In research that holds potential for prenatal health and brain injury, Scripps Research scientists identify cellular workings that stop and restart early brain development. Press release from Scripps Research, La Jolla.
We all know that food is essential to healthy development of the brain and body, especially in the earliest stages of life. But exactly how early brain growth is affected by nutrition is not as well understood, especially on a cellular level.
One reason for this lack of understanding is simply the difficulty of studying animals before they are born. But in a study involving tadpoles, which develop entirely outside of a mother’s womb, scientists at Scripps Research were able to unearth new findings about how brain cells respond to—and recover from—lack of nutrition.
“With tadpoles, we can look at early stages of brain development that are typically inaccessible to us,” says cell biologist Caroline McKeown, PhD, a senior staff scientist in the neuroscience lab of Hollis Cline, PhD, and lead author of the study. “This study showed us, for the first time in a vertebrate species, the cell signaling pathways that are integral to nutrient-responsive cell division in neural stem cells. These findings may lead to new approaches for starting and stopping cell growth in the brain.”
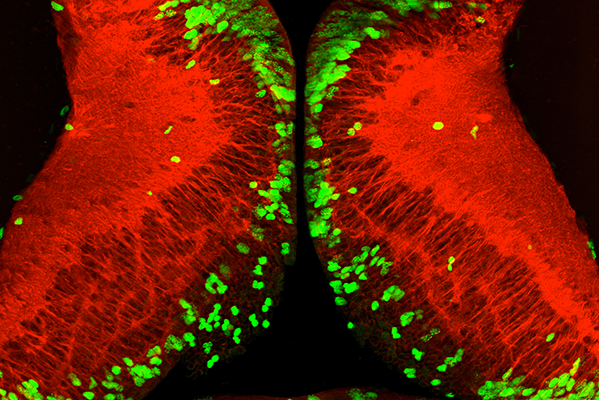
After periods without food, nutrition induces widespread proliferation of neural stem cells (green) in the tadpole brain. Mature neurons are shown in red. (Image courtesy of the Cline lab.)
The research, which appears in the journal Development, has multiple potential applications—including improved prenatal care in humans. McKeown said the findings also will contribute to on-going research in the lab on the role of neural stem cells in recovery from brain injury.
Typically, in a Xenopus tadpole and in most animals, stem cells known as “neural progenitors” flourish during early stages of development. These cells eventually mature into neurons, the cell type in the brain the controls thought and action.
In a previous study, McKeown and Cline found that when the tadpoles were deprived of food, their neural progenitor cells stopped dividing and their body growth decreased, but the animals remained alive and their behavior appeared normal. Surprisingly, if tadpoles were able to access food within about nine days, neural progenitor cells in the brain started dividing again and the tadpoles caught up to the growth state where they would have been if food had always been available.
What captured McKeown’s attention were the life-or-death questions: What triggered the neural progenitor cells to be able to divide again? And how did it work? In the new study, she and Cline identified the cellular mechanisms underlying this developmental response.
“We know a lot of these fundamental cellular events are conserved across animal species, so it’s possible that mammalian species are also capable of this kind of resilience to prenatal nutrient deprivation,” McKeown says.
Once the researchers found that early brain development could bounce back after periods without food, they wanted to understand what was happening on a cellular level to tell neural progenitors to stop dividing and to start back up. They traced it to a well-known signaling pathway known as mTOR (short for “mammalian target of rapamycin”), which is a central regulator of cell metabolism, growth, proliferation and survival.
Interestingly, even without providing the tadpoles with any food, their brains could be relaunched into growth mode by activating the insulin receptor that sits on the surface of neuronal progenitor cells, Cline says. Insulin is a hormone that allows cells to use sugars from food as energy and can activate mTOR signaling. Being able to bypass the need for food on a cellular level could advance medical therapies for poor nutrition.
By carefully tracking the neural progenitor cells over time, McKeown also discovered that they were poised to divide as soon as the nutrient signals reached them. This meant the cells had halted their progression when they were right on the verge of dividing. This is typically seen in cells under stress, and clearly starvation is a type of stress.
“Studying the ability of tadpoles to respond to environmental uncertainties helped increase our understanding of conserved cellular events controlling brain development,” McKeown says.
“The observation that food affects brain cell division was already known, but nobody dug more deeply into how food was having that effect,” adds Cline, Hahn Professor of Neuroscience and chair of the Department of Neuroscience in La Jolla. “We envision this knowledge becoming useful in understanding what can go wrong in the absence of maternal nutrition, and how important it is to respond quickly to a such an event.”
This research was supported by National Institutes of Health (EY011261, EY027473), Dart NeuroScience LLC and an endowment from the Hahn Family Foundation.
Biscutella laevigata – the subject of many of Saunders’ important plant breeding experiments. Photo: Atriplexmedia CC-BY-SA 3.0
The history of genetics has a few famous partnerships – such as James Watson and Francis Crick or Francois Jacob and Jacques Monod. But there’s one pair without whom this podcast wouldn’t exist at all, and that’s Edith Rebecca Saunders and William Bateson, who founded The Genetics Society one hundred years ago.
But while Bateson tends to get the glory, particularly for his popularisation of Gregor Mendel’s ideas about heredity, much less is heard about Saunders – the ‘mother of British plant genetics’, as she was referred to by JBS Haldane.
She was one of the first women to pursue a scientific education and research career at Cambridge University in an era when women were excluded from formal lectures and prevented from graduating. Rather than being a research assistant, Saunders was an equal colleague of Bateson.
She was a formidable teacher and researcher, eventually becoming director of the Balfour Biological Laboratory for Women in Cambridge, and made important contributions to genetics through her meticulous plant-breeding experiments. Saunders was also a key member of many scientific societies, and co-founded The Genetics Society together with Bateson in 1919.
If you enjoy the show, please do rate and review and spread the word. And you can always send feedback and suggestions for future episodes and guests to podcast@geneticsunzipped.com Follow us on Twitter – @geneticsunzip
By Amnon Sharir (UCSF), Allon M Klein (HMS), Ophir D Klein (UCSF)
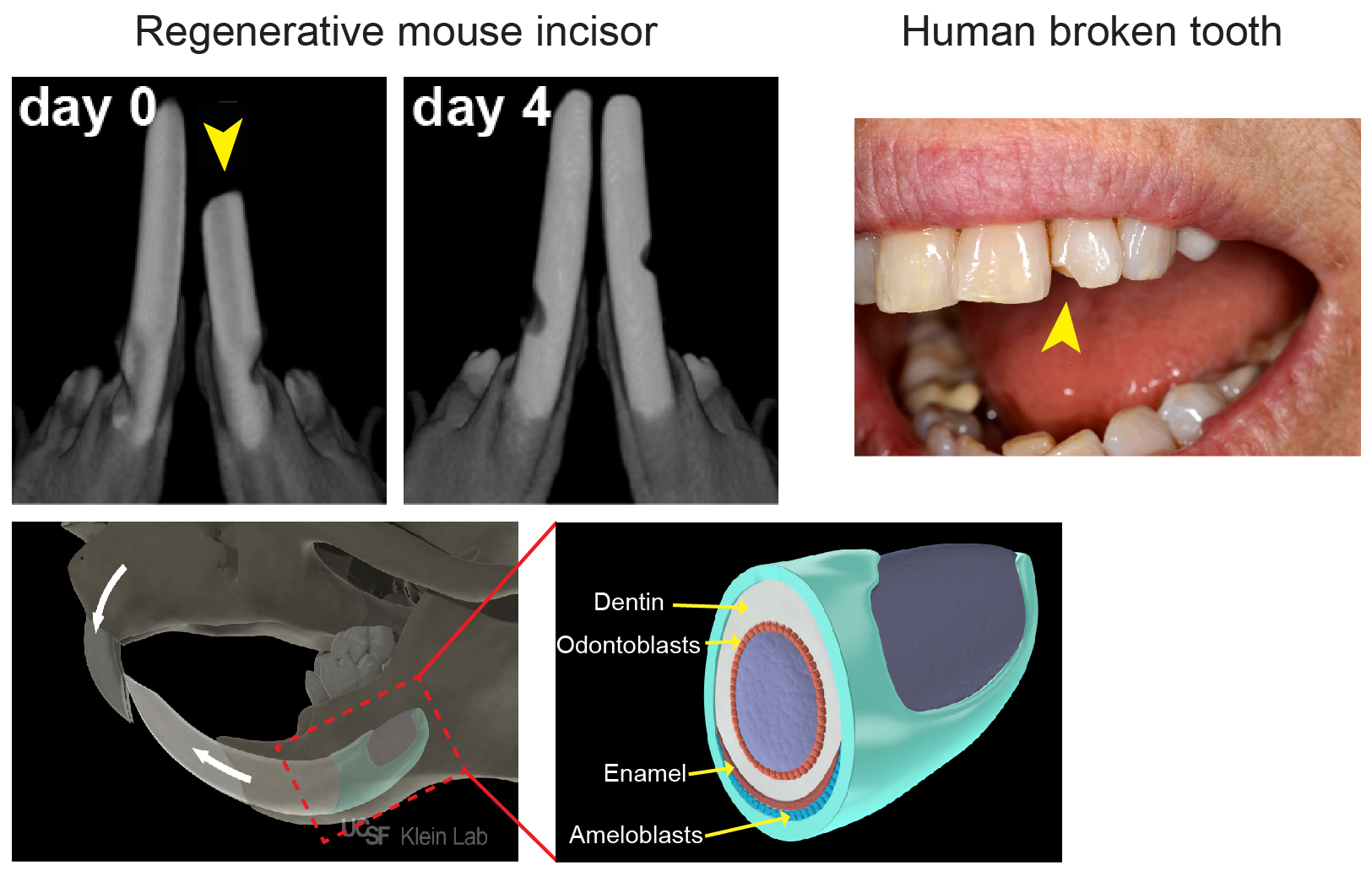
As most mouse geneticists know, treating a rodent with malocclusion of its front teeth can be a Sisyphean task: as much as one trims the tooth, it grows right back within a few days (Figure 1). The basis of this often-frustrating situation is that rodent incisors contain a group of adult stem cells (SCs) at the base of the tooth that drive continuous growth of the organ. These SCs produce a constant supply of new cells that replace the cells that are lost from the tip of the tooth due to normal wear or tooth breakage. In contrast, human teeth are quite different: once they are mature, they no longer grow. As a result, wear and tear of the enamel, the hard layer that covers the teeth, as well as diseases like caries that disrupt the enamel, cannot be self-repaired.
Figure 1: Top: In vivo µCT immediately (day 0) after trimming ~1 mm from the tip of one incisor, and 4 days later, demonstrating the remarkable regenerative capacity of the of mouse incisors. In contrast, a human broken tooth cannot self-repair. (Picture is a courtesy of Robert Ho (UCSF)). Bottom: Illustration showing the location of the incisor stem cell niche within the jaw bone. Magnification of the boxed area showing the epithelium (in green), the tissue which produces the ameloblasts that lay down enamel.
Because of their amazing regenerative abilities, our lab (klein.ucsf.edu), along with a number of others, has become very interested in mouse incisor SCs. We believe that understanding the mechanisms by which animals like mice normally renew their teeth will enable us to lay a foundation for human tooth regeneration. The mouse incisor provides a powerful platform for uncovering cellular behaviors, signaling pathways and transcriptional interactions that govern self-renewal and differentiation, and these findings have important implications for the field of SC biology. Most of the discoveries in the incisor epithelium, the tissue which produces the ameloblasts that lay down enamel (Figure 1), have resulted from candidate approaches. These studies identified several genes expressed by cells in the most proximal region of the incisor epithelium that give rise to the differentiated ameloblasts over a long period of time. Of note, these markers also largely, but not exclusively, co-localize with cells that are slowly cycling and therefore retain a label for long periods (label-retaining cells, LRCs). These experiments led to the widespread notion in the field that the SCs would be found among the LRCs. However, a number of key questions have remained unanswered, such as: Where exactly do the SCs in the tooth reside? How are SCs able to produce the correct number of ameloblasts to keep the tooth at a fixed length? How do SCs react to injuries such as trimming? Is there only one type of SC, or are there several discrete populations with variable functions?
Our adventure started back in 2013, when Allon Klein (klein.hms.harvard.edu) visited UCSF to share the quantitative approaches that his lab at Harvard was taking to study SC behavior. We realized immediately that these methods would be valuable for the mouse incisor field. The large, single-output incisor niche provides a contrast with the multiple, small, independent units found in other well-studied epithelial systems, such as the hair follicle and gut crypt. As such, it offers a rare opportunity to study the mechanisms that regulate cell number, as we can count the output of all of the SCs into the entire organ. The large size of the niche also offers the chance to study the unique challenges of maintaining proper SC numbers; for example, if a few crypts or hair follicles are lost, it is not a catastrophe for the animal, but loss of even one incisor SC niche would be fatal in the wild. Also, because teeth are largely dispensable for the survival of laboratory-housed mice, we can injure them and study their regeneration over extended periods without killing the animal.
At least initially, we assumed that our undertaking would involve a relatively straightforward analysis, as we thought that we knew the location of the SCs and their proliferation dynamics, and we had in our hands several inducible genetic tools for in vivo lineage tracing of these cells. As with many scientific adventures, however, things were more complicated than they seemed at first.
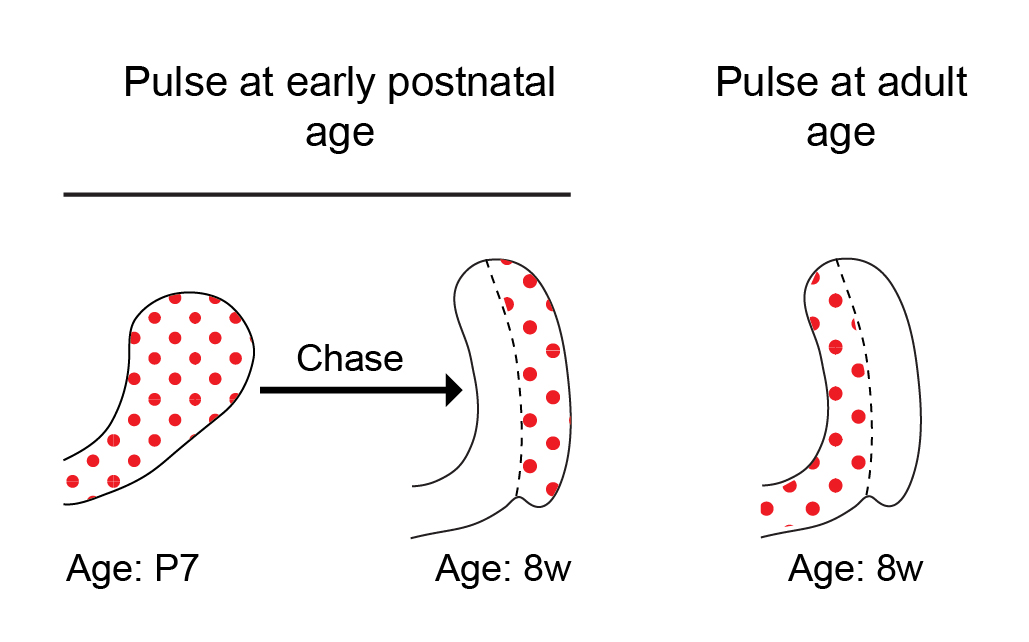
Historically, a classical view in the SC field in general has been that tissue SCs are slow cycling LRCs, and this was also true in the incisor field. While this view has evolved over the last decade, it is still often thought that SCs cycle less frequently than their immediate progeny, typically known as transit amplifying cells. In previous studies conducted by our lab and others, we pulsed mice with 5-bromo-2′-deoxyuridine (BrdU) to label proliferating cells during the early postnatal period and then aged the mice for several weeks. Cells within the presumed SC region at the base of the tooth, called the outer enamel epithelium, acquired BrdU label and retained it for several months. The extended retention of the BrdU label was interpreted to mean that these cells are SCs. However, to our surprise, when we pulsed 8-week-old mice with BrdU, the outer enamel epithelium was not labelled at all (Figure 2). No matter how much BrdU (or EdU, 5-ethynyl-2′-deoxyuridine) we used, the cells in the LRC region that we had presumed to contain an active SC population remained unlabeled! Lack of cell cycling in this region did not initially make sense to us, because the incisor fully turns over every 4-6 weeks, and therefore, we predicted the cycling time of the SCs to be relatively short — similar to that in other fast-cycling tissues, such as the epidermis and the intestinal epithelium.
Figure 2: Schematic models showing broadly distributed cycling cells in the incisor growth region when mice are pulsed at the perinatal period (left). Cells within the outer enamel epithelium acquired BrdU label and retained it after 7 weeks. In contrast, when mice are pulsed at 8 weeks of age, cycling cells are absent from the outer enamel epithelium (right).
The dramatic transition from active proliferation to a dormant outer enamel epithelium over the first few weeks of the mouse’s life prompted us to define the time period during which the incisor is in steady state. To this end, we performed a series of 3D micro computed tomography (µCT) analyses of the incisor during postnatal development. We found that the incisor SC niche, called the labial cervical loop, is in steady-state between 8 and 16 weeks of age. During this period, the incisor growth rate is minimal, the cervical loop size is constant and proliferation is stable. We reasoned that the LRCs identified in previous incisor experiments represent post-mitotic cells that proliferate only during the postnatal expansion period, and we therefore focused our analysis on the steady state period.
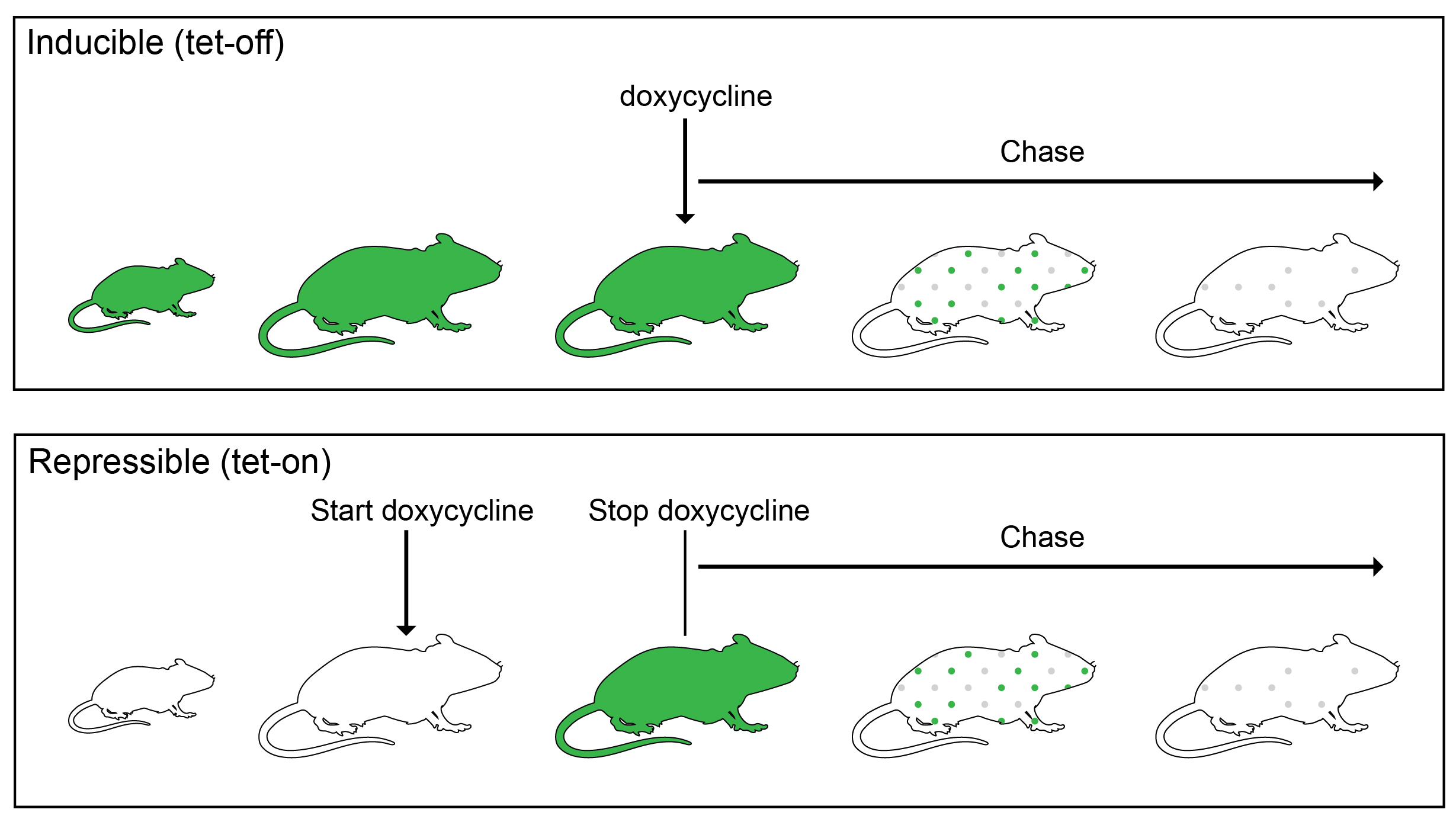
Some cells must be cycling to incorporate a BrdU or EdU label. Since cells were not cycling in the outer enamel epithelium during our desired nucleotide pulse period, we decided to instead use an H2B-GFP label dilution system. In such a system, expression of inducible or repressible H2B-GFP, driven by a tet-response element, is controlled by a tetracycline (Tet)-transactivator (rtTA in Tet-On or tTA in Tet-Off). The proliferation dynamics of the organ can be tracked, because during the chase period, the resulting GFP is diluted by half during each cell division (Figure 3).
Figure 3: Schematic models showing the inducible and repressible H2B-GFP systems. In a tetracycline-inducible (tet-off) double transgenic mouse system (top), constitutive GFP protein expression is shut off by treatment with doxycycline, while in the repressible (tet-on) system, doxycycline administration turns on GFP expression (bottom).
We initially used a K5tTa;tetOff-H2B-GFP, in which constitutive GFP protein expression in the incisor epithelium can be shut off by treatment with doxycycline1. However, we soon discovered that the incisors of the tetOff-H2B-GFP mice were abnormal: they were smaller, had cracks and frequently broke in our hands while we prepared them for analysis, and appeared chalky white (in mice, this is not a sign of good tooth hygiene, but rather an indication of lack of mineralization). Our μCT analysis confirmed that indeed there was a significant decrease in incisor volume and enamel density. So, this line could not be used to determine incisor cell kinetics (anomalies of other organs, such as the cornea, have been noted by others2, which perhaps is due to very high GFP levels in early life that are destructive to some organs). We then switched to the repressible TetOn-H2B-GFP line, in which H2B-GFP is activated by doxycycline treatment3. To our relief, the incisors of these mice appeared normal. However, again with this line, the outer enamel epithelium was devoid of labeling, unless we pulsed the mice during the perinatal period. A similar lack of cell labeling in other tissues, such as the olfactory bulb and the spinal cord, has been noted by the researcher who developed the line and was attributed to an inability of doxycycline to cross the blood-brain barrier4. In our case, we think that the cells are not labelled in the incisor because they are so quiescent that they don’t replace unlabeled histone H2B with the labeled one.
We decided to stick with the TetOn-H2B-GFP line, in which we were able to label the active region of the incisor epithelium and asses proliferation dynamics using a short pulse of doxycycline. We first used flow cytometry to sort the epithelial cells during a chase period and measure their GFP intensities, and then we modeled how the GFP signal distribution should change between chase day 1 to chase day 7 to infer two parameters: the fraction of cells that are proliferating, and their division rate. The model we specifically considered assumed that the number of divisions of any individual cell was Poisson-distributed, which gave a very good fit to the data. The best fit was when the average number of divisions in six days was around 3 (2.98 ± 0.20), and that initially 60±15% of cells were post-mitotic.
While the sorted GFP results provided us with valuable information regarding proliferation dynamics in the incisor epithelium, these data did not tell us anything about the spatial distribution of division. We decided to investigate the proliferation dynamics of the entire organ in situ. Using a two-photon microscope, we acquired images of the entire proximal region of the incisor at 45 minutes and 48 hours after EdU injections. We chose 48 hours, because at this time-point the cells have divided once on average, and no cells have yet been lost due to distal movement along the incisor length. It was clear from looking at the images that many EdU cells moved from the active proliferative region to other areas within the cervical loop. However, quantitative analysis of the location and level of EdU in each cell remained a challenge. This was especially true in our enormous region of interest, which contained about 175 images of 900 µm (length) X 900 µm (height) X 350 µm (width), with very densely packed cells and variabilities in laser intensity, due to the need to penetrate deeper sections of the sample. At this point, we were introduced by our colleague Jacqui Tabler to Kyle Harrington (kyleharrington.com), an expert in image analysis from the University of Idaho, who ultimately developed a segmentation pipeline tailored to our needs.
The software that Kyle developed uses a region competition algorithm to express image intensity and statistics of candidate regions as energy terms that are iteratively refined and balanced until the segmentation converges. After filtering regions based upon the expected size of cells, it measures the EdU signal at the center of the cell to determine proliferation status (Figure 4). The result of this image analysis pipeline was a 3D map of the spatial distribution of individual cells and their corresponding EdU signal, which confirmed our observations from the thin sections and fit well with our single cell RNA sequencing data regarding the sites of active cell division and the flow of cells from these sites to other regions in the incisor epithelium.
Figure 4: An example of the segmentation pipeline shown in video which was rendered in the freely-available ImageJ-based tool, SciView5. Cell membrane labeled in magenta and the center of segmented EdU+ cells shown in yellow.
In this blog post, we have shared several aspects of our thought process during our recently-completed project, in the hope that other researchers will find it useful when planning similar experiments or facing unexpected outcomes in their analyses of epithelial SCs. In our recent paper6, we integrate our quantitative proliferation kinetics with unbiased single cell RNA-seq analysis and genetic lineage tracing. We were able to uncover cell behaviors that upended the reigning dogma about the identity, location and function of progenitor cells in our tissue of interest.
The Echeverri lab at the MBL seeks a highly motivated individual to join the Eugene Bell Center for Regenerative Biology and Tissue Engineering as a Postdoctoral Researcher. The successful candidate will work on neurodegeneration and regeneration in axolotls.
The specific goal of the project is to examine how pathways that are essential for regeneration have evolved in different species with different regenerative capacity; focusing specifically on neural regeneration and spinal cord regeneration.
Basic Qualifications:
Applicants should have a Ph.D. in a biology related field. Must have prior experience working in the field of cell or developmental biology, as well as experience with molecular biology. Must be independent, enthusiastic, self-motivated, productive, and enjoy working in a highly collaborative environment.
Preferred Qualifications:
The ideal candidate will have direct experience with working in vivo in an animal model. Previous experience with molecular biology, generating transgenic lines and CRISPR knockins/outs and imaging would be a plus.
Apply online:
https://www.mbl.edu/hr/employment/
Two postdoc positions are available at Columbia University’s Zuckerman Institute in New York City. Both positions focus on Drosophila neuroscience. One aims to better understand how adult neural circuits are assembled during development (e.g. Venkatasubramanian et al.); the second studies how these circuits are used as adult flies walk (e.g. Howard et al.). Please email the PI if interested in joining our team!
A fully funded bioinformatics postdoctoral position is available in the Laboratory of Regulatory Evolution (Tschopp lab) at DUW Zoology, University of Basel, Switzerland.
We study the gene regulatory mechanisms of cell fate specification in the vertebrate skeleton. Depending on anatomical location, the vertebrate skeleton develops from three distinct progenitor populations – neural crest, somitic and lateral plate mesoderm. We are interested in the gene regulatory network (GRN) dynamics that transcriptionally re-code these distinct progenitor pools towards functionally analogous skeletal cells.
As part of a larger Swiss National Science Foundation (SNSF)-funded project, we are looking for a bioinformatics postdoc to analyze developmental single-cell RNA-seq and single-cell ATAC-seq data, followed by CRISPR/Cas9 perturbations, to infer the GRN dynamics underlying this progenitor convergence towards a common skeletal cell fate. These analyses will be performed in collaboration with the group of Prof. Erik van Nimwegen, experts in computational GRN inference, at the Biozentrum Basel. The project builds on a solid foundation of confirmed preliminary data. For more information please visit http://evolution.unibas.ch/tschopp/research/
The successful candidate will hold a PhD with a strong background in one or several of the following fields: bioinformatics; single cell analyses; statistics; computational data analysis; as well as interests in developmental and molecular biology. Good communication skills in oral and written English are essential.
We offer a highly interactive and interdisciplinary research environment, state-of-the-art technology platforms, attractive employment conditions and very competitive salaries by international standards. Full funding is available for 1+2 years.
Please send your application as a single PDF with a brief statement of motivation, a current CV and contacts for at least two references to patrick.tschopp@unibas.ch . Evaluation will begin on December 1st 2019 and suitable candidates will be contacted shortly after. Earliest starting date is January 1st 2020.
Venue: UCL Institute of Child Health, Guilford Street, London, WC1N 1EH
Organisers: Nick Greene (UCL), Cynthia Andoniadou (KCL), Andy Copp (UCL)
The Mammalian Genetics and Development Workshop is an annual meeting focusing on the development and genetics of mammals. The Meeting is based on the submitted abstracts, and include diverse topics ranging from mammalian development (not exclusively human or mouse) and identification of disease genes and developmental mechanisms, to human genetics and epigenetics. Other model systems (including Drosophila, zebrafish and chick) are also welcomed where these relate to general developmental questions and/or disease models.
The meeting will be similar format to the 29th Meeting in 2018, which had a fantastic set of short talks covering various aspects of developmental biology, mouse models of human disorders and genetics of human disease (abstracts from the 2017 and 2018 meetings are published at https://doi.org/10.1017/S0016672317000076 and https://doi.org/10.1017/S0016672319000016).
The workshop is traditionally a venue for post-docs and PhD students to talk rather than laboratory heads and is an excellent training ground and a friendly, informal forum. In keeping with this objective, we offer TWO PRIZES of £150 to individual post-graduate/post-doctoral presenters. In addition, thanks to the generosity of Mammalian Genome (https://link.springer.com/journal/335) publishers, we will offer an additional PRIZE of £150.
The Deans laboratory at the University of Utah is recruiting motivated postdoctoral fellows to fill NIH-funded research positions investigating the role of Planar Cell Polarity (PCP) signaling during Inner Ear development and cochlear innervation. We apply basic developmental biology, mouse genetics, and biochemical approaches to define mechanisms of sensory receptor differentiation and innervation using the mouse as a model system. Please consider these recent examples of our discoveries in this area:
“Frizzled3 and Frizzled6 Cooperate with Vangl2 to Direct Cochlear Innervation by type II Spiral Ganglion Neurons” Journal of Neuroscience (2019)
“A non-autonomous function of the core PCP protein VANGL2 directs peripheral axon turning in the developing cochlea” Development (2018)
The Deans lab is located in a highly dynamic research environment hosted by the Department of Neurobiology & Anatomy. In addition, Utah provides unparalleled lifestyle and outdoor recreation opportunities for a superior work/life balance. Candidates should carry a PhD in the areas of developmental, cell or neuro biology.Salary will be commensurate with prior research experience and NIH guides.
PhD studentship to start in October 2020 with Grant Wheeler at the University of East Anglia in Norwich.
The Neural Crest and Placodes are groups of cells found only in vertebrates, specifically in the embryo. They originate at the neural border between the ectoderm and neuroectoderm. Once specified the Neural Crest undergo an epithelial to mesenchymal transition (EMT) and then migrate to various parts of the embryo where they differentiate into important issues such as parts of the heart, the peripheral nervous system, the cartilage of the face and pigment cells. Placodal cells differentiate into sensorial organs such as the eyes, ears and nose. The Neural Crest and Placodes are therefore of importance for normal development and errors in their development are the cause of many birth defects.
Understanding the regulatory elements, such as enhancers, required for specification of the Neural Crest and Placodes is important in order to understand how they are first specified and then induced during development. Understanding these processes will help in developing techniques to engineer specific cells and tissues that the neural crest and placodes give rise to and which could be used in stem cell and regenerative therapies. ATAC-seq is a method to identify ‘open’ regions in the chromatin landscape which can correspond to active enhancers and promoters. We have previously carried out ATAC-seq on Xenopus embryonic tissue induced to form Neural Crest and Neural Ectoderm to determine active enhancers and promoters. In this project the student will generate ATAC-seq data on material induced to form placodal tissue. They will use bioinfomatics to analyse the data and compare it to the neural crest data. Differential analysis will uncover specific neural crest and placodal enhancers. Potential enhancers will be tested and validated using transgenic and CRISPR/Cas9 technologies.
For more information about how to apply please visit
The Posfai Lab at Princeton University (www.Posfailab.org) is looking to recruit a highly motivated postdoctoral fellow to study the molecular and cellular mechanisms of cell fate choice and emergent organization during early embryonic development, using the preimplantation mouse embryo as a model system. The project will combine genetic engineering and quantitative, high-resolution live imaging using light sheet microscopy to understand the finely coordinated spatiotemporal dynamics of developmental events on a molecular, cellular and whole embryo scale.
The postdoctoral candidate should have a strong interest in developmental biology and genetics. Experience in light sheet microscopy and/or computational image analysis is preferred, but not necessary. Motivation and excellence is valued more than previous field of study.
The candidate will benefit from an interdisciplinary and collaborative environment at Princeton and the vibrant and supportive atmosphere of a junior lab. Researchers at the rank of Postdoctoral Research Associate are ordinarily appointed for one year at a time. Appointments are reviewed annually to consider reappointment and salary level. The position is benefits-eligible.
To apply, email your CV and cover letter explaining your interests and motivation to Eszter Posfai (eposfai@princeton.edu) and arrange for three reference letters to be sent on your behalf.
 (No Ratings Yet)
(No Ratings Yet)
 (5 votes)
(5 votes){kind=link}