederick C.K. Wong, Sally Lowell, Osvaldo Chara, Valerie Wilson
Yiqian Li, Sean T.S. Law, Wenyan Nong, Wai Lok So, Yichun Xie, Thomas C.N. Leung, Tse Ho Li, Joyce Tse, Ho Yin Yip, Oli Jin, Jordan Zhang, Apple PY Chui, Kwok Fai Lau, Akbar John, Zhen-peng Kai, William G. Bendena, Alexander Hayward, Yingying Wei, Ting Fung Chan, Sai Ming Ngai, Jerome HL H
Ann K. Baako, Ragavi Vijayakumar, Daniel Medina-Cano, Zhaoquan Wang, Jesús Romero-Pichardo, Kelvin Fadojutimi, Stephanie C. Do, Yuan Lin, Mohammed Islam, Sanjana Dixit, Alissa J. Trzeciak, Justin S.A. Perry, Thomas Vierbuchen
Binayok Sharma, Xinyue Lu, Hamood Rehman, Vandré C. Figueiredo, Carol Davis, Holly Van Remmen, Shihuan Kuang, Susan V. Brooks, Krishna Rao Maddipati, James F. Markworth
Seda Akgün, Thomas Lenz, Annika Zink, Karina Stephanie Krings, Sebastian Wesselborg, María José Mendiburo, Alessandro Prigione, Kai Stühler, Björn Stork
Samuel G. Regalado, Chengxiang Qiu, Sanjay Kottapalli, Beth K. Martin, Wei Chen, Hanna Liao, Haedong Kim, Xiaoyi Li, Jean-Benoît Lalanne, Nobuhiko Hamazaki, Silvia Domcke, Junhong Choi, Jay Shendure
Cloe de Luxán-Hernández, Thomas J. Ammitsøe, Jakob V. Kanne, Sabrina Stanimirovic, Milena E. Roux, Zoe Weeks, Michael Schutzbier, Gerhard Dürnberger, Elisabeth Roitinger, Liechi Zhang, Oliver Spadiut, Masaki Ishikawa, Mitsuyasu Hasebe, Laura A. Moody, Yasin F. Dagdas, Eleazar Rodriguez, Morten Petersen
Edgar Demesa-Arevalo, Hannah Dorpholz, Isaia Vardanega, Jan Eric Maika, Itzel Pineda-Valentino, Stella Eggels, Tobias Lautwein, Karl Kohrer, Thorsten Schnurbusch, Maria von Korff, Bjorn Usadel, Rudiger Simon
Tasnim Zerin, Paola Ruiz-Duarte, Ann-Kathrin Schürholz, Theresa Schlamp, Yanfei Ma, Carlo Bevilacqua, Nabila El Arbi, Christian Wenzl, Andrej Miotk, Robert Prevedel, Thomas Greb, Jan Lohmann, Sebastian Wolf
Jie Fu, Brandon James, Madara Hetti-Arachchilage, Yingjie Lei, Brian McKinley, Evan Kurtz, Kerrie Barry, Stephen P. Moose, John E. Mullet, Kankshita Swaminathan, Amy Marshall-Colon
Callum V. Bucklow, Emanuell Duarte Ribeiro, Fabrizia Ronco, Nathan Vranken, Michael K. Oliver, Walter Salzburger, Melanie Stiassny, Roger Benson, Berta Verd
Allyson Caldwell, Liheng Yang, Rebecca L. Casazza, Rizban E. Worota, Cole McCutcheon, Patrick S. Creisher, Erika Zhan, Clara Reasoner, Ashley Higgins, Tony Schountz, Carolyn B. Coyne
Marie Lebel, Tiphaine Sancerini, Sharon Rabiteau, Solène Marchal, Pragati Sharma, Tal D. Scully, Estelle Balissat, Laurel S. Hiebert, Anthony W. De Tomaso, Allon M. Klein, Alexandre Alié, Stefano Tiozzo
Patrizia Pessina, Mika Nevo, Junchao Shi, Srikanth Kodali, Eduard Casas, Yingzhi Cui, Alicia L. Richards, Emily J. Park, Xi Chen, Florencia Levin-Ferreyra, Erica Stevenson, Nevan J. Krogan, Danielle L. Swaney, Qilong Ying, Qi Chen, Justin Brumbaugh, Bruno Di Stefano
Fangfei Jiang, Michael Boylan, Dale W. Maxwell, Wasay Mohiuddin Shaikh Qureshi, Charlie F. Rowlands, Gennadiy Tenin, Karen Mitchell, Louise A. Stephen, Elton J. R. Vasconcelos, Dapeng Wang, Tong Chen, Junzhe Zha, Jingshu Liu, Nouf Althali, Dragos V. Leordean, Meurig T. Gallagher, Basudha Basu, Katarzyna Szymanska, Advait Veeraghanta, Bernard Keavney, Martin J. Humphries, Jamie Ellingford, David Smith, Colin A. Johnson, Raymond T. O’Keefe, Sudipto Roy, Kathryn E. Hentges
Kristiina Uusi-Rauva, Anniina Pirttiniemi, Antti Hassinen, Ras Trokovic, Sanna Lehtonen, Jukka Kallijärvi, Markku Lehto, Vineta Fellman, Per-Henrik Groop
Atesh Kara Worthington, Beltran Borges, Tony Lum, Elisa Schrader Echeverri, Fareha Moulana Zada, Marco Cordero, Hyejin Kim, Ryan Zenhausern, Ozgenur Celik, Cindy Shaw, Paula Gutierrez-Martinez, Marzhana Omarova, Chris Blanchard, Sean Burns, Kyle Cromer, James Dahlman, Tippi MacKenzie
Quanyi Zhao, Albert Pedroza, Disha Sharma, Wenduo Gu, Alex Dalal, Chad Weldy, William Jackson, Daniel Yuhang Li, Yana Ryan, Trieu Nguyen, Rohan Shad, Brian T. Palmisano, João P. Monteiro, Matthew Worssam, Alexa Berezwitz, Meghana Iyer, Huitong Shi, Ramendra Kundu, Lasemahang Limbu, Juyong Brian Kim, Anshul Kundaje, Michael Fischbein, Robert Wirka, Thomas Quertermous, Paul Cheng
Annabelle Suter, Alison Graham, Jia Yi Kuah, Jason Crisologo, Chathuni Gunatilake, Koula Sourris, Michael See, Fernando J Rossello, Mirana Ramialison, Katerina Vlahos, Sara E Howden
Many times, the project or PhD is over, and the paper published, and yet the story unfinished. During the course of PhD, there arise many mysterious observations and unanswered questions.
My PhD work was published last year, where we describe the role of Cph (Chronophage) in locomotor behaviour of Drosophila. I report here an observation that isn’t completely explained by our present understanding of neural development in Drosophila.
Surprises on our path: When I started my PhD, with Prof. Upendra Nongthomba at the Indian Institute of Science, Bengaluru, this gene Cph was only a number (CG9650). Where to look for this unknown gene’s function? The project was a challenge, and it threw a couple of surprises in our path.
First, the only evidence that this gene might have a role in neural development (and hence our interest in it) came from a mis-expression screen. This screen showed that pan-neural expression of this gene i.e. in all neuroblasts and glia caused axon guidance defects. Was this due to its expression in neuroblasts/glia other than those in which it normally expressed? That’s what we thought – wrong cell, wrong time. The defects were grave enough to cause embryonic lethality.
However, upon knockdown of this gene in neuroblasts, or glia, or neurons, we didn’t see any axon guidance defect, or embryonic lethality. Knockdown in neurons resulted in flies that were constantly shaking, short-lived and failed to reproduce.
Normal fly movementMovement of Cph knockdown flies
The second challenge lay in understanding the behavioural defect. I first thought the constant shaking of these flies was due to seizures. I requested inputs from experts – the late Prof. K. S. Krishnan (National Centre for Biological Sciences, Bengaluru, India), and Prof. Dan Kuebler (Franciscan University of Steubenville, Ohio, USA). Prof. Krishnan asked me to take a video of higher magnification. (Unfortunately he passed away before we could reach a conclusion). Prof. Dan Kuebler, an expert in epilepsy in Drosophila, said the shaking didn’t look like seizures to him.
It occurred to me that the flies that lie on their back and keep shaking are perhaps trying to get back on their feet. Most of them were unable to stand up, and even if they did, they would fall down within seconds. So, the shaking was due to a motor defect, and not seizures. Electro physiological recordings also supported this – aberrant spontaneous spikes indicative of seizure were absent. (However, they did show bang sensitivity i.e. when subjected to a mechanical shock, they went into paralysis. This is mostly seen in seizure susceptible genotypes. So seizure susceptiblility cannot be ruled out in Cph knockdown animals.)
The baffling observation: We asked – at what stage of development was this gene necessary? Surprisingly, knockdown of this gene in adults didn’t have any effect! Knockdown during the larval stage reduced locomotor activity at that stage, but abolished locomotor activity in adults.
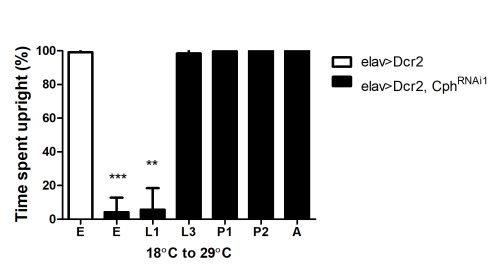
Effect of Cph knockdown at different stages of development: Cph was knocked down during different stages, and the resulting flies were assessed for their time spent upright.
Animals were initially reared at 18°C, and then shifted to 29°C at different stages of development. Shifting Cph knockdown animals at embryonic or L1 stage results in flies with locomotor defects. Shifting the animals post L3 stage results in flies with normal locomotor activity.
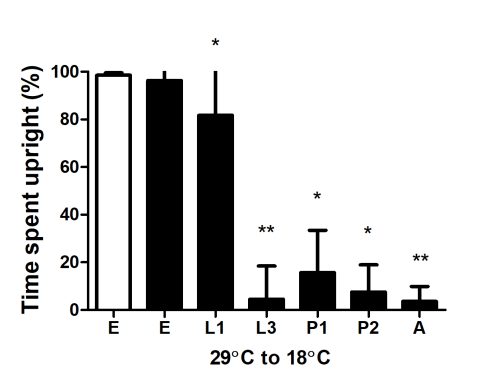
E, Embryo; L1, first larval instar; L3, third larval instar; P1, early pupa (less than 30hrs); P2, late pupa (70-90 hrs); A, adult
Animals were initially reared at 29°C and then shifted to 18°C at different stages of development. Shifting Cph knockdown animals at embryonic or L1 stage results in flies with normal locomotor activity. Shifting post L3 stage results in flies with locomotor defects.
How could this be explained? Were this protein expressed in larval neural stem cells, a possible explanation could be arrived at. But our observation was the result of knockdown in neurons! How could neuronal activity during development or the lack thereof affect their function in adults?
I outline two hypotheses:
Effect on re-specification of primary neurons.
Primary neurons are those born during the first phase of neuroblast divisions in the embryo. These neurons function during the larval stage and are reconfigured to connect to new targets to function during the adult stage. During neuronal remodelling that occurs during metamorphosis, neuronal processes are withdrawn from the larval targets and new axons and dendrites form towards adult specific targets. This re-specification of primary neurons could be affected if their functioning during the larval stage caused irreversible changes in them.
In case of motor neurons, activity dependent plastic changes during the larval stage could affect neuromuscular junction morphology, and this effect could persist during the remodelling stage.
Effect of energy state on formation/function of secondary neurons
Quiescent embryonic neuroblasts, post activation in first instar larvae, undergo another round of proliferation. This phase of neuroblast divisions produces the secondary neuronal lineages that function in the adult brain. Reactivation and subsequent division of these quiescent neuroblasts depends upon a signal from the fat body, which in turn depends upon the nutritional state of the animal (Chell and Brand 2010; Sousa-Nunes et al. 2011). Whether nutritional state also affects differentiation of neurons is not known.
Synapse function is an energy intensive process. ATP is required for axonal transport, vesicle fusion, neurotransmitter uptake etc. Animals with reduced PGK (PhosoGlycerateKynase), an enzyme required for ATP generation, show locomotor defects at larval and adult stage (Wang et al. 2004). PGK deficiency in humans has been shown to cause seizures, myopathy and muscle fatigue (Tsujino, et al. 1995), (DiMauro, Dalakas et al. 1983). The metamorphosis that follows the larval stage occurs using the energy stored during this stage. Metamorphosis is triggered by ecdysone release, which happens only after sufficient nutrients to survive metamorphosis have been acquired (Di Cara and King-Jones 2013). Though Cph knockdown animals probably acquire sufficient energy to survive metamorphosis, their neuronal development during this phase might be compromised.
I would welcome other ideas or suggestions.
References:
Chell, J. M. and A. H. Brand (2010). “Nutrition-responsive glia control exit of neural stem cells from quiescence.” Cell 143(7): 1161-1173.
Sousa-Nunes, R., L. L. Yee, et al. (2011). “Fat cells reactivate quiescent neuroblasts via TOR and glial insulin relays in Drosophila.” Nature 471(7339): 508-512.
Wang, P., S. Saraswati, et al. (2004). “A Drosophila temperature-sensitive seizure mutant in phosphoglycerate kinase disrupts ATP generation and alters synaptic function.” J Neurosci 24(19): 4518-4529.
Tsujino, S., S. Shanske, et al. (1995). “Molecular genetic heterogeneity of phosphoglycerate kinase (PGK) deficiency.” Muscle Nerve Suppl 3: S45-49.
DiMauro, S., M. Dalakas, et al. (1983). “Phosphoglycerate kinase deficiency: another cause of recurrent myoglobinuria.” Ann Neurol 13(1): 11-19.
Di Cara, F. and K. King-Jones (2013). “How clocks and hormones act in concert to control the timing of insect development.” Curr Top Dev Biol 105: 1-36.
The Center for Scientific Collaboration and Community Engagement (CSCCE) has announced its next offering of Scientific Community Engagement Fundamentals (CEF25F), which will run on Mondays and Thursdays beginning Thursday, 4 September until Thursday, 23 October.
The key dates are:
Social hour: Thursday, 4 September at 10am – 11am EDT / 2pm – 3pm UTC
Monday lessons: 8, 15, 22, 29 September; 6*, 20 October at 10am – 11:30am EDT / 2pm – 3:30pm UTC
Thursday Co-Labs: 11, 18, 25 September; 2, 9, 16 October at 10am – 11:30am EDT / 2pm – 3:30pm UTC
Graduation: 23 October at 10am – 12pm EDT / 2pm – 4pm UTC
*CSCCE will be closed on Monday, 13 October 2025 in observance of Indigenous Peoples’ Day. To accommodate this, we have slightly adjusted the schedule for this CEF cohort. “Reading Week” will now only take place during one session, on 2 October 2025, and the Week 7 lesson that would have taken place on Monday, 13 October will be held on Thursday, 9 October.
Scientific Community Engagement Fundamentals is an eight-week course designed to offer new or existing community managers core frameworks and vocabulary to describe their community’s purpose, refine or create strategic programming to engage community members around their shared goals, and identify ways to lower barriers to member participation. While the content is designed for any level of learner, it should not be thought of as a “beginner” course. Rather, it is intended to create common ground so that scientific community managers can converse across disciplines, more efficiently learn from one another, and build successful engagement strategies that are grounded in theory.
Each week, participants will meet virtually (using Zoom) for a 90 minute lesson and a 90 minute Co-Lab. While lessons will involve structured presentations and activities, Co-Lab time is for discussion, reporting out, and seeking feedback from instructors and fellow learners.
Each week will also include approximately 90 minutes of homework, for a total time commitment of 4.5 hours per week.
Pricing
Our pricing structure for CEF reflects the different organizations that community managers work for and the range of available budgets.
Discounted rate: Depending on participants paying the supporting rate, we may be able offer a limited number of discounted tickets for this cohort. If you would like to participate but your organization is unable to cover the whole cost, please complete this course discount request form.
Long range order and topological defects as tissue shapers
My PhD focused on studying the role of CDC42 isoforms during cell polarization and migration1. As CDC42 is a well-known regulator of actin polymerization, I developed extensive training in understanding actin regulation and dynamics in the context of cell polarization and migration.
Towards the final year of my PhD, during my time at the Institut Curie in Paris, I encountered the captivating field of biological active matter. There, I was introduced to the works of several renowned biophysicists working at the intersection of physics and biology. I became deeply intrigued by the emerging studies in this field, which is largely driven by biophysicists exploring long-range order in biological systems and investigating whether singularities in this order—known as topological defects—act as organizational centers that facilitate key biological processes.
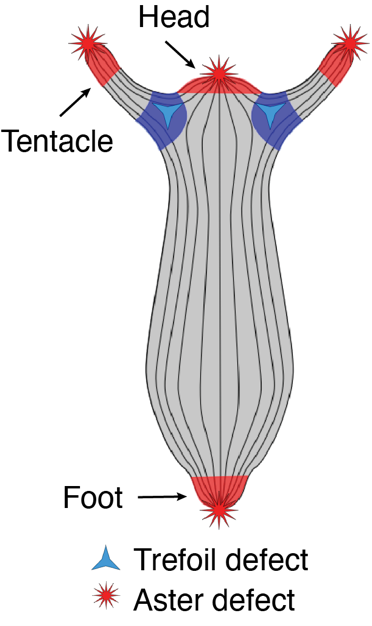
Figure 1 Skeleton schematic showing actin long-range order on the Hydra’s body plan
One striking example came from Benoit Ladoux, who demonstrated that cell extrusion events can occur at sites of topological defects in epithelial monolayers2. This was followed by two compelling preprints. The first, from the Roux Lab at UNIGE, showed that myoblasts confined to circular patches organize into nematic order (i.e., long-range, thread-like alignment). At aster-like topological defects, they observed cellular tornado-like 3D bulk cell extrusions, mimicking an in vitro reconstitution of muscle morphogenesis-like events3. The second preprint, from the Keren Lab in 2020, revealed that Hydra exhibits long-range nematic ordering in its supracellular actin organization, with topological defects correlating with head and foot morphogenesis4 (Fig. 1). Circling back to my PhD and my training in actin biology, seeing such single-cell-like highly organized actin structures in a tissue-scale regeneration in Hydra was fascinating and puzzling at the same time. I had so many agitating questions.
Beginning the postdoc: are topological defects shapers of in vivo morphogenesis?
I started in Aurélien Roux’s Roux Lab in September 2021—it was a rocky start, as I was still wrapping up my PhD manuscript and applying for postdoctoral fellowships, with deadlines fast approaching. I was also recovering from medical conditions that had worsened due to the sedentary lifestyle imposed on us during COVID.
In spite of these bottlenecks, I was genuinely excited to take on a new project to study the role of active matter in morphogenesis, especially in light of the recent breakthroughs in the field. Aurélien and I decided to explore the hypothesis of whether—and how—topological defects are required for shaping biological tissues, using various model organisms. We ventured into root morphogenesis in Arabidopsis (in collaboration with Luis Lopez Molina), slug morphogenesis in Dictyostelium (in collaboration with Thierry Soldati), and specifically, head regeneration in Hydra multi-headed mutants (in collaboration with Brigitte Galliot).
While I was juggling these different organisms and studying their fascinating morphogenetic events, I encountered a practical issue: my Hydra were constantly moving during imaging. I was performing these experiments with the help of Matthias Vogg, then a senior postdoc in the Galliot lab. To address the movement issue, I decided to image them using an agarose slab confinement method typically used for imaging plants—and Matthias agreed. Little did I know that this would compress the animal, leading to a mechanical induction of two-headed morphogenesis in Hydra.
Riding on this serendipitous discovery, I began following the phenomenon of mechanically induced morphogenesis in Hydra. During regeneration under compression, I tracked the emergence of new topological defects in the actin organization of the animal, which correlated with the formation of new heads. This observation confirmed the hypothesis proposed by the Keren lab, which suggested that aster topological defects are associated with new head formation during Hydra regeneration.
Topological defects shape animal tissues in a curvature dependent manner
Following this, Aurélien was thrilled and suggested I reach out to a theoretical physicist to explore the physical mechanisms behind how these additional topological defects could influence tissue shaping. He connected me with Daniel Pearce, an active matter theoretical physicist (then a postdoc in Karsten Kruse’s lab), who had already developed a mathematical model describing how long-range organization in Hydra could shape tissue5.
When I showed Daniel my findings, he was extremely excited and immediately came on board to help develop theoretical simulations of tissues under compression. Using his elastic nematic model, he described how +1 aster topological defects organize stresses and generate positive curvature (dome shapes), which is reminiscent of the shape of the Hydra head. In simulations under lateral compression, we observed that placing topological defects at the extremities, as seen in our regeneration experiments, led to the evolution of two dome-shaped structures.
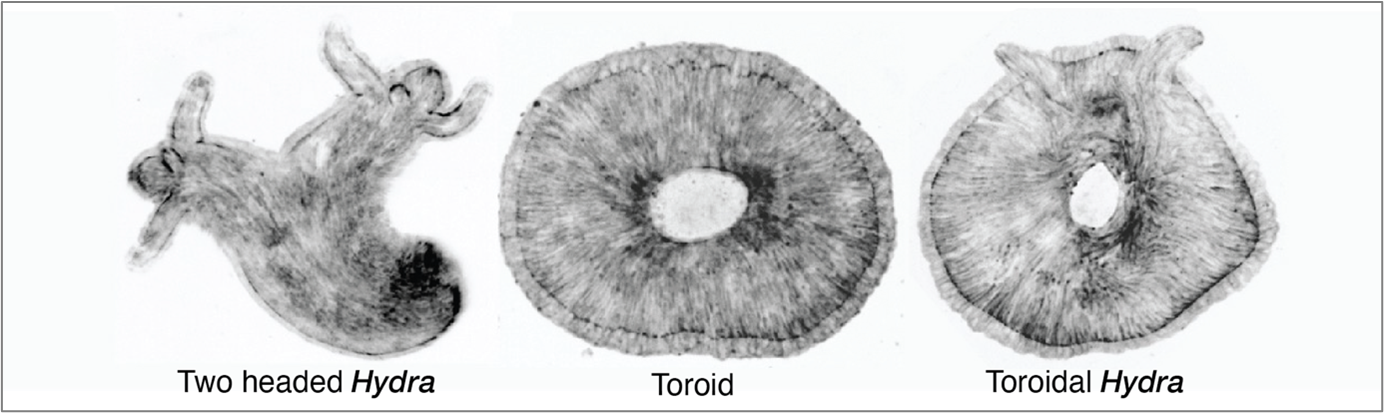
Furthermore, it was Dan who predicted that the orientation of tissue compression could dictate the fate of regeneration. The direction of compression influences how the long-range actin orientation experiences and responds to stress. As he suggested, we observed that when the tissue was oriented parallel to the compressive agarose slab, the sites of +1 aster defects buckled (inverted dome shapes) and underwent tearing, which then healed to form a defect-less toroid (Fig. 2).
This was a ground-breaking observation—it was the first time we observed the abolishment of body axis in an animal tissue that remained viable. I followed the defect-less torus over several days and observed that it failed to regenerate, as it maintained a perfectly symmetric actin configuration in which de novo +1 asters never arose. The tissue continuously attempted to regenerate a head but failed, due to the absence of a +1-aster topological defect.
Lastly, we all collectively thought however we should still be able to generate toroid with a +1 aster. By then we were also in touch with the Kerren lab and she invited me to Israel for a stay in her lab. Everyone suggested that theoretically a torus with defects could be generated. Then I hypothesized that if the compression occurred in a tissue with disordered actin to start with and a simultaneous tear occurred, then while the wound heals to create a 3D hole for the torus the disordered actin will order around it while generating a +1 aster required for a head. That is precisely what happened when we compressed the spheroid tissue that undergoes initial actin disorder and then ordering. This way we generated a torus with a +1 aster therefore a toroidal adult Hydra animal. This was really the cherry on top to see an animal with such a twisted muscle organization and topology.
In conclusion, these experiments established the need for +1 aster actin topological defects as head shapers in the animal. Their presence shapes the head and their absence give rise to no head formation, as observed in the defectless torus. The prospective research is to try to identify such long-range physical morphogens in other organisms, and strengthen the understanding of biological active matter.
Figure 2 Spinning disk microscopy images of fluorescent actin expressing (GFP-Ecto-LifeAct) Hydra at Day 6 of head regeneration. The three mechanically induced Hydra phenotypes, two-headed, actin-defectless toroid and toroidal Hydra. (2 votes) Loading...
Anyone—regardless of coding skills—should be able to generate a publication-quality plot of their data in minutes. That was the main motivation to develop a series of web apps to make state-of-the-art data visualization more accessible (huygens.science.uva.nl). But who cares, the same result can be achieved with generative AI (genAI) based tools, right?
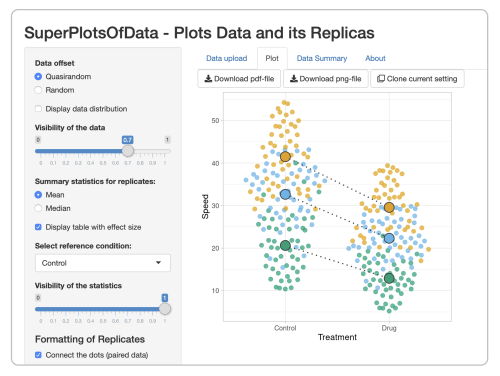
Before discussing what genAI can bring us for coding a plot, I briefly explain how the web apps work, so we can compare it with genAI later on. The data visualisation produced by the web app (the output) is coded in R and uses the {ggplot2} package. Another R-package, {shiny}, is used to create a graphical user interface (GUI). This GUI enables the user to optimize the data visualization by modifying the (invisible) code, through sliders, buttons, drop-down menus, and text fields. The process of creating a data visualization in a web app is highly interactive. By using a web app, the user can focus on what the data visualization should look like, without dealing with the code.
A screenshot of the web app SuperPlotsOfData. Users can optimize the data visualization with sliders, buttons, drop-down menus, and text fields.
By design, the web apps are somewhat limited in their options, so I started an online resource with dataViz protocols as well. My hope was that this resource would lower the barrier for people that wanted more control than what is possible in the web app and therefore would be motivated to learn R&ggplot2. But now, there is genAI. Coding can be done, rapidly and interactively, with websites that spit out code based on Large Language Models. Instead of focusing on the code and the technicalities that are required to build a data visualization, the user can focus on what the data visualization should look like (do you see the parallels with web apps?). This approach is aptly called vibe coding.
One of the prompts that was used in ChatGPT for vibe coding the data visualization of the output of a 96-wells plate that is shown below.
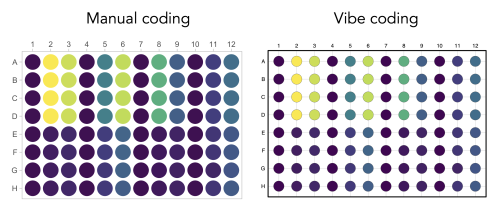
In a previous blog, I described that vibe coding “felt like I gained some kind of superpower”. But not everything is hunky-dory. It has been nicely documented by Mine Çetinkaya-Rundel that the AI-tool, besides the required changes, makes changes that are not explained and may be difficult to understand. I had exactly the same experience when I tried to vibe code a data valisualization that I had previously manually crafted (protocol 8 in the dataViz protocols book). In the end, the result (see below) is pretty neat, but it took several iterations (prompts), including some debugging of errors. I also noticed that understanding some of the basics (loading packages, knowing where to place the input data, how a plot is built using the {ggplot2} package) is needed to get the code to work. Worse yet, if the code seems to work but actually makes mistakes that are hard to spot things can go really wrong. For instance when doing some calculation for statistics that are difficult to understand or verify.
Graphical representation of readings from a 96-wells plate. The data visualization on the left was manually coded as detailed here and the data visualization on the right was generated by vibe coding in ChatGPT.
Are web apps still relevant when the same result can be obtained with vibe coding? Both the web apps and the genAI tools can be considered as a black box and allow the use to focus on the output. The genAI based tools offer great flexibility, but a strong point of the web apps is their predictable outcome, delivering a fully reproducible data visualization. The underlying code is available and the web apps are documented in (peer-reviewed) papers that can be cited. A practical advantage of web apps is that there is no need to install software or specific packages to run them. So I think there is still a future for the web apps. At the same time, I encourage experimenting with genAI as vibe coding offers new and exciting opportunities for data analysis and visualization. This will require at least a basic understanding of the coding language and sanity checks. Altogether, these are exciting times as the options for generating publication-quality data visualizations are expanding!
This week, we explore the story of Dr. Luis Cedeno-Rosario, a postdoctoral researcher in the Rutter Lab at the University of Utah. Luis’s path into metabolism began with a biochemistry class—an early glimpse into how cells adapt, survive, and respond to their world. His work explores how cancer cells alter their internal wiring to support unchecked growth and resist treatment—uncovering how shifts in metabolism can give tumors a survival advantage. These insights may help identify new ways to target cancer by exploiting its metabolic dependencies. Continue reading to learn how Luis is driven by curiosity, scientific precision, and how having a supportive mentoring environment impacted his journey. Check out his thoughts on how he winds science and music together, and how he views metabolism more than just chemistry— but as a language through which disease reveals its secrets and a window into how life adapts under pressure. Give him a follow over twitter and bluesky.
What’s your first memory of the field of metabolism? Could you share your journey into studying metabolism in disease contexts like cancer and cardiac disorders?
I have always been passionate about understanding how cells adapt to different environments and challenges, with a focus on cancer cell signaling and mitochondrial metabolism. I was taking a biochemistry and cell and molecular biology class as an undergraduate student at the University of Puerto Rico – Humacao and became fascinated by how multiple pathways intersect to regulate this process and their impact on cell behavior. I also had the great opportunity to do summer research internships at UT MD Anderson Cancer Center and at Johns Hopkins University which allowed me to learn more about the cell signaling and metabolism field. This is what led me to pursue a PhD in cell signaling in Dr. Deborah Chadee’s lab at the University of Toledo and a postdoc in metabolism in Dr. Jared Rutter’s lab at the University of Utah.
Introduce us to the field of cancer metabolism – you have worked on different types of cancer cells like ovarian cancer cells and liver cancer cells – tell us about your experiences.
During my first year of graduate school, I knew that I wanted to study cell signaling but I wasn’t sure in what context. I remember listening to Dr. Chadee’s talk in the signal transduction class and I was very fascinated by the complexity of the MAP Kinase signaling pathways and their role in ovarian cancer progression. Therefore, I decided to complete my PhD under the mentoring of Dr. Chadee where I worked on the regulation of the MAP3K MLK3 by CDK1 and CDK2 and their role in controlling cell division and proliferation in ovarian cancer cells (Check out the paper here). For my postdoc in the Rutter lab, I wanted to apply what I learnt during graduate school in the context of mitochondrial metabolism and their signaling pathways that are involved in liver cancer cell proliferation and progression.
How are different cells metabolically heterogeneous within the same tumor? Why is it important to study metabolic heterogeneity in cancer occurrence/progression – in term of both the cells themselves and the microenvironment?
Cells can have different metabolic profiles depending on the metabolites they need or are available in their surroundings. That heterogeneity can also come from where these cells are localized, for example, cells that are in a more hypoxic environment will probably have other metabolic needs than cells that are in a less hypoxic or normal environment. So cells have evolved in a way that they are very smart in choosing or taking what they need to meet their metabolic demands.
Tell us about your current work on metabolic signaling in the context of Wnt/beta-catenin pathway activation in liver cancer cells. How do you link it to the mitochondrial functioning and what future questions are you most excited about?
Our lab has done extensive work in characterizing the importance of the Mitochondrial Pyruvate Carrier (MPC) and its role in proliferation and tumorigenesis. I discovered that activation of beta-catenin represses MPC expression in liver cancer cells, and that this regulation rewires mitochondrial metabolism from glucose oxidation towards fatty acid oxidation. This is particularly interesting in the context of cancers in which MPC is downregulated and fatty acid oxidation is increased. I am very excited for the future since my findings opens up new avenues to explore ways to increase MPC expression in these tumors and increase the quality of life and survival of cancer patients.
Tell us how difficult some of these experiments are – do you have to deal with midnight timepoints or require an army of undergrads/ long hours, has to use some un-conventional/creative tools to overcome experimental challenges etc.
Some of these experiments have been truly a challenge and I have definitely spent many many hours in the lab trying to solve multiple research questions and/or developing new techniques to study the regulation of MPC by beta-catenin. I mentored an amazing summer research student, Nimo Abdi, who helped me a lot in the beginning of this project. I also have excellent collaborators, inside and outside the lab, who have contributed to the development of new ideas and have given me new perspectives on this regulation. I am very grateful to have them as collaborators and truly believe that these efforts will make a great impact in the metabolism field.
Building upon your work in the context of liver cancer metabolism, what are your upcoming plans? What metabolic pathways do you aim to investigate further to understand cancer progression from a cell growth and signaling perspective?
This switch in metabolic profile from glucose towards fatty acid oxidation is very exciting. So we are definitely looking more in depth at the metabolic processes that are changing and at the proteins and enzymes behind that regulation. One of the big questions we are investigating right now is to understand what fatty acids these cells prefer to utilize and their implication in liver cancer progression.
Before this, you studied cell signaling in ovarian cancer cells. How was the transition between fields, and what do you carry over from your previous research? Could you shed some light on your results regarding how MLK3 (Mixed lineage kinase 3) regulates cell cycle of ovarian cancer cells and tell us about your cool findings?
I have always thought about metabolism as another way of cells to sense their environment and metabolites are signaling molecules. These are multiple signaling pathways that are interconnected, and this concept was very similar to what I studied during graduate school. During my PhD, I found that the MAP3K MLK3 (Mixed lineage kinase 3) becomes phosphorylated by CDK1 and CDK2 to control ovarian cancer cell cycle progression. This research was published in the Journal of Biological Chemistry (JBC) doi: 10.1016/j.jbc.2022.102263 and I would encourage everyone to read it. It is a very interesting story that shows how these phosphorylation events act as “on” and “off” switches to control ovarian cancer cell division and proliferation.
How do you think scientific paradigms in the field of cancer metabolism will evolve in the coming decades – in regard to the new upcoming tools? Are we moving toward a more nuanced understanding, or do you see potential pitfalls?
I feel like we have bright future in our metabolism community. We have seen the development of great techniques such as mass spectrometry integrated with equilibrium dialysis for the discovery of allostery systematically (MIDAS) that was developed in our lab to identify novel interactions between metabolites and proteins. This is a fast-growing field and we are opening more doors to understand the complexity of metabolic pathways in multiple contexts, including cancer, cardiac function and neurodegenerative diseases, and in development. I am very excited for our future findings and hope that I can contribute significantly and have a positive impact not only in the research field itself but also in training the next generation of scientists.
What role does curiosity play in your life, both within and outside of science?
I believe that curiosity plays an important role in my scientific career. Understanding what is happening at the cellular level is pivotal in the development of new therapies, and that is what drives my passion for science. I want to be able to use my knowledge from cell and molecular mechanisms to develop new and better ways to treat multiple diseases or to at least increase the quality of life of the people affected by a particular disease.
What are the future research questions you are most excited to ask?
I am very excited about pursing metabolism in the context of cell biology and development. I think it is a field that is also growing very fast and I would like to contribute to it and make new discoveries.
Were there any pivotal moments that shaped your career path —and how have you found ways to build a supportive community in science?
I think that the most pivotal moment in my career path was creating a strong and supportive network of mentors within the cell metabolism and mitochondrial biology field. I have met many of these mentors in conferences and through the Burroughs Wellcome Fund Postdoctoral Diversity Enrichment Program (PDEP) that have been critical in my development as a future independent scientist. I am also very grateful to be part of the biochemistry department at the University of Utah and to receive a lot of internal support as a postdoctoral fellow.
How do you maintain a balance between your rigorous research activities and personal life? Are there hobbies or practices you find particularly rejuvenating?
Music! I have extensive training in classical music and I am actually a member of the Utah Medical Orchestra (UMO) where I play the flute and the piccolo. Music is definitely a big part of who I am.
If you hadn’t embarked on a career in biological research, what other profession might you have pursued, and why?
I would have pursued a law degree or a music degree in flute performance. In the law aspect, I like the complexity of finding new solutions to diverse problems. In the music aspect, I like how we can create art using a universal language and enjoy that art as a whole. Music can bring you different feelings and helps us express ourselves.
Last week we learnt about how viruses rewire and utilize host lipid metabolism using mosquitoes as a host model system with Wolbachia and dengue as viral players, check out the article – Lipids and Labyrinths(Cassandra Koh). Cassandra is a new PI, studying metabolic interactions of symbiosis and virus-virus host interactions. She is seeking motivated students and collaborators.
Madalena M. Reimão Pinto (Schier lab, University of Basel, Switzerland) and Sebastian Castillo Hair (Seelig lab, Washington University, Seattle, USA) joined forces to understand how zebrafish embryos orchestrate protein synthesis during early development. Through this collaboration, their recently published study in Developmental Cell identifies features in mRNA 5′ UTRs that act during early zebrafish development to regulate translation.
Madalena, what brought you to Alex Schier’s lab and how did you embark on this project?
During my PhD at the Vienna BioCenter, I worked on understanding the molecular mechanisms regulating mRNA biogenesis and function in the fruit fly. It so happened that the exonucleolytic enzyme that I was working on at the time had a striking effect on spermatogenesis, and the data pointed to defects resulting from misregulated mRNA translation. By the end of my PhD, I was fascinated by mRNA translational control in the context of organismal development. I quickly realized that we lacked a comprehensive understanding of how mRNA translation initiation – which is rate limiting for protein production – is regulated during the very fast-paced and temporally coordinated stages of early embryogenesis. This motivated me to tackle this question in the context of early vertebrate development. I also realized that my background in RNA biochemistry gave me an edge to start studying developmental biology at a mechanistic level from a different perspective. I chose to address this question using the zebrafish model because it allows me to combine high-throughput approaches with biochemistry and powerful genetic tools in live embryos, as they develop.
I had heard great things about the Schier lab from Andi Pauli (Schier lab Alumna, group leader at the Vienna BioCenter, Austria) and was inspired by Michal Rabani’s work on mRNA stability during zebrafish embryogenesis (Schier lab Alumna, group leader at the Hebrew University of Jerusalem, Israel). On one hand, I was looking to join a lab which had in depth knowledge about developmental biology and zebrafish genetics; on the other hand, I wanted to be part of a multidisciplinary and vibrant group of people who aspired to become group leaders, so the Schier lab was the perfect choice.
The main question motivating my postdoctoral work is: how do embryos know how much protein to make and when? To start dissecting this question systematically, I decided to focus on the 5′ UTR sequence, which is crucial for regulating translation initiation, and developed an approach to interrogate at transcriptome scale how the 5′ UTR contributes to regulating translational dynamics as embryos develop.
Madalena holding a tank of zebrafish. Photo credit: Annette Roulier, Biozentrum Basel.
How did the collaboration with the Seelig lab start?
Well, it took more than two years to get to a stage when I had designed and generated the 5′ UTR massively parallel reporter assay (MPRA) library, validated it and acquired the in vivo data. To be honest, I also spent quite some time running data analyses before convincing myself that the assay had actually worked and was recovering biologically meaningful information! Once I realized that was the case, I was super excited to try and learn as much from the data as possible. I was familiar with Georg Seelig’s work and had actually met him in person when he gave a talk at the Biozentrum’s Discovery Seminar. At that time, I was still running experiments but I approached Georg and asked if he would be interested in collaborating to explore the data with deep-learning models. Georg was excited about my project and told me to reach out when the right time came, and so I did! He then paired me up with Sebastian, and we started a wonderful collaboration together. Honestly, it was so efficient and so much fun: we would meet every two weeks, and at each meeting discuss our progress, come to an agreement on next analyses and experiments and then execute them. It gave the project a really nice momentum and it was just great to feel part of a team working towards a common goal. At the same time, Georg joined the Schier lab for a sabbatical, so it was great to have the opportunity to discuss and get feedback on a regular basis from both PIs in person. The project would have not been the same without Sebastian’s contributions, who developed the Danio Optimus 5-Prime (DaniO5P) deep-learning model to evaluate and interpret the 5′ UTR MPRA data. And Georg was the one who came up with the model’s name!
What were for you the most exciting findings, or particular moments during the project that stuck with you?
The most exciting moment was running the Kozak sequence and uORF analyses and realizing that the assay had actually technically worked. It also meant that the data likely contained additional sequence information to be uncovered, which was a super exciting prospect. And then of course, seeing motifs emerging from the motif enrichment analysis and the DaniO5P model!
I’d also like to say that when I started this project, I had no experience performing polysome profiling. I was extremely fortunate to meet Sunil Shetty, at the time a postdoc in the Hall group at the Biozentrum (now leading his independent research group at the Tata Memorial Centre in Mumbai, India), who taught me how to prepare sucrose gradients and perform polysome profiling. His happiness and positivity are contagious, and some of the most fun moments of my postdoc were spent next to the polysome profiling machine learning from Sunil. I am deeply grateful to him, and also to Michael Hall for kindly welcoming me in his lab.
And what was most challenging?
For sure it was starting my postdoc and having the COVID-19 pandemic hit a few months later. At the time, my plan was to experimentally define zebrafish 5′ UTRs by long-read sequencing to design the MPRA. There was so much uncertainty about when we could be back in the lab full time, that I decided to instead use an alternative approach to computationally infer 5′ UTR sequences from a public dataset of cap analysis of gene expression data. I don’t have a background in computational biology, so it was definitely a slow and iterative learning process – but also very rewarding in the end, and most importantly it allowed me to move forward with the project.
What’s ahead for you?
I am currently applying for Group Leader positions. Going on interviews and getting to meet faculty members from different research institutions is really an amazing and rewarding experience, but also very time-consuming, so it’s hard to get any experiments done alongside applications and interviews. I am super excited to start my own independent research group soon, and I hope to continue to collaborate with Sebastian on future projects!
Montage of zebrafish embryo images acquired at consecutive stages of development (from 2 to 10 hours post-fertilization) that were injected at the 1-cell stage with a fluorescent dextran dye (magenta) and an mRNA reporter encoding GFP (green), reminiscing of the moon phases. (1 votes) Loading...
At the end of each month, I pick the same month from a random year from the past 15 years of the Node, and take a look at what people were talking about back then.
Previously, I travelled back to February 2011, March 2013 and April 2014 to have a look around the Node. Luckily, I didn’t get lost along the timeline and managed to get back to the present day. But now I’m itching for another adventure. So this time, let’s fasten our seat belts and turn the dial to May 2016…
In this series, we asked prominent researchers to recommend their favourite hidden gems from history – papers that are, for whatever reason, unjustly overlooked today. Read the other Forgotten classics posts.
We love it when you’ve organised a science outreach activity and then tell us how it went. It’s also useful for other researchers to get inspiration about their own outreach events. Check out other ‘outreach’-related posts.
Catarina Vicente was the Node’s Community Manager from 2013 – 2016. What’s she been up to since then? Find out from this conversation with all the past and present Community Managers.
It lives. It lives! What lives, you may ask? Well, somewhere in a lab at Yale University, one young scientist has stuck human brain cells and chimp brain cells together to make a chunk of hybrid brain. A few weeks ago, I met with her to ask more about this research. She admits that it all sounds like “mad science,” but this mad scientist might be taking a big step forward on our path to find out what really makes us human.
Chimpanzees are our closest relatives, and closer than most of us would probably like to think about. We share some 98.8% of our DNA with chimps.1 This means only about 1.2% of our DNA accounts for the uncanny power of our species to build cities, write symphonies, split atoms, and do all the other things we alone do so well. We know that much of this uncanny power resides in our brain, which is massive compared to a chimp’s brain,2 and has a much larger wrinkled region at its front3 that does most of our complex “higher” thinking. This wrinkled region at the front of our brains takes almost twice as long to finish developing in us as it does in chimps,4 and scientists have long thought that its slow development in humans helps to explain our subtle and adaptable “higher” thinking abilities.4 What we do not know is how that meager 1.2% of our DNA goes about making our wrinkly-fronted brains develop so slowly.
For the last two years, a young scientist named Reem Abu-Shamma has been trying to change that. Since graduating summa cum laude from UCLA, Reem has made a career of mutating genes, creating artificial 3D clusters of human intestinal cells (delightfully called “organoids”), and using computer programs to study vast amounts of DNA. These endeavors might sound eerily sci-fi, but have in fact taught us a lot about public health and disease. Her work mutating genes in parasites could shed light, down the line, on how we treat some particularly nasty strands of malaria,5 and her work with human intestinal organoids promises to tell us more about the cellular basis for inflammatory bowel disease. Now, as a PhD student at Yale University, Reem has set her sights on what makes our brains human.
” Slower development means more time to make a big brain…So where in the genetic code is it telling our brains to develop slower? “
To investigate what makes our brains unique, Reem has created something like a hybrid “half-human, half-chimp brain.” This phrase baffled me as much as it’s probably baffling you, so I sat down with Reem to ask her more about what inspired this research. And, as I listened to Reem’s enthusiastic, down-to-Earth explanation for her project, it began to seem less like mad science and more like vital research. “Large brains allowed us to dominate the world for better or for worse,” Reem explains. She wants to find “the underlying code in our cells that has allowed us to do that.” In searching for this code, Reem has focused on the speed at which that wrinkly portion at the front of our brains develops. “Slower development means more time to make a big brain…and we know that the human brain takes a really long time to develop.” This slow pace of human brain development manifests at the cellular level6—individual human brain cells take years to branch out and mature, whereas those of chimps develop much faster. Reem’s research question is simple, then: “Where in the genetic code is it telling our brain cells to develop slower?”
To answer this question, Reem recently joined the Noonan Lab at Yale University, which has a long history of using the best-available gene-editing technology to study human brains. One particular focus of the Noonan Lab has been to find particular bits of DNA that distinguish humans from chimps and other animals. What exactly are these bits of human specific DNA? Well, as Reem explains, “they’re parts of the genome that are not genes,” but “dials” for genes, which make various brain-building genes more or less active as the brain develops.7 These bits of DNA are part of that 1.2% of our genetic code separating us from chimpanzees, and could tell us a lot about how that huge wrinkly portion at the front of our brains develops so slowly, gets so big and complex,5 and makes us so clever. Each one of these bits, once found, “gives us a hint that maybe this part of the genome helped us evolve big brains.” However, that hint alone doesn’t prove the bit’s role in making our brains bigger or tell us how it did. In order to actually verify what these human bits of DNA do, scientists have to mutate them, and see how those mutations affect the development and interaction of different human and chimp brain cells over time. Obviously, no one at any credible research institution wants to mess with the brain of an actual living human—institutional and federal guidelines fortunately forbid that kind of work. But scientists dowant to understand what these human-specific bits of DNA are doing. So how do you mutate realistic human brains without using actual real human brains?
Well, remember those “organoids” I mentioned before? Reem uses those. And they’re a lot less scary than they sound. “We’re not using real animals, or growing real brains” Reem assures me with a laugh. Instead, she’s using what amounts to just a few cells: To create brain “organoids”—again, 3D clusters of living brain cells—she uses cells that other labs have collected from human or chimp skin. These labs treated those skin cells with various molecules to “reprogram” them into stem cells, which can turn into almost any other kind of cell if given the right molecular cues. “In our case,” Reem explains, “we make them turn into neurons.”8 Reem uses this approach to create human and chimp brain cells, and then grows each reprogrammed brain cell into a different 3D cell cluster or “organoid.” The result in each case is a separate ball of brain cells9 for each species that develops much like they would in a real brain.
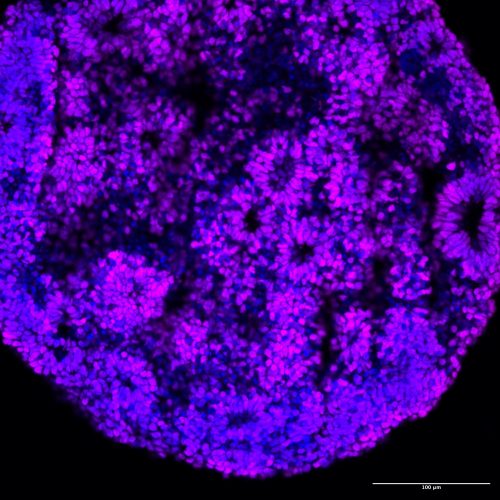
A human brain organoid, 30 days old, made by combining two different human cell lines. Cells are labeled with two overlain molecular markers——a blue one marking all cell nuclei and a violet one that marks “forebrain cortical neuron progenitors” (the kind of cells that end up forming the wrinkly front of our brains). The cells in this organoid have spontaneously arranged themselves into “rosettes”, much like brain cells do in an embryonic brain. Image courtesy of Reem Abu-Shamma.
With each brain organoid, Reem plans to test what our human-specific bits of DNA are doing to make our brains grow slower and larger. She will do this by tweaking or changing10 various human-specific bits of DNA to make them act more like the corresponding regions of chimp DNA, and vice versa. Then she’ll see how these modifications affect the activity levels of various brain cell genes and the “speed” at which those brain cells ultimately develop. “By ‘speed’, we don’t mean absolute time; rather, we have the technology to look at a single cell and figure out how mature it is based on the molecules we observe in it,” Reem clarifies. Then, for each bit of human-specific DNA, she’ll see whether the humanized chimp cells appear to develop more slowly, while the “chimpanized” human cells develop more quickly. Then we would know that this specific bit of our 1.2% unique genetic code is partly responsible for making our brains so weirdly human.
Reem finds the sheer size of this mystery fascinating. “The genome is a really big place,” she explains. “It’s so vast and we don’t know what most of it does. It kind of feels like detective work, because you’re trying to see where in this really big space it’s telling us to be human.” By tweaking little bits of human and chimp DNA so they behave more like their counterparts—a sort of genetic Freaky Friday—Reem can do just that, finding which bits of human-specific DNA tell our brain cells to grow in a human way. This in itself is the stuff of science fiction. However, Reem and her PhD advisor, Dr. James Noonan, are taking this approach one step further.
They aren’t just growing human brain-cell colonies and chimp brain-cell colonies. They’re mixing them together, to make something like a miniature hybrid brain. Despite their different origins, these cells branch out and interconnect much like the cells in our own brains, possibly creating a cellular communication network unseen in nature. “Why would you make a half-human, half-chimp brain?” Reem jokes that her mother and even her colleagues have often asked her this question. But Dr. Noonan initially suggested this approach, and Reem has pursued it, because we can learn a lot from it.
” It kind of feels like detective work, because you’re trying to see where in this really big space it’s telling us to be human. “
Brain cells don’t usually grow on their own. They grow in response to cues from neighboring cells, and these hybrid brains can show us the extent to which human brain cell development is genetically encoded. How much of how our brain cells behave is written in their DNA, and how much is determined by interaction with their cellular neighbors? Specifically, Reem is curious whether the sum of brain cell interactions, and the presence of similar brain cells from other species, together affect how fast that wrinkly portion at the front of the brain develops. Previous studies have found that these external cues (the “cellular environment”) don’t matter much for the development speed of human brain cells.11,12 However, few if any studies have used hybrid chimp-human brain organoids to study that big wrinkly-fronted part of the human brain. By creating hybrid chimp-human organoids with this specific type of brain cell, Reem will finally test whether environmental cues help it grow slower in humans. Reem gives me an example to help me wrap my head around this. So, suppose you “take a human cell and transplant it into a chimp brain organoid,” Reem explains. And then suppose you collect molecular data from human brain cells inside a purely human organoid and then do the same to human brain cells inside a human-chimp hybrid organoid. “If they’re exactly the same, then the environment the cell is in isn’t as important!”
Making and mutating hybrid brains is intense work. To do it right, Reem has to set up hundreds of different brain organoids, each in its own plastic well, and from a variety of different human and chimp donors. She goes into the lab every day. “I check on my cells immediately…first thing. I make sure they’re still alive.” She recounts instances where some of her organoids became cancerous, and others spontaneously collapsed and started dying—both unplanned events that threatened to skew her work and required hours of manual labor to remedy. “So many things can go wrong…it’s a lot of manual labor to make sure they’re alive and happy.” On a daily or weekly basis, she has to feed her many hundreds of brain organoids, and look at each one under a powerful microscope to make sure nothing has gone horribly wrong. She has to modify them all at just the right time, in just the right way. And then, when all of that is done, within the next few months, she’ll have to extract specific molecules from these organoids and analyze the resulting vast amounts of data to see how her mutations changed the approximate speed of brain cell development in her human brain, chimp brain, and hybrid brain organoids.
Reem is eager to find “what inferences we can make about the speed of development using these models”—at what rate the brain cells are likely growing, dividing, branching out, and developing their various special functions. With this approach, Reem wants to pinpoint some of the intrinsic genetic factors responsible for speeding up and slowing down the “molecular rate” of human brain cell development.
A hybrid human-chimp brain organoid from Reem’s first experiment on this project. Molecular markers tag chimp brain cells red and human ones green. This hybrid chunk of brain is 17 days old. Image courtesy of Reem Abu-Shamma.
When I asked Reem about the benefits of this research, her answer surprised me. Of course, her work does have implications for treating and understanding psychiatric or developmental conditions—autism spectrum disorder, schizophrenia, and other cognitive differences that often relate to brain development. That was the answer I expected. But Reem went on to highlight something else. “This is a very exploratory study,” she explained. “It’s hypothesis-generating,” and “in the history of science, doing fundamental research can sometimes lead you down unexpected paths, just because you’re exploring your curiosity.” This “fundamental research” is done not for its direct societal benefits, but to better understand ourselves and our world, and often has unexpected humanitarian value. For example, Reem points out, CRISPR was discovered by fundamental research projects on a few seemingly random repetitive patterns in microbe DNA. And yet, CRISPR now forms the most promising avenue for therapeutic gene editing and has a variety of other applications for human health and disease worldwide.13,14
Reem’s work on hybrid brains is fundamental research in the same way. Yes, it has biomedical implications. But its potential value is so much broader. It can shed light on the parts of our genetic code that separate us from chimps and other animals. As Dr. Noonan told her when he first suggested making human-chimp hybrids, “no one’s done it before,” and we can hardly begin to predict what it might tell us about what makes us human.
Caleb Gordon is a Postdoctoral Associate at Yale University, where he studies the evolution of reptiles during the time of the dinosaurs.Check out his website to follow his research and popular science writing.
Note from the author: This piece was written as part of a workshop series taught by Carl Zimmer, and organized by Yale’s Graduate Writing Lab, on science reporting intended for a general audience.This workshop challenged us to write a popular science article without any scientific jargon. However, for any scientists missing this jargon, I’ve included more scientific terminology in the References Cited below. This article benefited greatly from feedback by Lauren Gonzalez and Joseph Lee at the Graduate Writing Lab.
[2] For more information about the evolution of human brain size, you can check out this research paper: Smaers, J. B., R. S. Rothman, D. R. Hudson, A. M. Balanoff, B. Beatty, D. K. N. Dechmann, D. De Vries, et al. “The Evolution of Mammalian Brain Size.” Science Advances 7, no. 18 (April 30, 2021): eabe2101. https://doi.org/10.1126/sciadv.abe2101.
[3] This wrinkled region at the front of the brain is called the “prefrontal cortex.” For more information about this remarkable brain region and its implications for our higher executive functioning abilities, you can check out this research paper: Preuss, Todd M., and Steven P. Wise. “Evolution of Prefrontal Cortex.” Neuropsychopharmacology 47, no. 1 (January 2022): 3–19. https://doi.org/10.1038/s41386-021-01076-5.
[4] Additional information on the prefrontal cortex is provided in this research paper from the same journal: Kolk, Sharon M., and Pasko Rakic. “Development of Prefrontal Cortex.” Neuropsychopharmacology 47, no. 1 (January 2022): 41–57. https://doi.org/10.1038/s41386-021-01137-9.
[5] The following research paper summarizes the results from Reem’s collaborative gene-editing work with malarial disease vectors: Subudhi, Amit Kumar, Anne-Lise Bienvenu, Guillaume Bonnot, Reem Abu-Shamma, Faryal Khamis, Hussain Ali Abdulhussain Al Lawati, Stephane Picot, Eskild Petersen, and Arnab Pain. “The First Case of Artemisinin Treatment Failure of Plasmodium Falciparum Imported to Oman from Tanzania.” Journal of Travel Medicine 30, no. 3 (May 18, 2023): taac092. https://doi.org/10.1093/jtm/taac092.
[6] The human brain matures more slowly in part because individual human brain cells take longer to develop. For a great review highlighting the uniquely protracted nature of human brain cell development, check out this recent paper: Lindhout, Feline W., Fenna M. Krienen, Katherine S. Pollard, and Madeline A. Lancaster. “A molecular and cellular perspective on human brain evolution and tempo.” Nature 630 (19 June 2024): 596–608. https://doi.org/10.1038/s41586-024-07521-x.
[7] For more information on what these human-specific bits of DNA are and what they do, check out this recent paper from the Noonan Lab: Pal, Atreyo, Mark A. Noble, Matheo Morales, Richik Pal, Marybeth Baumgartner, Je Won Yang, Kristina M. Yim, Severin Uebbing, and James P. Noonan. “Resolving the Three-Dimensional Interactome of Human Accelerated Regions during Human and Chimpanzee Neurodevelopment.” Cell 188, no. 6 (March 2025): 1504-1523.e27. https://doi.org/10.1016/j.cell.2025.01.007.
[8] For additional information about how these scientists create brain cells from stem cells, check out the following paper: Mariani, Jessica, Maria Vittoria Simonini, Dean Palejev, Livia Tomasini, Gianfilippo Coppola, Anna M. Szekely, Tamas L. Horvath, and Flora M. Vaccarino. “Modeling Human Cortical Development in Vitro Using Induced Pluripotent Stem Cells.” Proceedings of the National Academy of Sciences 109, no. 31 (July 31, 2012): 12770–75. https://doi.org/10.1073/pnas.1202944109.
[9] These colonies of brain cells are called “cortical organoids.” For more information about these remarkable 3D brain cultures, check out the following paper: Pollen, Alex A., Aparna Bhaduri, Madeline G. Andrews, Tomasz J. Nowakowski, Olivia S. Meyerson, Mohammed A. Mostajo-Radji, Elizabeth Di Lullo, et al. “Establishing Cerebral Organoids as Models of Human-Specific Brain Evolution.” Cell 176, no. 4 (February 2019): 743-756.e17. https://doi.org/10.1016/j.cell.2019.01.017.
[10] Reem mutates brain cell colonies using “arrayed CRISPR screens,” which are described in more detail in the following research paper: Bock, Christoph, Paul Datlinger, Florence Chardon, Matthew A. Coelho, Matthew B. Dong, Keith A. Lawson, Tian Lu, et al. “High-Content CRISPR Screening.” Nature Reviews Methods Primers 2, no. 1 (February 10, 2022): 8. https://doi.org/10.1038/s43586-021-00093-4.
[11] This research paper studied pure human and pure chimp brain cell organoids: Otani, Tomoki, Maria C. Marchetto, Fred H. Gage, Benjamin D. Simons, and Frederick J. Livesey. “2D and 3D Stem Cell Models of Primate Cortical Development Identify Species-Specific Differences in Progenitor Behavior Contributing to Brain Size.” Cell Stem Cell 18 (April 7, 2016): 467–480. http://dx.doi.org/10.1016/j.stem.2016.03.003.
[12] This research paper took human brain cells from that wrinkly region at the front of the brain and transplanted them into a live mouse brain: Linaro, Daniele, Ben Vermaercke, Ryohei Iwata, Arjun Ramaswamy, Baptise Libé-Philippot, Leila Boubakar, Brittany A. Davis, Keimpe Wierda, Kristofer Davie, Suresh Poovathingal, Pier-Andrée Penttila, Angéline Bilheu, Lore De Bruyne, David Gall, Karl-Klaus Conzelmann, and Vincent Bonin. Neuron 104 (December 4, 2019): 972–986. https://doi.org/10.1016/j.neuron.2019.10.002.
[13] Doudna, Jennifer A., and Emmanuelle Charpentier. “The New Frontier of Genome Engineering with CRISPR-Cas9.” Science 346, no. 6213 (November 28, 2014): 1258096. https://doi.org/10.1126/science.1258096.
[14] Barrangou, Rodolphe, and Jennifer A Doudna. “Applications of CRISPR Technologies in Research and Beyond.” Nature Biotechnology 34, no. 9 (September 2016): 933–941. https://doi.org/10.1038/nbt.3659.


 (2 votes)
(2 votes)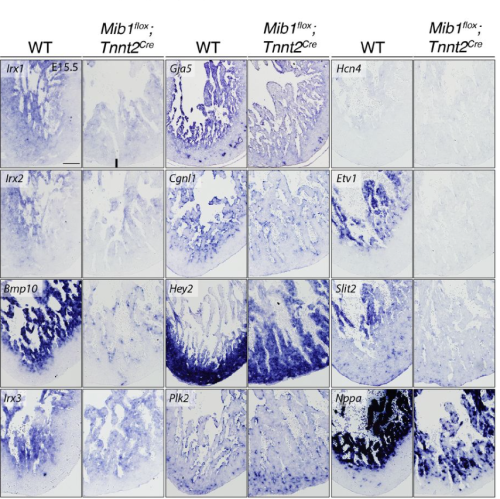
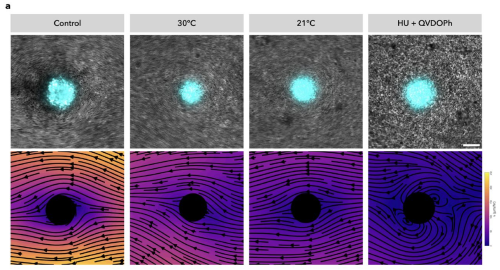
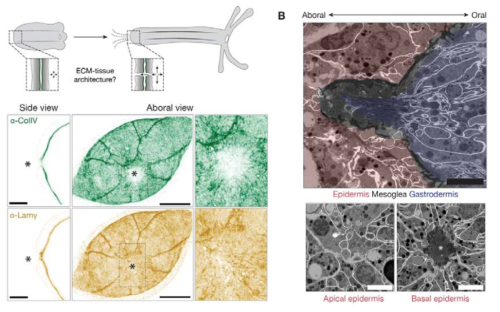
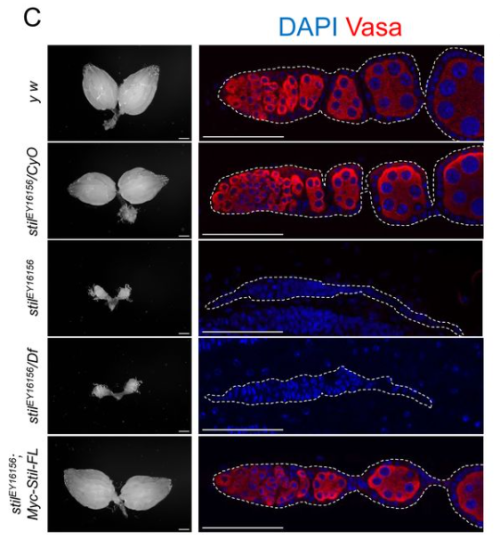

 (No Ratings Yet)
(No Ratings Yet)