Applications are invited for a four year PhD studentship funded by The Royal Society to conduct research under the supervision of Dr Amanda Sferruzzi-Perri at the Department of Physiology, Development and Neuroscience.
Title: Role of placental endocrine malfunction in the programming of disease in offspring
Closing date: 17th June 2018
For more details about the project and application process please see the following webpage:
DanStem is looking for a postdoctoral candidate with a strong cell biological and cell signaling background in directed differentiation of human pluripotent stem cell biology.
The candidate is expected to work on the engineering of human pluripotent stem cell-derived islet of Langerhans-like aggregates with functional properties close to human islets. The functionality and therapeutic potential of the aggregates will be tested in vitro and in vivo in mice.
The goal of this project is to engineer therapeutically active islet-like aggregates for future cell therapy phase 1 trials in Type 1 Diabetes (T1D).
The candidate is expected to use state-of-the-art genetics, molecular and cell biological, and tissue engineering experimental strategies. The candidate will work together with a dedicated team of scientists and technicians who together will tackle bottle-necks towards implementing the phase 1 clinical trials in T1D.
Qualifications:
The candidate is required to hold a PhD degree in pluripotent stem cell/developmental biology. A few years of postdoctoral experience in the same areas is a merit. The candidate should also have hands on experience in human pluripotent stem cell maintenance and differentiation, 3D culture of pluripotent stem cells, various cell and molecular biological methods, flow cytometry and live-cell imaging. Experience in differentiation towards pancreatic lineages is a merit. Finally, we are looking for applicants with a good record of peer reviewed scientific publications, grant writing skills and an interest in team work.
About our center:
The Novo Nordisk Foundation Center for Stem Cell Biology – DanStem – addresses basic research questions in stem cell and developmental biology and translates results from basic research into new strategies and targets for the development of new therapies for diabetes and cancer. DanStem was established as a result of a series of international recruitments coupled with internationally recognized research groups focused on insulin producing beta cells and cancer research already located at the University of Copenhagen.
The position is for 2 years with a possible extension. The employment is planned to start as soon as possible or upon agreement.
The terms of employment are set according to the Agreement between the Ministry of Finance and The Danish Confederation of Professional Associations or other relevant professional organization. The position will be at the level of postdoctoral fellow and the basic salary according to seniority is 32.700-34.400 DKK/month. A supplement could be negotiated, dependent on the candidate´s experiences and qualifications. In addition, a monthly contribution of 17.1% of the salary is paid into a pension fund.
Applicants recruited from abroad may be eligible for a special researcher taxation scheme. In all cases, the ability to perform the job will be the primary consideration, and thus we encourage all – regardless of their personal background and status – to apply.
The application must be submitted online and in English, by clicking on “Apply online” below.
The closing date for applications is July 15th 2018 at 23.59pm.
The application must include:
Cover letter detailing the basis on which the applicant scientific qualifications meet the requirements for this position.
Curriculum vitae.
List of references (full address, incl. email and phone number)
Diplomas – all relevant certificates.
List of publications.
The further process:
After the expiry of the deadline for applications, the authorized recruitment manager selects applicants for assessment on the advice of the Appointments Committee. All applicants are then immediately notified whether their application has been passed for assessment by an expert assessment committee. Selected applicants are notified of the composition of the committee and each applicant has the opportunity to comment on his/her assessment. You may read about the recruitment process on
DanStem and The University of Copenhagen wish to reflect the diversity of society and welcome applications from all qualified candidates, regardless of personal background
The Laboratory of Early Mammalian Developmental Biology (LEMDB), University of South Bohemia, České Budějovice, Czech Republic (Alexander W. Bruce group).
The LEMDB is currently recruiting both Masters (Mgr) & Ph.D. level studentships to research projects related to cell-fate choice.
Research projects offered in relation to p38 mitogen-activated-kinase’s role in primitive endoderm/ PrE development, the regulation of spatial allocation of cells relevant to first differential cell-fates i.e. outer trophectoderm/TE & inner-cell-mass/ ICM and others…
University provided stipend guaranteed for 4-years (plus grant based salaries and performance related bonuses)
Please send your CV and covering letters to lab head Alexander W. Bruce at: awbruce@prf.jcu.cz
In September, Development is hosting the third of its highly successful series of meetings focusing on human developmental biology. Held in the Wotton House estate near Dorking in Surrey and organised by Paola Arlotta, Ali Brivanlou, Olivier Pourquiéand Jason Spence, the meeting will cover the latest developments and future prospects for this rapidly evolving field.
A postdoctoral position is available starting in October 2018 at the Developmental Biology Institute of Marseille, France, located on the Luminy campus, in the group of Vincent Bertrand. The position is funded for 3 years.
The acquisition and maintenance of the differentiated state of a neuron are achieved by terminal transcription factors that are expressed throughout the life of the neurons. However, gene expression is often noisy, presenting a high stochastic variability. The objective of this project is to determine how the identity of a neuron can be established and maintained throughout the life of the animal in a reliable manner despite this variability. This question will be addressed using C. elegans, an excellent system to analyze the robustness of neuronal development. The project will combine recent genome engineering methods (CRISPR), quantitative in vivo imaging and single molecule FISH.
We are seeking a highly motivated candidate with a PhD in molecular biology, genetics, cell biology, developmental biology or neurobiology. The working language in the laboratory is English.
A letter of motivation, a CV and the names of two referees should be sent to Vincent Bertrand (vincent.bertrand@univ-amu.fr).
A postdoctoral position is available in the laboratory of Dr. Sophie Astrof at Thomas Jefferson University to study roles of cell-extracellular matrix (ECM) interactions in cardiovascular development and congenital heart disease. We have recently discovered that progenitors within the second heart field (SHF) give rise to endothelial cells composing pharyngeal arch arteries. Projects in the lab focus on the role of ECM in regulating the development of SHF-derived progenitors into endothelial cells and their morphogenesis into blood vessels. The successful candidate will combine genetic manipulation, embryology, cell biology, and confocal imaging to study molecular mechanisms of micro-environmental sensing during vascular development.
Astrof laboratory is a part of a modern and well-equipped Center for Translational Medicine at Jefferson Medical College (http://www.jefferson.edu/university/research/researcher/researcher-faculty/astrof-laboratory.html) located in the heart of Philadelphia.
To apply, please send a letter of interest detailing your expertise, CV and names and contact information of three references to sophie.astrof@gmail.com
Short CLE peptides regulate a wide variety of processes during plant development. In the developing root, the receptors and co-receptors for CLEs have remained largely unclear, as have the relationships between different CLEs and different (co-)receptors. A new paper in Development addresses this problem by reporting a new receptor kinase involved in CLE signaling. We caught up with lead author Pauline Anne and her PI Christian Hardtke, Professor in the Department of Plant Molecular Biology at University of Lausanne, to find out more.
Christian, can you give us your scientific biography and the questions your lab is trying to answer?
Out of an early interest in genetics, I studied Biology at the University of Munich. I became particularly fascinated by Developmental Biology when I learned about the then still recent isolation of the Bicoid morphogen. At that point I was determined to work with Drosophila. Incidentally, Thomas Berleth, who had cloned Bicoid a few years earlier, worked with Gerd Juergens in the Genetics department at that time. They were recruiting an M.Sc. student to investigate the “Arabidopsis embryo patterning mutant monopteros”. I only realised during the interview that Arabidopsis was not some short germ-band insect, but an upcoming plant model organism. Maybe my judgement was clouded by the dense pipe smoke in Gerd’s office, but I committed to the project, which also became my Ph.D. In hindsight, it was a stroke of luck, I enjoy working with plants ever since.
Four years later I had cloned MONOPTEROS and walked enough along chromosomes. Thus, after briefly wrapping up my Ph.D. work at the University of Toronto with Thomas, I started my postdoc as an HFSP fellow in the lab of Xing-Wang Deng at Yale University, to learn biochemistry. It was an exciting time as the lab discovered the mechanistic basis of the COP9-COP1 signalosome-ubiquitin ligase system in Arabidopsis photomorphogenesis, and its operational conservation but differential output in distant eukaryotes. When I started my own lab at McGill University however, I turned to root system architecture with the aim to analyse its intraspecific natural genetic variation. Since then, we isolated several QTL alleles. Because I like to explore the molecular foundations of their developmental phenotypes, my lab’s focus has shifted in the years after I moved to the University of Lausanne. Nowadays we are mainly interested in deciphering molecular pathways that govern the patterned differentiation of plant vascular tissues, and in identifying the non-cell autonomous signals that are emitted from phloem vasculature to organize surrounding tissues.
And Pauline, how did you come to work in Christian’s lab and be involved with this project?
Christian and I share common interests. In my PhD, I characterised the phloem regulator OCTOPUS (OPS) and its interaction with the brassinosteroid phytohormone pathway. I wanted to continue working on phloem differentiation, and Christian is a leader in that field. We discussed possibilities, I wanted to continue exploring signalling pathways, so he proposed to analyse CLE26 sensing, which we suspected to play a role in phloem differentiation.
When did your lab first get interested in CLE peptide signalling in the root?
CH When we got stuck in our analysis of BREVIS RADIX (BRX), another positive regulator of phloem formation. Stephen Depuydt, a postdoc in my lab at the time, proposed to screen for suppressors of the phloem defects in brx mutants. Although I was rather sceptical about the prospect of healing the strong brx phenotype by second site mutation.
PA But it worked! Loss-of-function in BARELY ANY MERISTEM 3 (BAM3), a receptor kinase, was isolated as a perfect brx suppressor from this screen. They later found that BAM3 is the receptor for the phloem-specific CLE45 peptide, pointing to a role of autocrine CLE peptide signalling in phloem differentiation.
CLERK expression pattern in seedlings from Fig. 3 in the paper
Can you give us the key results of the paper in a paragraph?
PA We isolated CLERK as a genetic component of CLE26 sensing in the root, but it is also required for perception of other CLE peptides. Moreover, CLERK encodes a co-receptor protein that acts in the phloem and is necessary to prevent premature protophloem specification.
CH Also, CLERK acts genetically in parallel to described CLE sensing pathways in the protophloem, corroborating the redundancy we suspected at the outset.
You have lots of CLE peptides, lots of receptors, lots of co-receptors, and lots of promiscuous receptor-ligand and receptor-co-receptor interactions. How does this system achieve any degree of specificity?
PA Although many exogenously applied CLE peptides suppress root protophloem differentiation, only few, like CLE45 or CLE26, are actually expressed in root development. Recent studies by colleagues suggest that some CLEs are induced by specific environmental stimuli, to shut down root growth in unfavourable conditions. So part of the answer probably lies in specific ligand expression.
CH …and in specific receptor and co-receptor expression. It is also noteworthy that some receptors are not only more promiscuous with respect to ligands than others, but that ligand binding affinities can vary by an order of magnitude. So even in presence of multiple receptor-coreceptor pairs, ligand interaction can likely trigger vastly different signal outputs. It will be a major challenge to map these interactions in time and space, also because cross-regulation between CLE-sensing receptors has been observed.
How might CLERK help you in future analysis of the CLE sensing pathway?
CH One surprising discovery was that in brx or ops mutants, CLERK is not required for CLE26 or CLE45 sensing. We can therefore perform a sensitized screen for CLE resistance, for example in a brx clerk double mutant background. This should enable us to home in on the CLE26 receptor, as well as receptors for other CLEs.
PA The system is more and more complex, and genetics does not always give the expected answers. But it looks like it is less complex in the root than in the shoot. For example, while our paper was under review, colleagues found that CLERK also plays a role in the shoot. However, there its action is masked by redundancy between four related co-receptors.
CLERK’s effects on CVP2 expression, from Fig. 5 in the paper
When doing the research, did you have any particular result or eureka moment that has stuck with you?
PA Yes, the day I received the whole genome sequencing results of our original clerk mutant! I was on vacation in Italy. As you guess, I could not wait to be back in the lab to analyse the results…and At2g23950 popped up, a receptor-like kinase. This was the best candidate I could expect at that moment.
And what about the flipside: any moments of frustration or despair?
PA Of course! Even if the project went well, there are more instances of frustration in research life than eureka moments. I think my main frustration was the very weak expression level of CLERK in the root tip. But on the upside, this forced me to hone my confocal microscopy skills…
What next for you after this paper?
PA In the short term, I will focus on the characterization of other candidates isolated from the screen. I am most interested in a downstream CLE signalling component that I isolated recently, a prey that the lab has long been after. In the long term, I hope to turn my projects into more publications, to be competitive in the academic job market.
And where will this work take the Hardtke lab?
CH It will force us deeper into investigating the mechanics of CLE perception. One unexpected result contributed by our collaborators, Michael Hothorn’s lab, was the observation that the CLERK ectodomain cannot interact with the ectodomains of a few known bona fide CLE receptors, in the presence of ligand. If CLERK is indeed a generic CLE co-receptor, which is a parsimonious assumption, then this indicates that something is missing to make the system work biochemically in isolation. Maybe the sensitized screen will deliver this piece of the puzzle.
Finally, let’s move outside the lab – what do you like to do in your spare time?
CH Depends on the time of year. In the winter, I take every opportunity to go skiing. Luckily, from Lausanne it only takes about one hour to reach the bottom of some of the most spectacular ski slopes on the planet. In the summer, I like to attend a random selection of sports events. In general, I like to watch people perform feats that are impossible for myself, like swimming 100 m in less than a minute…
PA …I am a mom of two young kids, 3 years and one and a half years old. I spend all my free time with them and I really enjoy it. I love to read books with them. But sometimes I also have to fold up to fit in a cardboard hut, to escape wolves or other monsters. Sometimes it is more tiring to be a mom than to be in the lab. But my favourite moments are with the kids outside, observing nature, playing with its treasures, showing them birds, insects and, of course, plants!
We are seeking to recruit a research technician to work in the research group of Dr Vikas Trivedi at the newly established European Molecular Biology Laboratory (EMBL) Barcelona. We are situated in the Barcelona Biomedical Research Park (PRBB), one of the largest infrastructures in Southern Europe dedicated to translational research and which is located on the Mediterranean coast. Work in the group focusses on studying self-organization in multi-cellular systems, through comparative study of embryos and embryonic organoids, using both zebrafish and mouse models.
The position is available for a period of 3 years, in the first instance and can be renewed, depending on circumstances at the time of the review.
The successful candidate will be responsible for providing technical support for all projects running in the laboratory and general lab management.
Tasks
• Cell culture and cell line maintenance of mammalian (primarily mouse) embryonic cells
• Maintenance of zebrafish colonies, setting up crosses
• Zebrafish embryology techniques (cell transplantation, dissection, zygotic injections etc.)
• Basic molecular biology (PCRs, in vitro transcription, cloning, Western Blots, in situ hybridization and immunohistochemistry)
• Overseeing day to day operations, general organization of the laboratory such as administration of several databases (plasmids, antibodies, cell and fish lines, etc)
• Maintaining supplies (preparing and ordering reagents), servicing equipment, developing new protocols and training new laboratory members
• Setting up routine imaging and cell sorting experiments and sample preparation for the same.
• In addition to the technical support, the successful candidate will be encouraged to carry out their own research project within available capacity
Qualifications and Experience
The successful candidate must have:
• a Bachelors or Master’s degree in biology or related life science field
• one or more years of experience with cell/tissue culture techniques and cell sorting
• background in basic molecular biology techniques
• experience with zebrafish research is ideal but not necessary
• basic understanding of imaging (e.g widefield microscopes) is desirable but not necessary. Any prior experience with standard packages such as ImageJ /FIJI would be an advantage
• strong interpersonal and verbal communication skills to interact with internal/external contacts at all levels in a friendly and professional manner. You will be a team worker in an international and collaborative environment.
• openness to learn new techniques and work independently
• well organized and reliable
• fluency in English; knowledge of Spanish will be a plus
The successful candidate will be expected to take over routine imaging and analyses over time. However, any skills needed in the process will be taught and the candidate is expected to have proactiveness and enthusiasm toward new techniques.
Further details about the application can be found here
Additional Information
The remuneration package comprises from a competitive salary, a comprehensive pension scheme, medical, educational and other social benefits, as well as financial support for relocation and installation, including your family. Further details can be found here
Visualizations for comparing datasets is a topic in all my data viz classes. Current solutions for comparing 2,3, 4 and more datasets are diverse and some are controversial. A one-fits-all solution does not exist, but there are well-working solutions, and some that should be avoided.
1-3 datasets
Comparing two or three datasets works well in Venn diagrams. Most people learn them in school, and if not they are intuitive*. Each dataset is shown as a circle and they are arranged such that all overlaps are shown. Done.
More datasets
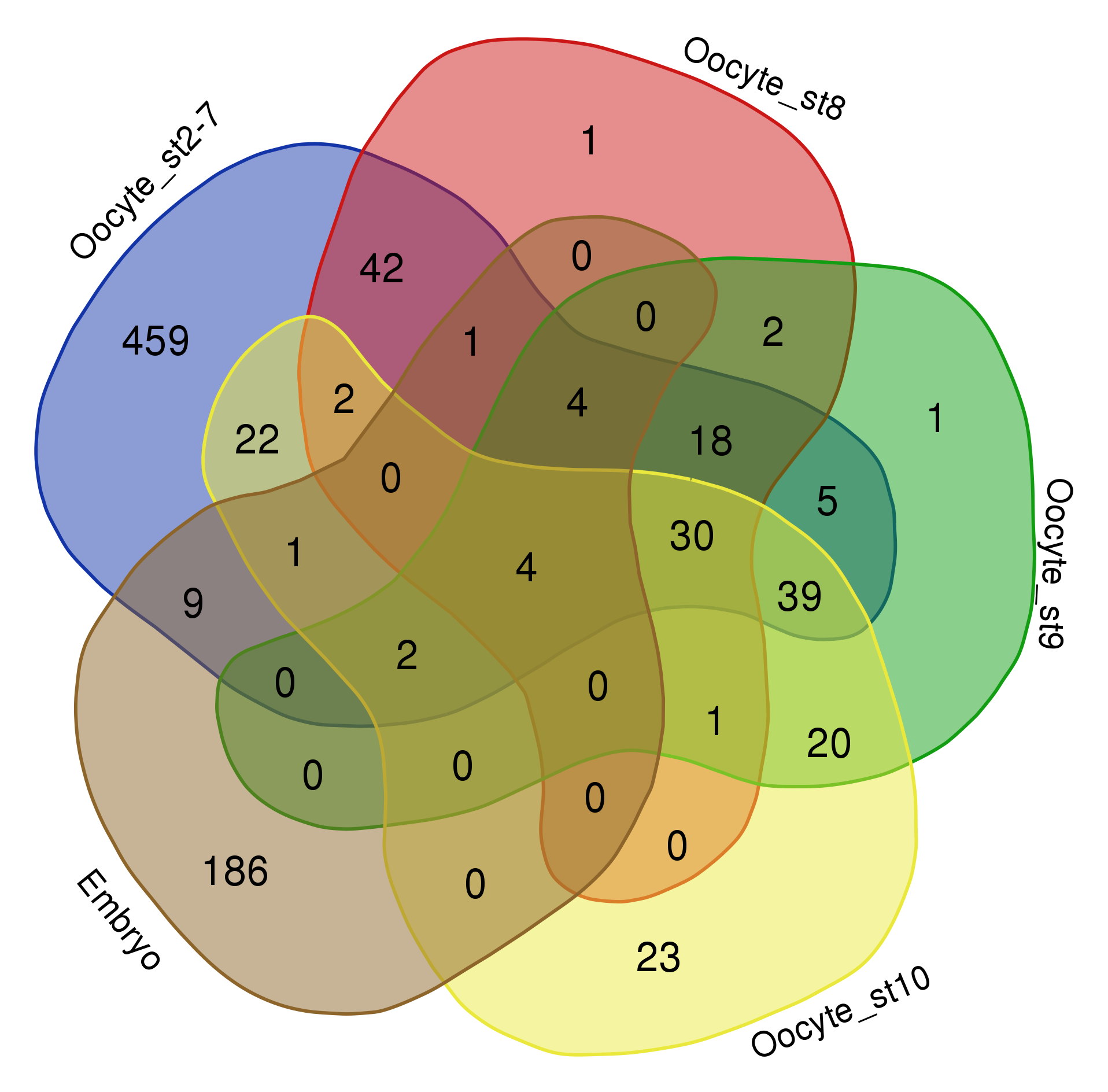
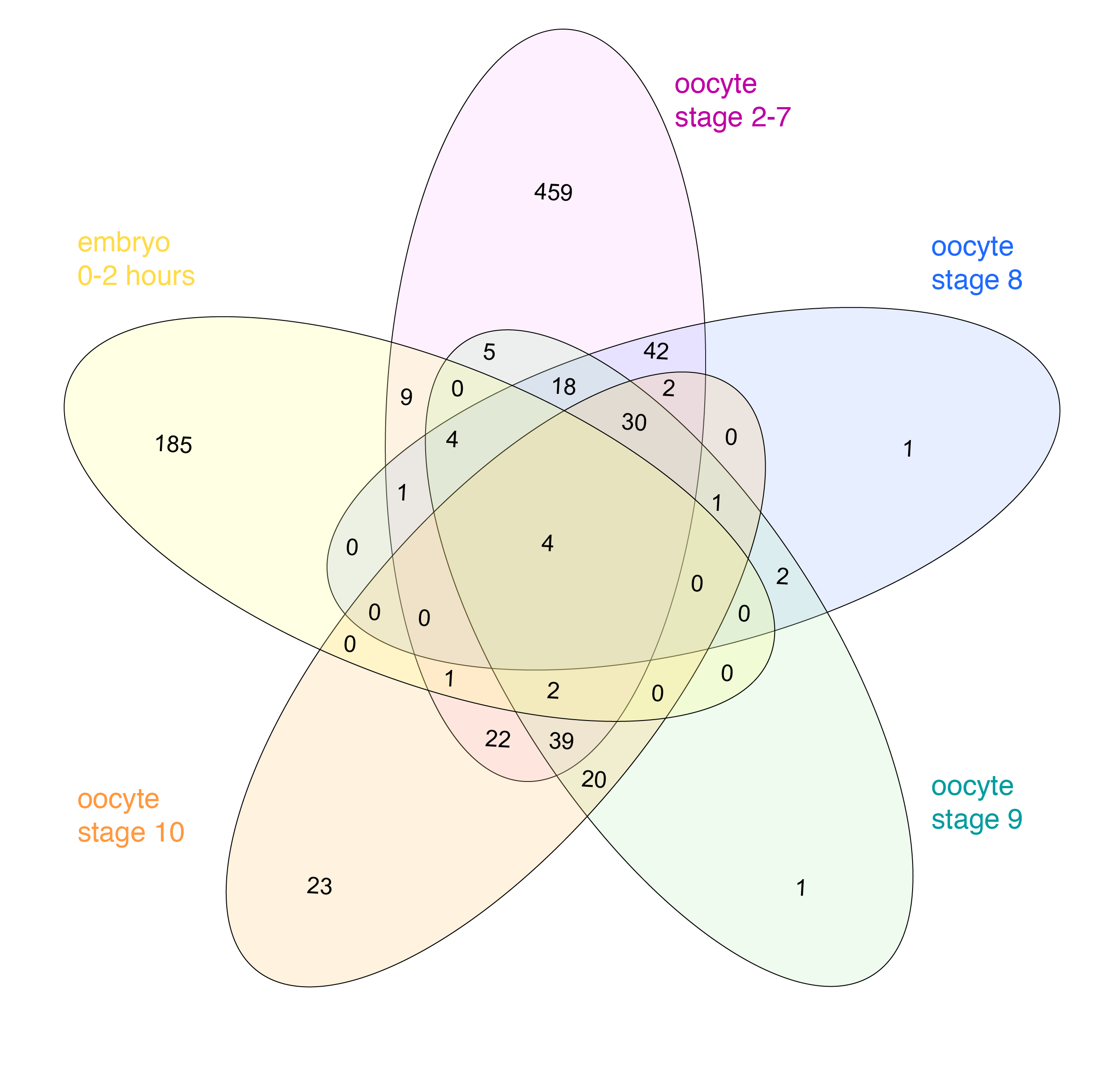
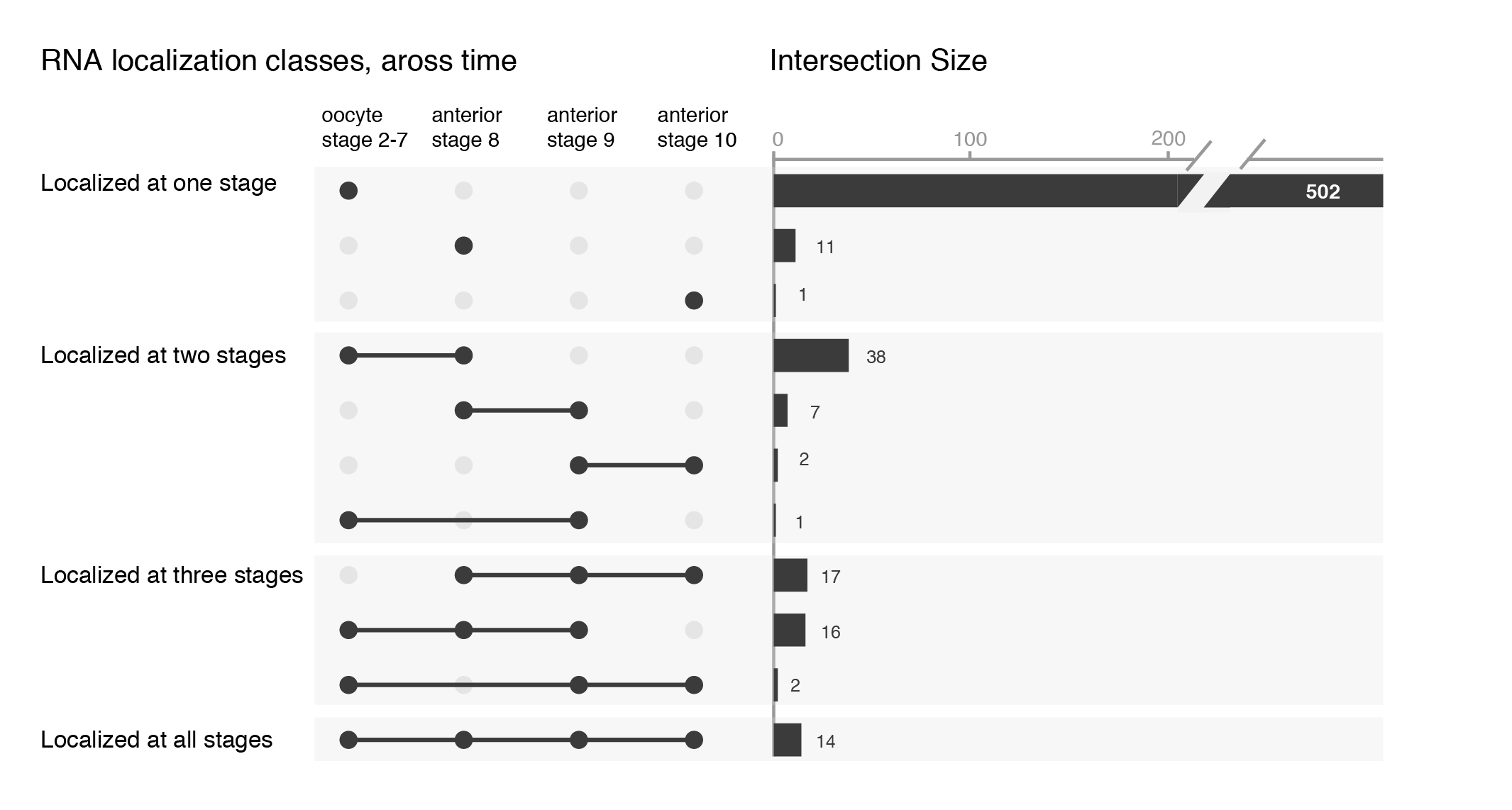
Things get problematic when comparing more than three datasets. Mathematically, it is not possible to show all overlaps of four or more datasets with circles. One possibility is to leave out some overlaps, as is often done in Euler diagrams. In the example below the overlap between “oocyte stage 2-7” and “oocyte stage 9” is for instance not visualized (RNAs localized in the oocytes across development, see publication). I find it however confusing when data is left out and sometimes “no overlap” is an important information itself.
Datasets of RNAs subcellularly localized in Drosophila oocytes.
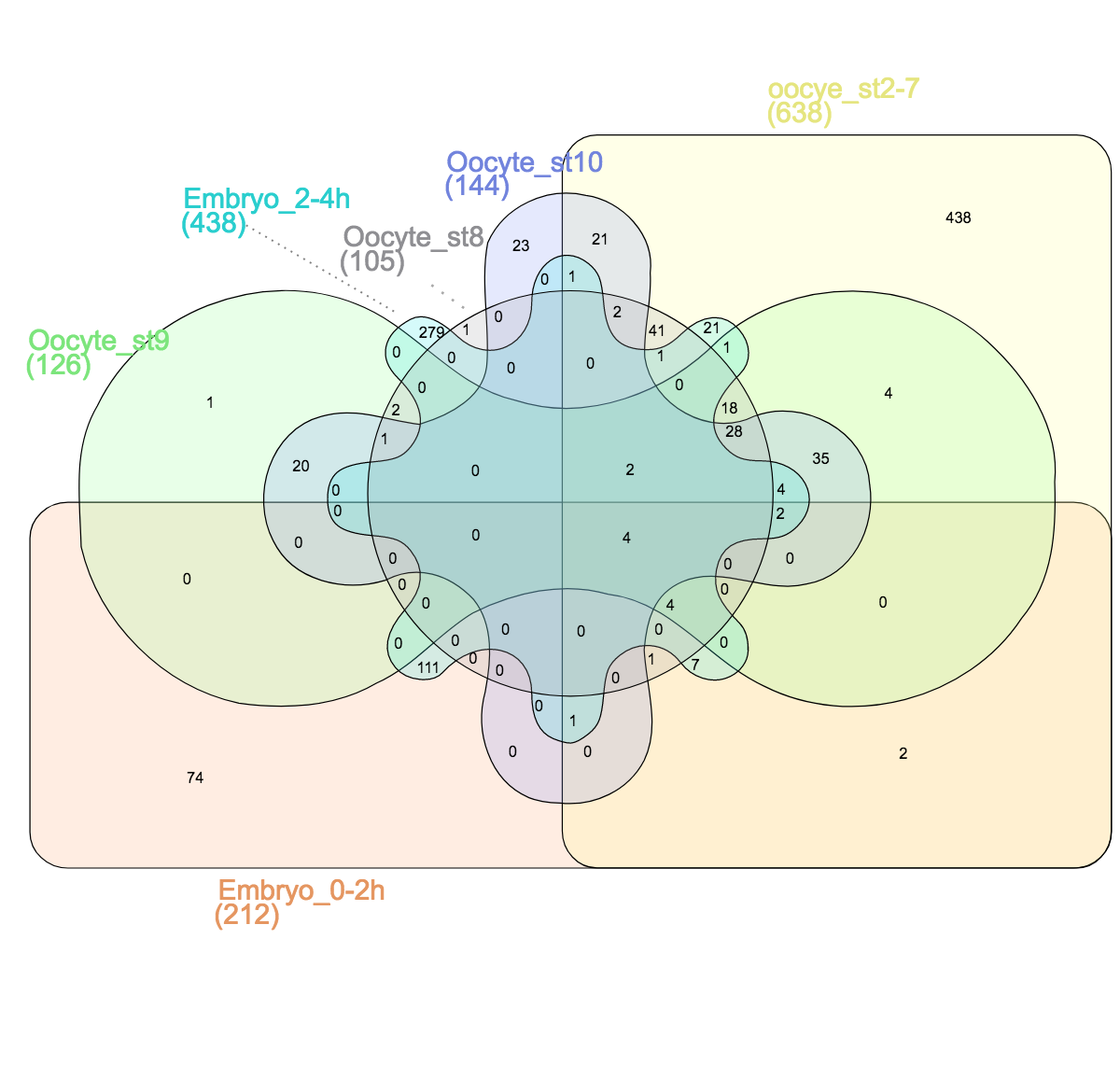
Venn himself devised the diagram comparing four and more datasets by switching from a circles to ellipses. Branko Grünbaum developed the ellipsoid representation further for comparison of five dataset by . Their strategies are used by the online tool Draw Venn (Yves Vandepeer, Univ of Gent) where you can make Venn plots by simply uploading your data there. A variation is used by Heberle et al here (publication). There is also an R package by Victor Quesada.
I find there are two problems for Venn diagrams with more than three datasets. First, it takes long to read them and extract all information: comparing four datasets gives a diagram with 15 regions/11 overlaps, five datasets gives a diagram with 31 regions/26 overlaps! I invariably end up writing the numbers down into my own table. Secondly, the areas can’t possibly be representative for the overlap size – and this is a lost information.
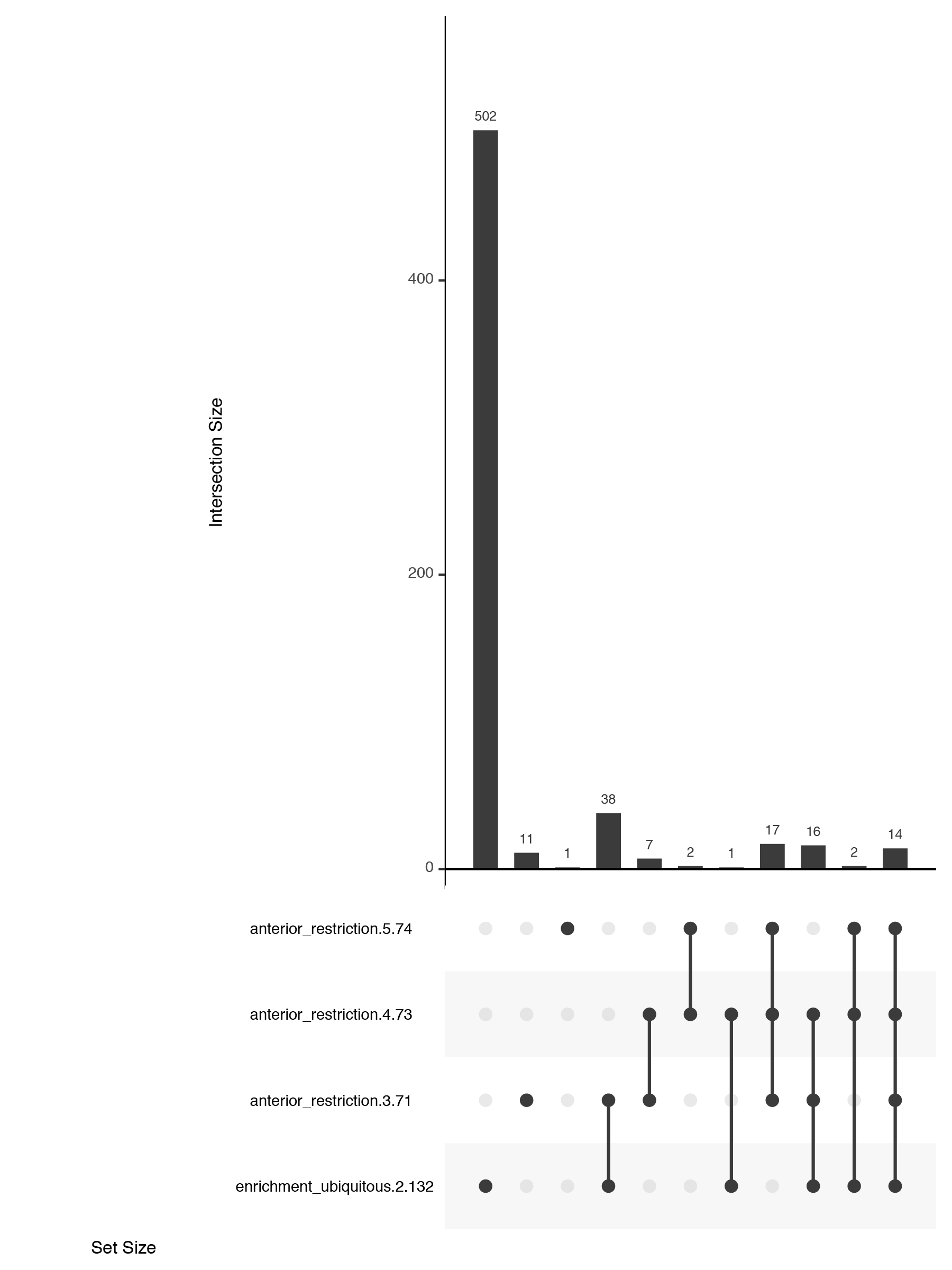
New: upset plots
An alternate solution, the upset plot, was developed by Niels Gehlenborg and Jake Conway. Presence of dataset elements in a given intersect is shown with a dot in a simple table. The size of the intersect is represented with a bar chart. Both are simple visuals that are easy to consume. Their package is available in R and simple to use.
Customising upset plots
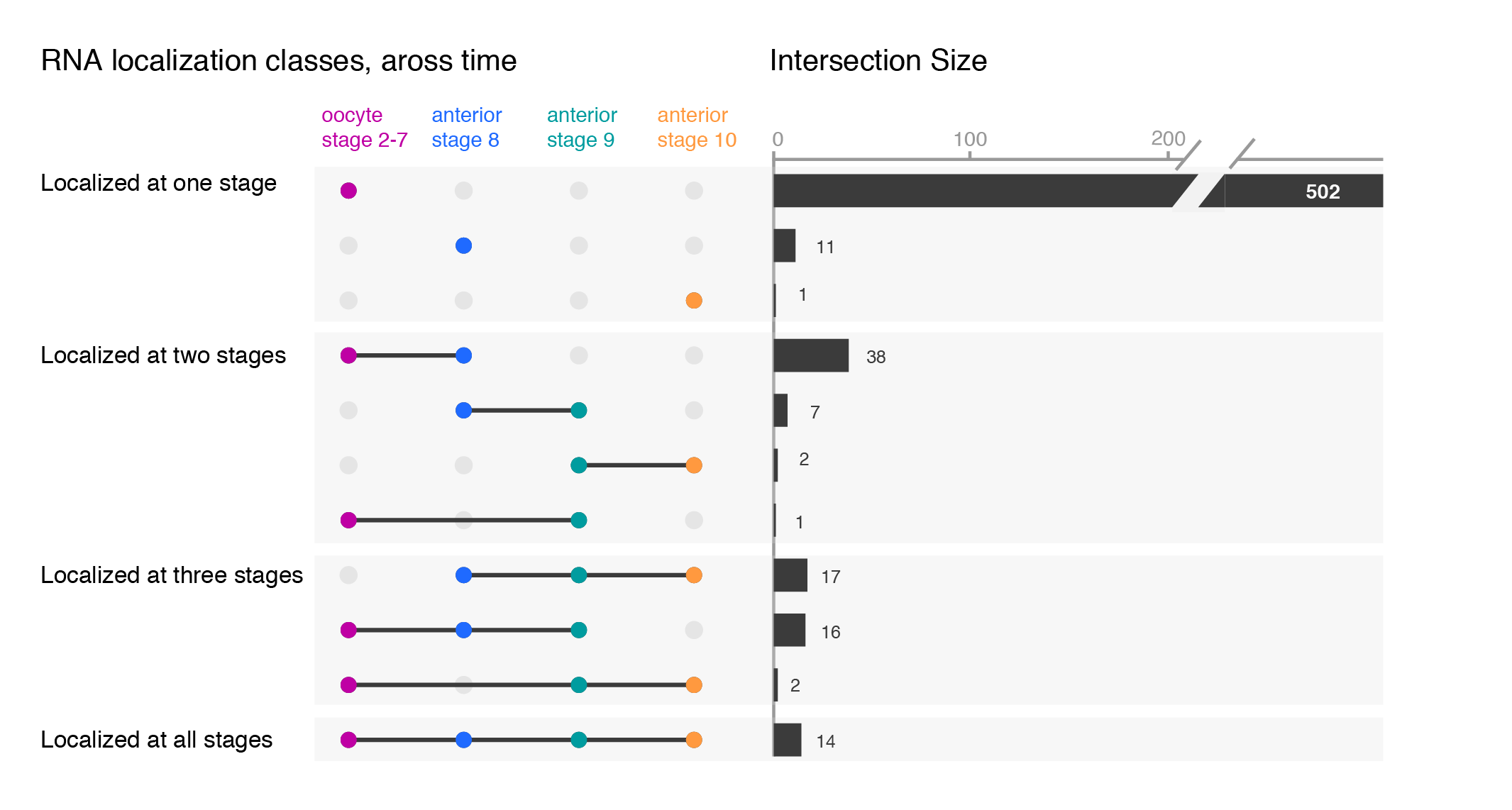
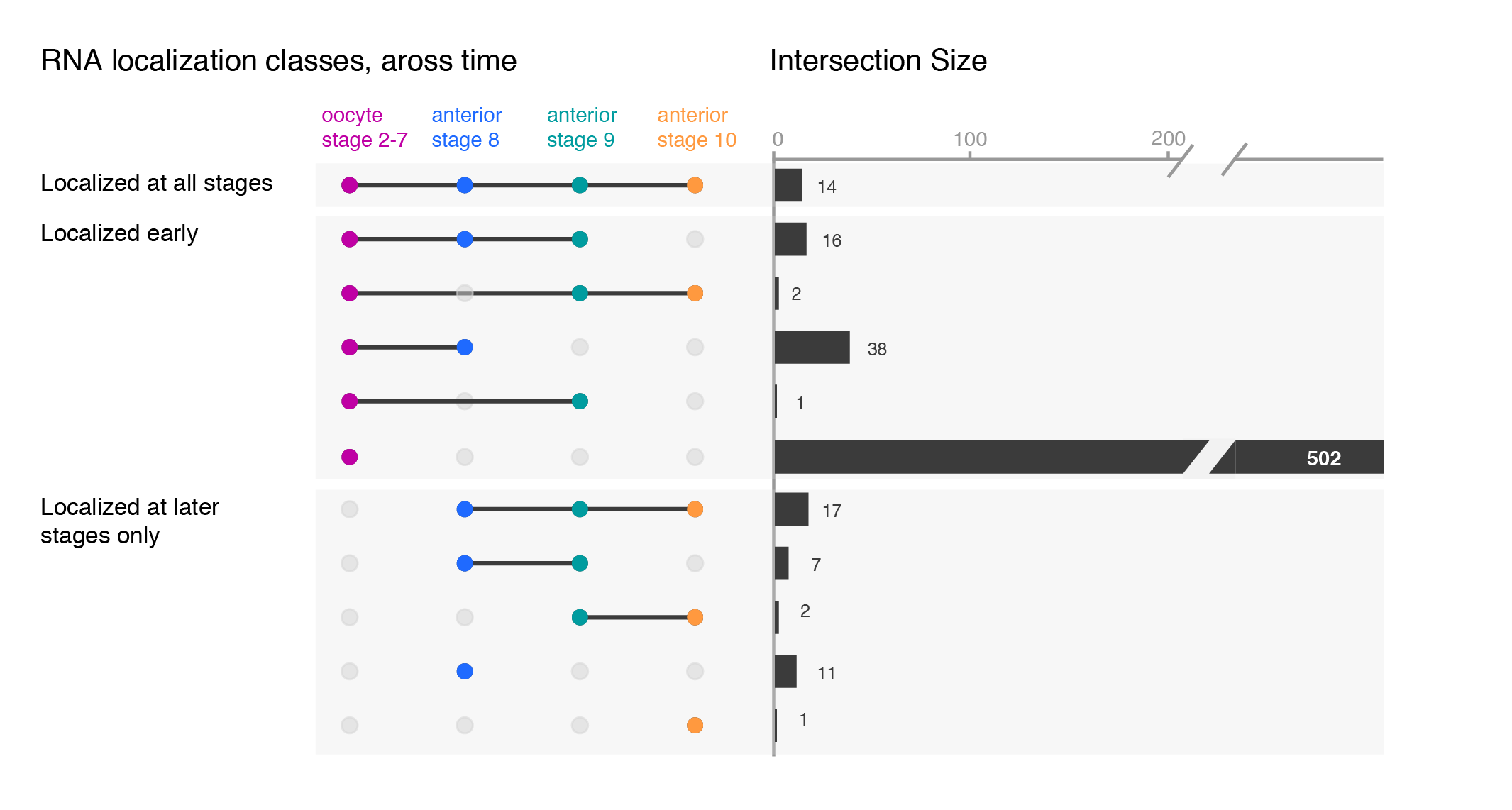
While the upset plots are simple, I think they can be improved. In upset plots the intersect is shown above the actual datasets, that serve as the legend. Basically, one is forced to read the upset from the bottom up. By flipping the plot horizontally this caveat is overcome: now the datasets are on the left, where we typically read first, and the bar is shown on the right nicely accompanying the respective set. Another improvement is to clearly label the intersects e.g. “present in one set”, “two sets” and to group them visually. Additionally, I have also color-coded the datasets to provide a quicker way of orienting the reader.
Depending on your message, you will have to find the optimal ordering strategy. I visualized the subcellular enrichments of RNAs and how they change localization during the development of the fruit fly oocyte. I would want to learn e.g. what happens to the hundreds of specific RNAs that enrich at early stages? Do they remain localized at all stages? It turns out the majority gives up their specific subcellular enrichment and instead become distributed inside the cell while other RNAs (not visualized here) take their place (more information on the biology).
Note, I did all the fine-tuning of the upset plot with illustrator but most likely it is also possible in R directly.
* Be aware that more people than you expect do not know Venn diagrams & require an introduction!
My name is André Dias and I am a PhD student in Portugal. I work in a wonderful institute called Instituto Gulbenkian de Ciência, under the supervision of Moisés Mallo. I started my PhD at the beginning of 2017 and since then it has been a fantastic journey…
In 1925, after looking to the formation of the chick embryo, Holmdahl proposed that the development of the vertebrate body axis is accomplished by two separate and distinct processes, termed primary and secondary body formation1. In our lab, considering the differently regionalized vertebrate body structures: head, neck, trunk and tail, we are expanding his vision, trying to understand the molecular mechanisms that regulate the formation of these different and somewhat independent building blocks.
At the end of gastrulation, after the anlage for the most anterior part of the head is formed, the embryo starts elongating along its main body axis, as new tissue starts to be continuously generated in the caudal part of the embryo. Thus, the remaining structures of the body arise in anterior (rostral) to posterior (caudal) direction; finishing the head, making the neck, then the trunk and finally the tail. This process of axial elongation depends on groups of cells located in the caudal part of the embryo that are known to have stem cell-like properties. The Neural-Mesodermal Progenitors (NMPs) are a very important and specific subset of these cells, which seem to have the potential to generate both neural and mesodermal tissues.
Professor Valerie Wilson and her lab members at the MRC Centre for Regenerative Medicine in Edinburgh have been studying the process of axial elongation in the mouse embryo for several years. They belong to the selected group of researchers who have pioneered the characterization of the different cell populations involved in body axis formation, as well as mapping their location within the embryo2.
During the 18th International Congress of Developmental Biology in Singapore, I met Filip Wymeersch – who was then a Post-Doc in Val’s lab. Talking to him about these special cells that make up our bodies, we considered collaborating in the future by joining the expertise of both labs. An idea was put to paper, an experiment was designed… but, as a lot of things in science, we would need funding to be able to accomplish it. So, I applied to a Travelling Fellowship from “The Company of Biologists” (COB) and surprise, surprise… I got it!!
Thanks to the COB (and to the journal Development) I had one of those life/career-changing moments with the opportunity of staying in Val Wilson’s lab in Edinburgh for three weeks. During my stay, we put our ideas into practice and conducted several experiments involving single cell analyses that will help us understand the molecular mechanisms regulating, specifically, the formation of the vertebrate head and neck. And now that I’m able to locate these special cells in the embryo, both in time and space, as well as micro-dissect them, I’ll continue this work using embryos with specific genetic backgrounds that we have available in our lab, in Portugal.
For me, this opportunity given by the COB was of great importance, since it definitely took my PhD into new, cutting-edge heights. I had the possibility to learn a variety of techniques, some of them even coming all the way from Rosa Beddington, as well as to discuss several questions regarding the formation of the vertebrate body and to nurture future collaborations. I also had the opportunity to present the results of my PhD project at the MRC Centre for Regenerative Medicine and get important feedback from their community. Finally, I still had time to taste a bit of Scotland, having the opportunity to climb Arthur’s seat and venture myself for one day into the Highlands :) .
To finish I would like to express my deep thanks to the COB for the financial support that allowed all this. Indeed, in times such as the ones we are facing now where walls are being built and people are told to look no further than themselves and forget the rest of the world, initiatives like the ones supported by The Company of Biologists are of great value: science still is universal, and we have a lot to give and to learn with each other. I also would like to thank Val for receiving me in her lab, for teaching me so many things, and for the discussions and suggestions regarding my PhD project. To Filip for the friendship, for trying to make my time in Edinburgh as good as it could have been, for showing me the techniques that they have implemented in the lab and for the scientific discussions that sometimes lasted well into the night. Also to the remaining colleagues in Val’s lab and to the MRC Centre for Regenerative Medicine community (specifically Julia and Anahi). Lastly, I would like to thank Moisés, my supervisor, for all the support throughout the entire process. Thank you!
To be continued…
References
1Holmdahl D. E., Experimentelle Untersuchungen uber die Lage der Grenze primarer und sekundarer Korperentwicklung beim Huhn, Anat. Anz., 1925. 2Wymeersch F. J. et al, Position-dependent plasticity of distinct progenitor types in the primitive streak, eLife, 2016
 (No Ratings Yet)
(No Ratings Yet)

 (1 votes)
(1 votes)




 (16 votes)
(16 votes)

{kind=link}