This is the story of a journey that started a few years back with the finding of a rare community of single-celled organisms in a small and isolated nature reserve’s pond in Dorset. This microbial community was unusual in that the majority of these unicellular organisms were ‘protozoa’ (microbes that feed on other microbes) that harboured other microscopic life inside them; particularly interesting were those that harboured microscopic algae. Furthermore, most of the species had only been reported from one other location in the world – Germany!
Microscopic algae are plants and thus carry out photosynthesis. If we remember from our school years, photosynthesis is an outstanding biological mechanism allowing plants to produce their own food (i.e. carbohydrates) by using the sun’s energy; the waste product of this mechanism is oxygen, which is released into the environment. Interestingly, the protozoa that harbour the ‘endosymbiotic’ algae benefit from their photosynthesis in two ways; the first is as a nutritional source by taking up the carbohydrates produced through algal photosynthesis. This is not new as it has long been known that the majority of protozoa with endosymbiotic algae benefit from such a nutritional boost. However, this rare community of protozoa in a Dorset pond led us to discover that some protozoa did not use the photosynthetic nutritional rewards, but rather the oxygen produced as the end product of algal photosynthesis.
The protozoa with endosymbiotic algae are found in highly-productive ponds where oxygen levels are depleted; having an association with an oxygen-producing machine (the microscopic algae) seems the perfect arrangement. An example of this was the rare ciliated protozoan Loxodes rostrum (pictured) – an extremely unusual ciliate in many ways. Firstly, it has seldom ever been recorded (only in Germany and the UK, so far); secondly, it is a ciliate that harbours several different species of microscopic algae (most ciliate-algae associations are with one species only of the genus Chlorella). Unfortunately, the pond in Dorset dried out before we could finish our fascinating investigations on this unique species.
Fast forward a few years and I was invited to the Workshop Symbiosis in the Microbial World: from Ecology to Genome Evolution in November 2017, organised by The Company of Biologists on the impressive grounds of Wiston House (West Sussex). The site has an old and productive pond with ample overhanging vegetation – just like the pond in Dorset! Would this other pond have Loxodes rostrum? We collected samples for the workshop’s microscope session on ‘Life under the Lens’ that I was running, and voila – there it was, adding a new location to its biogeographic map. This finding at Wiston House allowed us to finalise the investigations on this ciliate species, including looking at its DNA sequence. The research is in collaboration with Prof Martin Embley and Dr Kacper Sendra at the Institute for Cell and Molecular Biosciences at Newcastle University.
We thank The Company of Biologists, particularly Nicky Le Blond for the enthusiasm and for facilitating pond sampling at Wiston House, and to the Wiston House estate for allowing access to the pond.
A 60% permanent position as Facility Technician is available from July in the newly established research group of Dr. Pawel Burkhardt at the Sars Centre. Work in the group focuses on the evolutionary origin of synapses and neurons, using choanoflagellates, sponges and ctenophores as main model systems. The Sars Centre belongs to the University of Bergen, and is partner of the European Molecular Biology Laboratory (EMBL).
Tasks:
The main responsibilities of the position will be to assist in the building and daily running of a sponge and ctenophore facility, including:
Culturing of algae and copepods for feeding animals
Feeding and breeding of animals
Operation and maintenance of the animal facility
Maintenance of equipment and building of new equipment to improve functioning of the facility
Periodic field collection of sponges and ctenophores
Some weekend duties might be requested
Qualifications:
Essential requirements for the successful candidate are to be able to demonstrate reliable working habits, independency, initiative and the ability to interact in a productive manner with other group members
Prior experience with building and/or maintaining animal facilities is required
Experience maintaining algae and/or zooplankton cultures is desirable
The minimum formal requirement is a high school education
The applicant must be able to communicate (written and spoken) in English and Norwegian
We offer:
A challenging position with interesting tasks in an internationally acknowledged scientific environment
Salary as Senior Research Technician (“seniorforskningsteknikar”, code 1513) from grade level 40, based on a formal assessment of civil service seniority and qualifications. Currently equal to NOK 376.000
Enrolment in the Norwegian Public Service Pension Fund (Statens pensjonskasse, SPK)
A position in an inclusive workplace (IA enterprise)
Good welfare benefits
Application letter stating your motivation for applying to this position and why you feel you are a good candidate for this position
Overview of your education and work experience (CV)
Certified copies of educational transcripts and other relevant documents
Two references (names and contact information)
Send your application electronically via JobbNorge by clicking on the button “Apply for this job” – see here. Please note that applications will be assessed only with the information available in JobbNorge when the deadline expires. It is the applicant’s responsibility to ensure that all relevant attachments are submitted by the deadline.
Applications by e-mail only will not be considered.
Application deadline: 12 June 2018
General information:
Detailed information about the position can be obtained by contacting: Group Leader Pawel Burkhardt, tlf.: +47 55 58 43 57, email: Pawel.Burkhardt@uib.no.
The state labour force shall reflect the diversity of Norwegian society to the greatest extent possible. Age and gender balance among employees is therefore a goal. People with immigrant backgrounds and people with disabilities are encouraged to apply for the position.
We encourage women to apply. If multiple applicants have approximately equivalent qualifications, the rules pertaining to moderate gender quotas shall apply.
The University of Bergen applies the principle of public access to information when recruiting staff for academic positions.
Information about applicants may be made public even if the applicant has asked not to be named on the list of persons who have applied. The applicant must be notified if the request to be omitted is not met.
Further information about our employment process can be found here.
Primary supervisor: Professor Claudio Stern FMedSci FRS, Department of Cell and Developmental Biology, University College London
Project title: “Dynamics of cell behaviour during somite formation”
A studentship funded by the Anatomical Society is available in Claudio Stern’s lab. The project will study the mechanisms of somite formation, to elucidate the molecular and physical mechanisms that control somite size, shape and regional identity and the role of the “segmentation clock” and local cell-cell interactions in this process. It is a multi-disciplinary project and will involve advanced live imaging (including super-resolution microscopy in vivo), molecular biology, biophysics and some computational modelling. The project is likely to involve travel to work with collaborators in the USA and/or Singapore.
Conditions and requirements: This PhD studentship is open to British, Irish or European citizens who have spent at least three years at a British or Irish institute of higher education. It is funded by the Anatomical Society of Great Britain and Ireland and offers a stipend (tax free) of £16,553 per annum (revised annually), university fees (UK/EU rate) and a contribution to research expenses as well as funds to travel to meetings. Funding is for 3 years but it may be extended to a fourth year if necessary. Candidates should have a 2.1 (or equivalent) degree or better in a Biomedical, Physical or Computational science-related area and strong interest in developmental biology, ideally along with some laboratory experience working in a biomedicine-related research project. Experience with microscopy, programming (preferably PYTHON) and/or other computational/mathematical skills, are not essential but will be an advantage.
Starting date: 1 October 2018 or earlier by arrangement.
To apply (with a cover letter, CV a brief statement of your interests and the names and contact details of two academic referees), or for further information please contact Prof. Stern: c.stern@ucl.ac.uk
There is no formal closing date for applications but a student will be appointed as soon as a suitable, high quality candidate is identified.
The group of Sara Wickström, funded by an ERC Consolidator Grant in 2018, is looking for
ERC-Funded Post-doctoral Researchers in Stem Cell Research
for 2 years starting from 1.8.2018 with possibility of extension up to 4 years.
We are looking for talented and highly motivated scientists to join a young and dynamic research team in the analysis of stem cell regulation in tissue morphogenesis, maintenance and cancer. We employ a wide range of techniques, including mouse genetics, organoid and bioengineering tools, live cell imaging, mass spectrometry and single cell sequencing to study how stem cell and cancer stem cell fate is regulated through interactions with their niche.
The applicant is required to hold PhD in biology, genetics, bioengineering, biochemistry, or a related field, as well as strong written and oral communication skills. The working language is of the lab is English; knowledge of the Finnish language is not necessary. Experience in cell biology is required, experience in computational data analysis and next generation sequencing is an advantage.
To apply:
To apply, please submit your application using the University of Helsinki electronic recruitment system https://www.helsinki.fi/fi/avoimet-tyopaikat/post-doctoral-researchers-in-stem-cell-research. Upload a single pdf-file containing a one-page letter with a personal statement outlining your research interests and relevant work experience, your CV and bibliography, contact information for 2-3 references.
Helsinki Institute of Life Science, HiLIFE, is a new life science institute at the University of Helsinki, a leading Nordic university with a strong life science research. HiLIFE operates mainly in the campuses of Meilahti and Viikki. In Meilahti, top-notch medical research, education and care meet daily to benefit of the patients and the entire society. Meilahti provides access to state of the art core facilities for imaging, genomics, proteomics and metabolomics, as well as bioinformatics. It is also home to the Faculty of Medicine and the Helsinki University Central Hospital (HUCH) which makes it an interactive research community with interests in a broad range of topics from fundamental biology to clinical applications.
Translational stem cell research for type 1 diabetes therapy is in prime focus with a new strategic alliance between The Novo Nordisk Foundation Center for Stem Cell Biology (DanStem) and Helmholtz Zentrum München (HMGU). Through the establishment of a strong R&D platform for manufacturing of human pluripotent stem cell-derived pancreatic islet cells for future stem cell-based cell therapy in type 1 diabetes, the cooperation offers a unique opportunity for advancement of translational stem cell diabetes research and therapy.
Diabetes is one of the primary worldwide health challenges. Patients with type 1 diabetes (T1D) suffer from a destruction of insulin-producing cells of the pancreas. There is currently no cure for the disease; however, cell therapy in the form of pancreatic islet transplantation can prevent uncontrolled fluctuations of blood sugar levels. The ability of human pluripotent stem cells to self-renew and differentiate into pancreatic islet cells offers a unique, exogenous scalable source of islet cells for such cell therapy in T1D. Human pluripotent stem cell-derived insulin-producing beta cells have yet to be transplanted into patients, and the new collaboration between UCPH and HMGU provides the essential steps to the first clinical trial.
A platform for translational research The key element of the new alliance is a platform for translational research associated with the manufacture of cells, in a ‘Good Manufacturing Practice’ (GMP)-dedicated facility. This platform will provide scalable GMP-grade cell production, regulatory expertise, and the testing of stem cell-derived insulin-producing cells, thereby establishing an optimal, complementary arrangement for translational research and accelerating the planning of the first-in-man clinical trial using human pluripotent stem cell-derived insulin-producing beta cells.
“We believe that we are well on track with our efforts to develop a stem cell-based replacement therapy and that our latest breakthroughs will pave the way for initial clinical trials. It is all the more important for us to exchange ideas with other top diabetes researchers, and we are therefore delighted to be connected with Helmholtz Zentrum München – a beacon of excellence in diabetes research,” says Professor and Executive Director of DanStem Henrik Semb.
At DanStem, Professor Semb leads a research group focused on the translation of basic research discoveries into new human pluripotent stem cell-based treatment of T1D. With the strategic partnership, he will also drive the Institute of Translational Diabetes Research at Helmholtz Zentrum München, the German Research Center for Environmental Health.
Partnership for the benefit of patients Strategic alliances are indispensable in science, especially to bring together complementary expertise and capabilities for success in translational research. For DanStem and SUND the strategic alliance with HMGU is an important part of becoming a global leader in both basic and translational stem cell research. The partnership advances the development of treatment options for T1D, moving toward a more sustainable future in cell replacement therapy.
See more information here: http://danstem.ku.dk/news/danstem-embarks-on-a-strategic-partnership-with-helmholtz-zentrum-mnchen/
In January, students from across the Americas gathered in Chile to participate in the International Course on Developmental Biology, an EMBO Practical Course held at the Marine Biology Station of Quintay (CIMARQ). In the course of two weeks of intensive training (you can read a wonderful summary of the meeting from the students in this previous Node post), the students generated some beautiful images of development in different species. We’re excited to launch a competition to pick an image taken by the students to become the cover of an issue of Development, and thus immortalised in print and on screen!
Check out the images below (clicking on them gets you a full size version), and then vote using the poll at the bottom of the post. Happy voting!
1. Pax3 in Drosophila embryos of different stages
Maria Belen Palacios
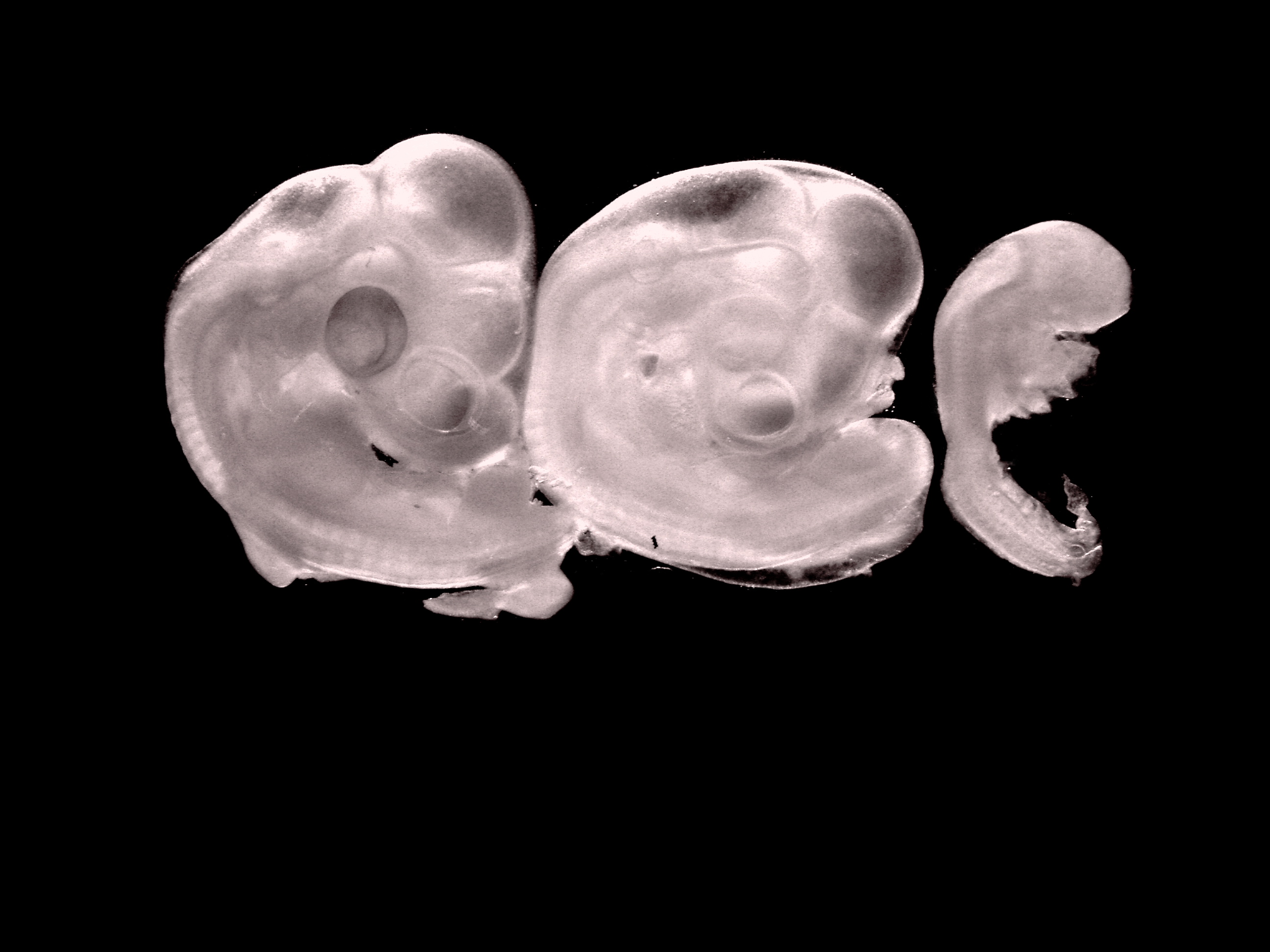
2. Chick embryos, stages 25, 22 and 16 HH
Estefanía Sánchez Vásquez
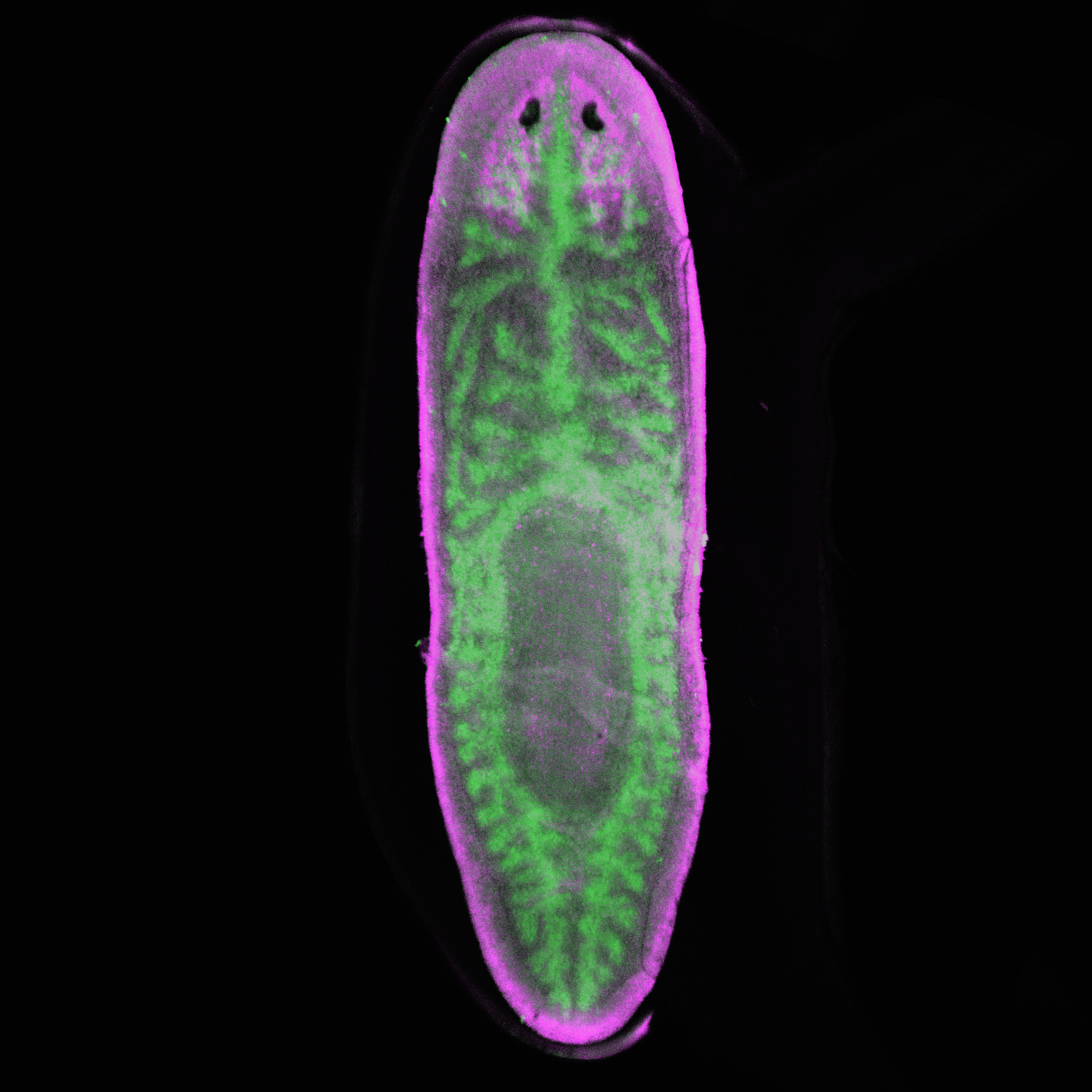
3: Planarian (gut in green, nervous system in pink)
Luiza Saad and Estefanía Sánchez Vásquez
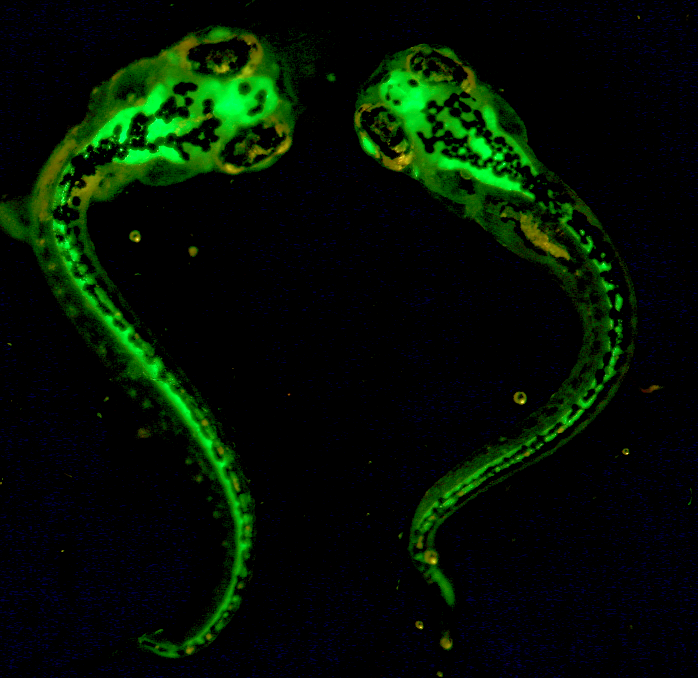
4: Zebrafish embryos injected with GFP
Maria Belen Palacios
5: Drosophila embryo labelled with Dapi, 312 and Elav
Eugene Tine
6: Sea anemone with an Amphineura (chiton) in the center
Maria Belen Palacios
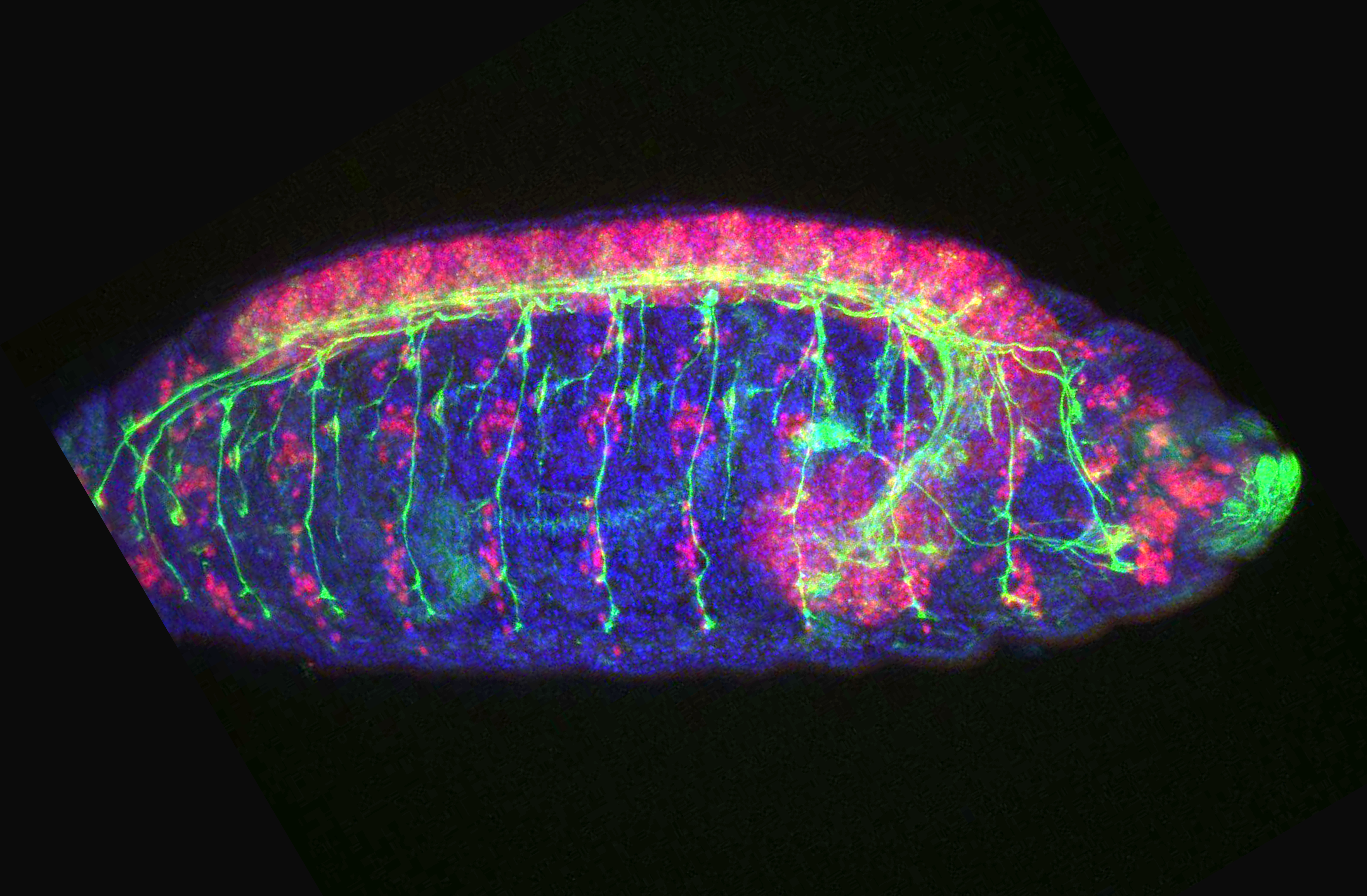
7: Drosophila embryo (blue: DAPI, green: motor neuron axons, pink: motor neuron nuclei)
A postdoctoral position is available in the laboratory of Philippe Soriano to study FGF and PDGF regulated craniofacial morphogenesis in the mouse embryo. Possible projects focus on intracellular signaling pathways using receptor point mutant mice and proteomics; the impact of ERK1/2 and PI3K signaling dynamics on morphogenetic movements; or differential signaling mechanisms underlying ligand specific responses. A strong background in cell signaling or developmental biology is desired. Expertise in molecular/cell/systems biology, imaging technology, or mouse genetics are valuable but not required.
For more information, please visit our laboratory website at labs.icahn.mssm.edu/sorianolab. We benefit from a highly interactive scientific environment in the Department of Cell, Developmental, and Regenerative Biology at Mt. Sinai (www.mssm.edu/drb), and an outstanding developmental biology community within the city. In addition, New York City offers unequaled cultural and recreational opportunities.
Creative, interactive but independent, and highly motivated applicants are in particular invited to apply. Please send a letter of application including a brief description of previous research experience and interests, a CV, and the name and contact information of three references to Philippe Soriano at philippe.soriano@mssm.edu
The vertebrate lymphatic vascular network provides crucial circulatory and immune functions but its developmental origin has been a contentious issue, in particular the question of whether lymphatic endothelial cells have an exclusively venous origin. A new paper in Development addresses this issue in the dermis of the mouse embryo. To find out more about the story behind the work, we caught up with first author Cathy Pichol-Thievend and last authors Natasha Harvey (Associate Professor at the Centre for Cancer Biology, SA Pathology and University of South Australia) and Mathias Francois (Associate Professor at the Institute for Molecular Bioscience, The University of Queensland).
Mat and Natasha, can you give us your scientific biographies and the questions your labs are trying to answer?
MFIn 2004, I was awarded a PhD in molecular genetics from the University PARIS VI (Pierre et Marie Curie). During this period I worked on nuclear receptors PPARs and their role in cartilage inflammatory response (INSERM UMR-S747- Paris V). At the end of 2004, I moved to Australia to gain postdoctoral experience with Professor P. Koopman, an expert in the field of sex determination and SOX transcription factors (2005-2011, The Institute for Molecular Bioscience, The University of Queensland). During that time the key discovery from my work identified the transcription factor SOX18 as a molecular switch that induces the lymphatic differentiation program by turning on Prox1 expression in pre-existing veins.
In 2012, I set up my own independent research group at the IMB. Since then we have designed a novel molecular strategy to pharmacologically manipulate developmental transcription factor activity. We successfully identified and characterised small compounds that interfere with SOX18 activity. This gives us a unique handle to complement classic genetic methodologies with pharmacological approaches to study vascular development. The research interest of my group revolves around on how endothelial cell fate is dynamically regulated at a transcriptional level. To decipher how transcription factors orchestrate lymphangiogenesis and subsequent assembly of vessel networks we rely on the utilisation of mouse and zebrafish model systems.
NH After a PhD with Sharad Kumar focussed on defining the molecular mechanisms underlying caspase-2 activation during apoptosis, I wanted to investigate the role of caspase activation during embryonic development. To this end, I cloned a novel Drosophila caspase, Damm, and in collaboration with Helena Richardson’s group, investigated the role of Damm during Drosophila development. This work ignited my interest in developmental biology and stimulated me to embark on postdoctoral training with Guillermo Oliver at St Jude Children’s Research Hospital. When I joined Guillermo’s lab, the team had just published a landmark paper in Cell demonstrating that the homeobox transcription factor Prox1 is crucial for development of the lymphatic vasculature during embryogenesis in the mouse. The scene was set to investigate how Prox1 controls embryonic lymphangiogenesis and that was the focus of my project. My postdoc in Guillermo’s lab was transformative, it was my introduction to both mouse embryogenesis and transcription factors and I was hooked! I remain completely fascinated by developmental biology!
After four years in Guillermo’s lab I returned to Australia to the Centre for Cancer Biology in Adelaide, to establish my independent laboratory focussed on understanding how the lymphatic vascular network is built during development. A really rewarding aspect of our recent work has been our synergy with Hamish Scott’s genetics team at the Centre for Cancer Biology. Together with Hamish’s group, we work to dissect the genetic and developmental mechanisms underlying human lymphatic vascular diseases including primary lymphoedema and non-immune foetal hydrops. It’s an exciting time to be a developmental biologist!
My research program aims to understand how the lymphatic vasculature is constructed during development and how defects in this process result in human disease. We are particularly interested in defining the mechanisms by which endothelial cell identity is transcriptionally programmed and in understanding how lymphatic vessel valves are built.
I understand this is not your first collaboration with Natasha?
MF Tash and I have been long standing collaborators since 2005; at this time we were developing our own independent research programs focussed on embryonic lymphangiogenesis. Given our shared interest in the transcriptional control of lymphatic endothelial cell identity, it made immediate sense to join forces in order to develop this exciting new area of research. We have continued to collaborate since then; we regularly discuss ideas, have published many papers together and often apply together for grant funding to develop our synergistic approach. For this particular paper, our common interest in understanding how the lymphatic vessel network is assembled during development led to our teams making independent but complementary observations while studying the process of lymphangiogenesis in embryonic mouse skin. We therefore decided to join forces and combine research tools to push the investigation further. Our initial observations and discussions regarding clusters of lymphatic endothelial cells in embryonic skin date back to 2008!
And Cathy, how did you come to be involved with this project?
CP-T When I joined the Francois lab, Mat and I discussed several projects to work on. One of them was about the cellular origin of the dermal lymphatics. Mat and Natasha previously observed the presence of isolated lymphatic endothelial clusters in the skin that were not connected to the sprouting lymphatic vessel emerging from deeper tissues suggesting a potential distinct source of lymphatic progenitors. I was really excited about this observation and the biological question behind it so I embraced the challenge of investigating the cellular origin of dermal LECs.
Why has the origin of the lymphatic vasculature been so hard to pin down?
MF & NH Despite the advances that have been made to date, tools and technology have been limiting factors in completely defining the embryonic origins of lymphatic vessels. A major limitation in studying embryonic mouse development is the lack of live imaging capability, so even though we have a good understanding of morphogenetic events as observed in fixed tissues, we have not yet had the capacity to image lymphangiogenesis in real time in the developing embryo. We’re sure to be amazed once the technology is available to do this!
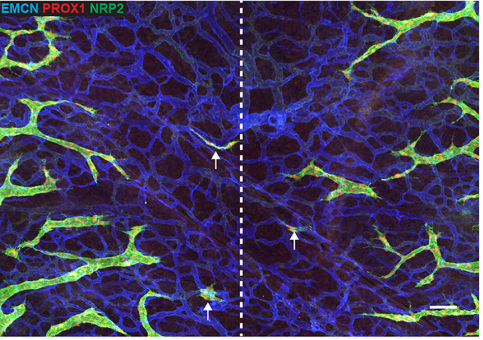
Whole-mount immunolabelled skin at 14.5dpc, from Fig. 1 in the paper
Can you give us the key results of the paper in a paragraph?
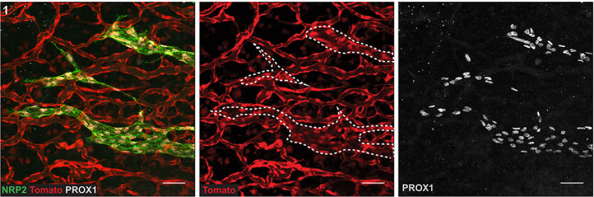
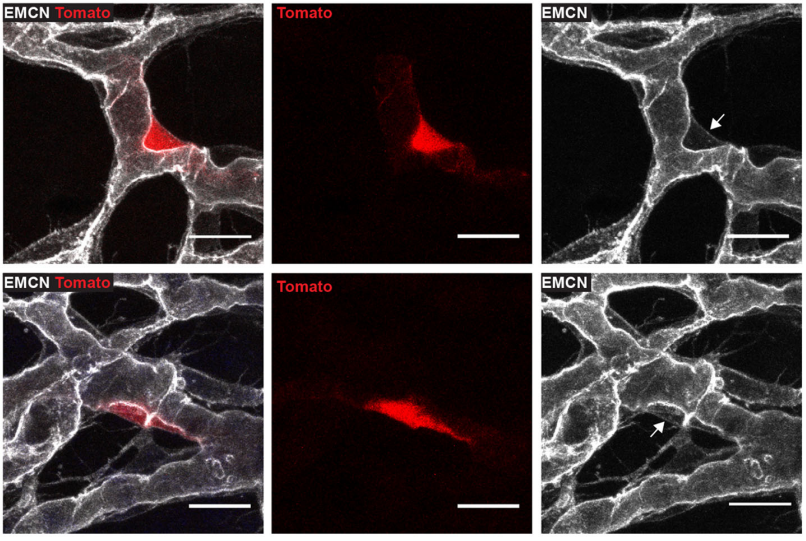
CP-T, MF& NH Our manuscript identifies a novel source of lymphatic endothelial progenitor cells that is employed during construction of the lymphatic vasculature in embryonic mouse skin. While it is well established that the majority of lymphatic endothelial cells originate from progenitor cells located in the cardinal and intersomitic veins, we identify here the dermal blood capillary plexus as a new source of progenitor cells. These progenitors are first observed as isolated Prox1-positive cells within the capillary bed in the midline region of dorsal embryonic skin around embryonic day 13 of development. These cells bud off from the capillary bed and proliferate to generate clusters of lymphatic endothelial cells (LECs) which sprout to meet up with LECs sprouting either from other clusters or the venous derived lymphatic plexus. Like the exit of LEC progenitors from veins, the exit of progenitors from the capillary bed is dependent on Ccbe1 and regulated by VEGF-C. Our data suggest that progenitor cell sources including the capillaries might contribute to the development of new lymphatic vessels both in other tissues and in pathological settings.
Tie2-Cre:tdTomato skin at 14.5dpc, from Fig. 2 in the paper
What might be upstream of Prox1 expression in the cells that become LEC progenitors, and why do only certain cells in the capillary plexus adopt the LEC fate?
MH & NF This is really fascinating question and one we don’t yet have an answer to. Endothelial cells within the capillary plexus appear relatively plastic and display markers of both arterial and venous identity. It is therefore likely that many of these cells have the potential to become LEC, however only a discrete population is programmed at particular times and locations in the skin. Whether this is due to spatially restricted cues that turn on Prox1, or to other cellular events remains to be established. We think that the emergence of LEC clusters might act as anchor points throughout the skin to guide the actively sprouting lymphatic plexus towards the midline. During this stage of development the embryo is growing rapidly, so it seems logical that a dual mechanism of vascular growth would allow the plexus to be built and connected more rapidly than one relying on sprouting growth alone.
What might be the functional relevance of building the lymphatic system in this way?
CP-T, MF& NH It is well established that the signalling pathways important for embryonic lymphangiogenesis are reactivated and face dysregulation in pathological states. This is the case during tumour induced lymphangiogenesis for instance, where the production of VEGF-C and VEGF-D by tumour cells promotes the growth of new lymphatic vessels within the tumour microenvironment and facilitates tumour metastasis. Our finding that the capillary plexus harbours lymphatic endothelial progenitor cells during embryogenesis raises the possibility that this, or a similar progenitor source might be called into play during pathological lymphangiogenesis, in contrast to new lymphatic vessels sprouting entirely from pre-existing lymphatics as current dogma suggests.
Prox1-CreERT2:tdTomato embryonic skin at 13.5dpc, from Fig. 4 in the paper
When doing the research, did you have any particular result or eureka moment that has stuck with you?
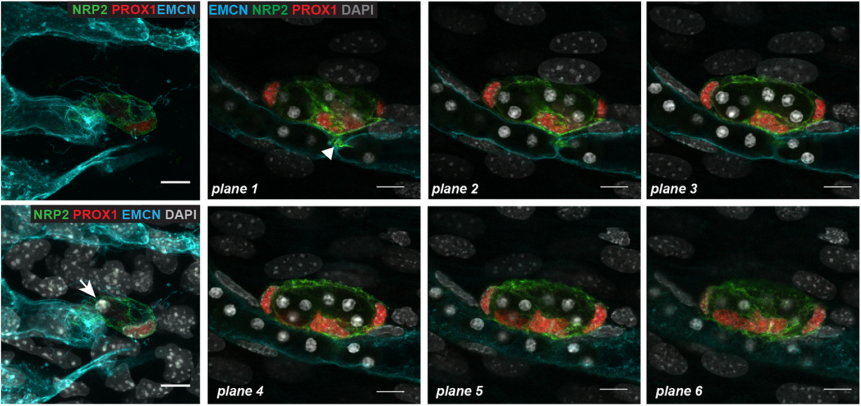
CP-T Probably one of my best moments while I was working on this project was when I studied the Ccbe1 mutant mouse embryos. We had this hypothesis that if the capillary plexus in the skin is a source of lymphatic endothelial cells, then LEC progenitors should be unable to leave the capillary plexus in signalling mutants that interfere with LEC mobility. We were able to capture images of some lymphatic progenitors “stuck” inside the capillary plexus, and this was a key advance to support our hypothesis. I remember screening a skin sample from a CCBE1 mutant embryo under the confocal microscope and then seeing it. This observation got me really excited!
And what about the flipside: any moments of frustration or despair?
CP-T What was really challenging about this project was the fact that we were dealing with a rare event so it was hard to study it. As every scientist I guess, I had many moments of frustration and despair, but that make the “eureka moments” event more pleasant!
LEC progenitors in the blood vascular capiliary plexus, from Fig. 5 in the paper
What next for you Cathy – I hear you’ve moved to a new lab?
CP-T Yes, I have moved back to France where I am working now at the Curie Institute. I will still work on the vascular system but related to brain tumour biology.
Where will this work take the Harvey and Francois labs?
MF At present, the lab is focussing on new genes that we have identified as key regulators of lymphangiogenesis. We will use the identification and characterisation of LEC clusters as a readout to characterise subtle phenotypic responses.
NH We would love to build our imaging capacity! I’m fascinated at the prospect of watching lymphatic vessels grow in real time.
Finally, let’s move outside the lab – what do you like to do in your spare time?
MF At the moment I am doing under-water rugby, lots of fun.
NH Weekends involve time at the beach or the park with my kids, together with enjoying the fantastic food and wine that Adelaide has to offer. My early morning boxing class is as good for my brain and my energy levels as it is for my physical health!
CP-T Back in my home country just recently, I am enjoying spending time with my family and old friends.
A postdoctoral position is available in the group of Taija Mäkinen at the Department of Immunology, Genetics and Pathology (IGP) at Uppsala University in Sweden.
IGP provides a well-developed infrastructure for advanced molecular sciences. Several prominent vascular biology groups have recently been recruited to the department, creating an inspiring frontline environment for vascular research.
Research in the lab focuses on discovering fundamental mechanisms controlling vascular morphogenesis and functions. The selected candidates will focus on studying organ-specific mechanisms of lymphatic vessel formation by utilising and developing advanced mouse genetic tools and cell and molecular biology techniques (e.g. single cell RNA sequencing, flow cytometry, confocal, light-sheet and super-resolution microscopy). Available projects include: 1) identification and functional characterisation of vascular-bed specific genes, and 2) functional characterisation of tissue-specific lymphatic endothelial progenitor cells.
Qualifications needed:
We are looking for a highly motivated individual with a PhD in a relevant area and a proven track record of successful scientific work. Strong background in molecular/cell biology, mouse genetics and/or imaging is required.
Please send you CV together with the names of three references and a short description of yourself and the motivation to join the group to: taija.makinen[at]igp.uu.se
In this post we report the backstories behind our recently published paper. It was an enjoyable research adventure driven by discussions, readings, exciting experiments and unexpected discoveries. As a result, we described a novel molecular mechanism underpinning stem cell and progenitor maintenance during development. Here is the sequence of the main events that inspired us in making this story.
The foundations; it’s good to have a side project
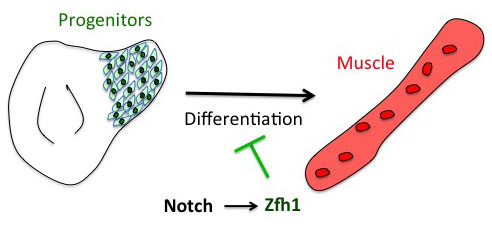
The project has started from the following basic developmental biology question: How are the Drosophila flight muscle progenitors (MPs) kept undifferentiated throughout much of the animal development? To solve this question, we initially embarked on a genome wide approach to define their unique characters. But after many rounds of dissections and ChIPs, the ChIP-seq results that came back (over the Christmas break!) they were basically meaningless and we were lucky that we had a side project in parallel with one of our favourite genes. Our enthusiasm for the zinc finger homeodomain transcription factor Zfh1/ZEB came from its identification in a previous genome wide screen for direct Notch targets in different Drosophila cell lines, including Drosophila DmD8 cells (MP-like cells) (Krejcí et al., 2009). Also, it was well established that Zfh1, as well as its mammalian homologues ZEB1/2, had roles in maintaining stem cells. Together these elements prompted us to investigate the role of zfh1 in MPs maintenance and we ended up showing that indeed Notch pathway activates directly zfh1 expression in vivo in the MPs and this cascade is required to keep them undifferentiated. This regulatory mechanism set the basic foundations for our subsequent work.
Figure 1. Zfh1 expression is directly activated by Notch to maintain muscle progenitors and prevents premature differentiation.
How the microRNA miR-8 entered the scene
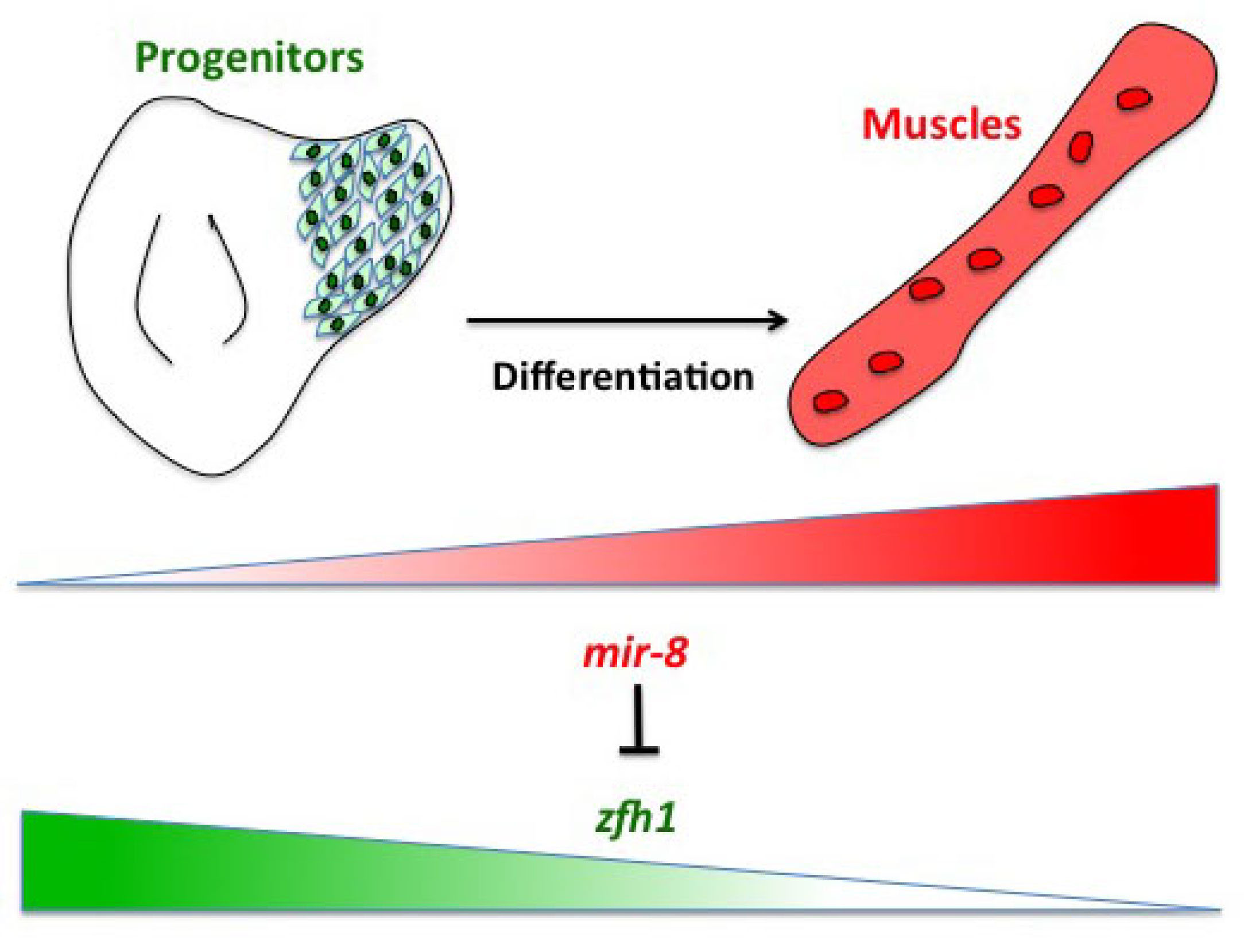
Having described the Notch/Zfh1 regulation, we realised that one important question was how the progenitors cells get rid of Zfh1 in order to differentiate. This led us to a paper from Maria Dominguez lab, which linked zfh1 to the micro RNA miR-8 in the intestinal stem cells (Antonello ZA et al., 2015). Indeed we discovered from there that miR-8/zfh1 regulation orchestrates many developmental process in either Drosophila or vertebrates so it seemed natural to check whether it plays any role in the muscle differentiation program … as turns out to be the case. We found that miR-8 down regulated zfh1 in the MPs to facilitate the transition into functional flight muscles. Thus, we added miR-8 to the Notch/Zfh1 regulation and started to fill the gaps, strengthen the data and think about how to complete the story.
Figure2. The micro RNA miR-8/miR-200 antagonizes zfh1 to promote the muscle differentiation.
Satellite cells do exist in Drosophila ?
So far the focus was on the muscle progenitors prior to the muscle differentiation. But might some of these cells persist in the adult? At that time, I had no idea of how to do antibody stainings on adult muscles. Thankfully, Hannah Green and Juanjo Perez-Moreno kindly agreed to show me how to dissect and stain adults and pupal muscles. Excitingly there were Zfh1-positive cells associated with the adult muscles. But this did not immediately mean they were persistent muscle precurors…. the lab (aka my PI!) was not convinced! They pointed out that blood cells were also likely to be cruising around and that the zfh1 enhancer I used overlapped with a powerful blood cell enhancer identified by Alex Stark group (Zabidi M A et al., 2015). So next, I was tasked with double staining with blood cell markers to rule out this possibility. After a few trials I could finally show that indeed there was a population of cells expressing Zfh1 that were distinct from the plasmatocyte blood cells. So it seemed I was right! There is a persistent muscle progenitor cell population in adult flies similar to the satellite cells in mammals.
Very excitingly, another group had reached the same conclusions. In early 2016, we came across an intriguing pre-print paper from VijayRaghavan’s Lab describing muscle Satellite Cells (SCs) in adult flies (Now published as Chaturvedi D et al., 2017). In this manuscript the authors described and characterised a SC population, resident beneath the basal lamina of the adult muscles, and had found that they are lineal descendant of the larval MPs. So we carried out some similar lineage analysis to confirm that indeed the Zfh1-expressing cells had the same properties as their satellite cells. Furthermore, we could show, using one specific zfh1 enhancer to drive expression of a pro-apoptotic gene, that their elimination affected muscle homoestasis. Good news never comes alone: while we were writing our paper, an updated version of the VijayRaghavan Lab SC pre-print was released, they included new data with Zfh1 stainings in the SCs. It was reassuring to see that our data converged together!
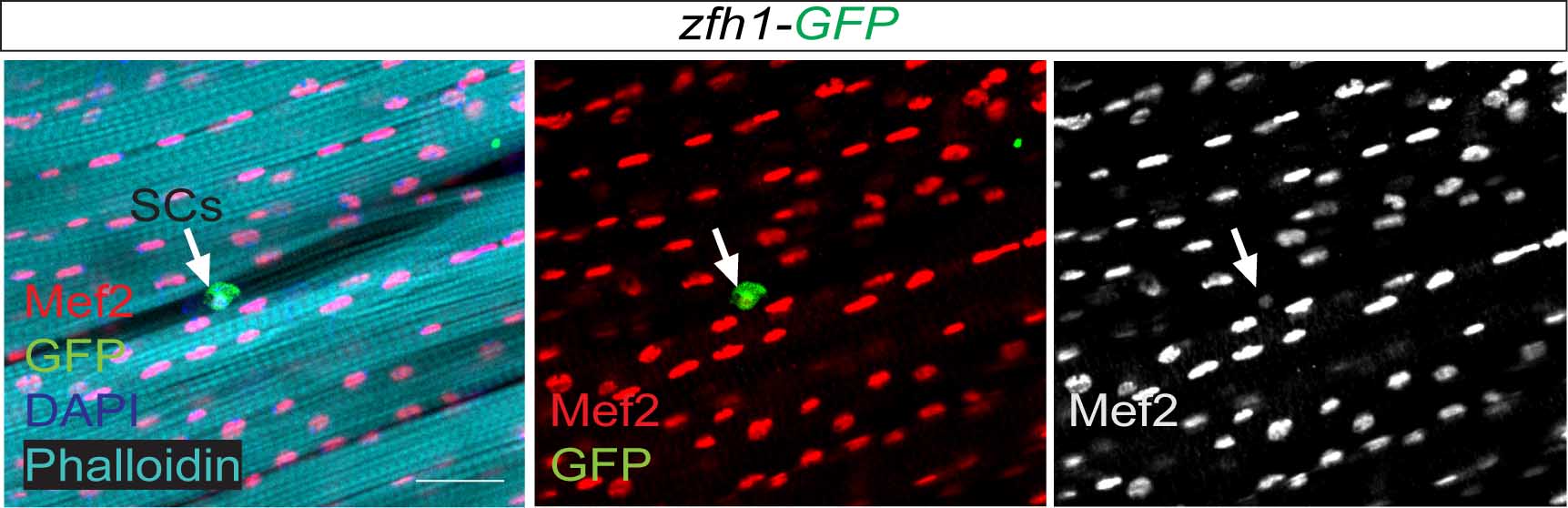
Figure 3. From Figure 2 of the paper. Zfh1 expression (Green) indicates the existence of satellite cells (SCs; arrows) associated with the muscle fibres (Phalloidin (Cyan)) and expressing low level of the myogenic transcription factor Mef2 (Red).
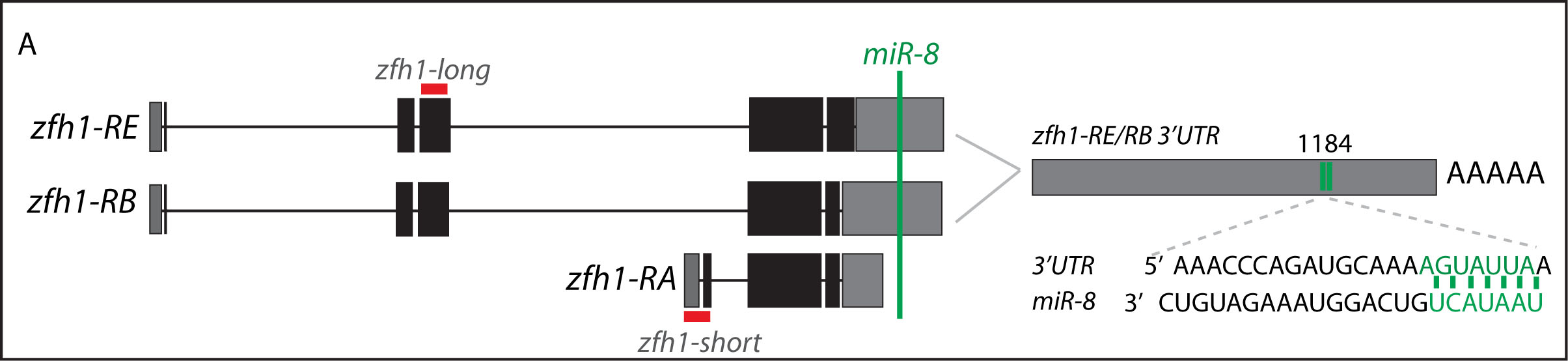
Short vs Long zfh1 isoforms: Why does length matter?
The discovery of these Zfh1-SCs in the adult flies placed us in an unprecedented position to address the exiting question of how such SCs escape the differentiation program during muscle development. Indeed, It is quite intriguing that this small subset of MPs behaves differently from the rest of the main pool of myoblasts that normally initiate the muscle differentiation program. In a discussion after my lab meeting presentation we ended up looking at the different zfh1 transcripts on Flybase (The online Drosophila database) and realised that there is a second zfh1 isoform, which is characterised by a shorter 3’UTR. Importantly, and unlike zfh1-long isoforms, zfh1-short isoform lacks the miR-8 seed site so it could be invisible to miR-8 detection. We thought this could be an exciting mechanism to protect the SCs and so we set out to test the hypothesis that a switch in zfh1 isoforms could be the trick to prevent SCs from differentiation. We quickly designed RNA-probes and checked the expression profile (by smFISH) of these isoforms during development. The results were striking and consistent with the initial hypothesis, since we found that zfh1-short is accumulated to high level specifically in the SCs. A series of experiments followed up and the manuscript started to have a pretty nice shape especially after the first round of elife reviewing process where the reviewers gave us many valuable comments and suggestions.
Figure 4. From figure 7 of the paper. Schematic representation of zfh1 isoforms. zfh1-short (zfh1-RA) is initiated from a different transcription start site and has shorter 3’UTR that lacks the target site for miR-8 (Green; Antonello et al., 2015) present in zfh1-long isoforms (zfh1-RB,zfh1-RE); the position of the miR-8 seed sites in zfh1-long 3’ UTR are depicted. Non-coding exons and coding exons are depicted by grey and black boxes respectively, red lines indicate the probes used for FISH experiments.
Conclusion
Our voyage ended in a very different place than we started it! Key to its success was challenging ourselves to go beyond the simple results and seizing the ideas from the literature. Bringing these together meant that we could delve into the mechanisms, but only with the additional inspiration from the data in Flybase did we make the final leap, which shows how important these resources are! In the end, we discovered a mechanism that explains how stem cells can escape the differentiation program and survive until the adult stage where they contribute to tissue homeostasis. At the root of it is a switch in RNA isoforms to produce RNAs that are insensitive to miRs. We believe that this is a powerful mechanism that may be of widespread significance.
 The protozoa with endosymbiotic algae are found in highly-productive ponds where oxygen levels are depleted; having an association with an oxygen-producing machine (the microscopic algae) seems the perfect arrangement. An example of this was the rare ciliated protozoan Loxodes rostrum (pictured) – an extremely unusual ciliate in many ways. Firstly, it has seldom ever been recorded (only in Germany and the UK, so far); secondly, it is a ciliate that harbours several different species of microscopic algae (most ciliate-algae associations are with one species only of the genus Chlorella). Unfortunately, the pond in Dorset dried out before we could finish our fascinating investigations on this unique species.
The protozoa with endosymbiotic algae are found in highly-productive ponds where oxygen levels are depleted; having an association with an oxygen-producing machine (the microscopic algae) seems the perfect arrangement. An example of this was the rare ciliated protozoan Loxodes rostrum (pictured) – an extremely unusual ciliate in many ways. Firstly, it has seldom ever been recorded (only in Germany and the UK, so far); secondly, it is a ciliate that harbours several different species of microscopic algae (most ciliate-algae associations are with one species only of the genus Chlorella). Unfortunately, the pond in Dorset dried out before we could finish our fascinating investigations on this unique species. (2 votes)
(2 votes)
 (No Ratings Yet)
(No Ratings Yet)


 During that time the key discovery from my work identified the transcription factor SOX18 as a molecular switch that induces the lymphatic differentiation program by turning on Prox1 expression in pre-existing veins.
During that time the key discovery from my work identified the transcription factor SOX18 as a molecular switch that induces the lymphatic differentiation program by turning on Prox1 expression in pre-existing veins. 

 (4 votes)
(4 votes)