On 2016-5-18, the second day after my first research paper 1 was published online at my third year of PhD courses, my mentor Rongwen Xi told me to take over the “EEP project”. This project had begun long before I started my PhD courses, and until my participation, has been passed along by three researchers in turn: Na Xu, Pin Huang, and Chenhui Wang; each of them subsequently graduated and moved on with their own academic or industrial paths. Encouraged by my first successful publication, I quickly agreed to take this seemingly never-ending project. I told Dr. Xi a sentence that amused me afterwards, “I won’t give it up until you give up.” This is how this long and tough process begins, and this is also the instant that determines the end of the story.
Introduction
Even as adults, we have stem cells throughout our bodies that are responsible for maintaining many of our tissues. These adult stem cells constantly divide and produce daughter cells, which, through a process called differentiation, become multiple types of mature cells. The fate of the daughter cells can be actively specified by asymmetric cell division, in which cell fate determinants are specifically segregated into one of two stem cell daughter cells 2. Alternatively, cell fate can be specified passively; in this case, cells physically depart from the self-renewal niche environment, as with the specification of cystoblasts from Drosophila germline stem cells, and the initiation of differentiation of stem cells in the mouse small intestine upon their departure from the Paneth cell niche 3,4. Despite several implications from these “renew or differentiate” fate determination events, very little is known about the molecular mechanisms by which distinct, lineage-restricted progenitor cells are generated from a common stem cell pool.
To study this question, we investigated cell fate in a multipotent intestinal stem cell (ISC) experimental model from adult fruit flies. The default mode for cell fate is that ISCs differentiate into enterocytes (EC), which have been shown to occur from approximately 90% of ISC divisions 5. However, there is a less-well-understood mode in which ISCs differentiate into pairs of enteroendocrine cells (EEs), which occur from approximately 10% of ISC divisions.
When I started to do this project, previous studies suggested that EEs are directly differentiated from ISCs, implying that the decision of EE specification may occur at the stem cell level in ISCs 6,7, but how this occurs remains unclear. It has also been revealed that the four-gene cluster acheate-scute complex (As-c) act as EE-fate-determination factors. Furthermore, one of the As-c genes, scute (sc), is both necessary and sufficient for EE specification. Nevertheless, important questions remain about both the molecular and cellular mechanisms through which Sc functions in EE fate decision, and we do not yet know how Sc is regulated in ISCs to control EE fate.
We finally answered these questions in our recent paper, in which we reported that transient activation of Sc determines both the type and number of committed progenitor cells from Drosophila ISCs.
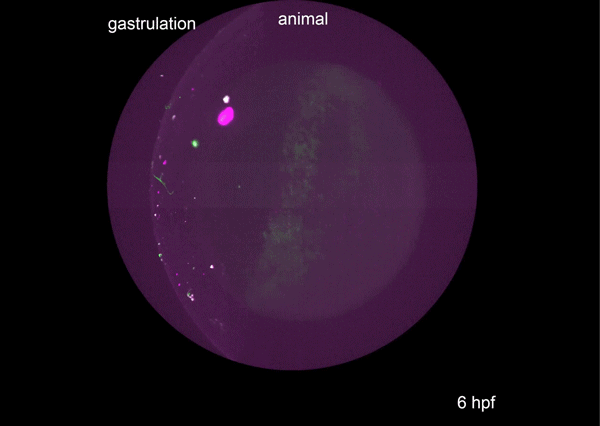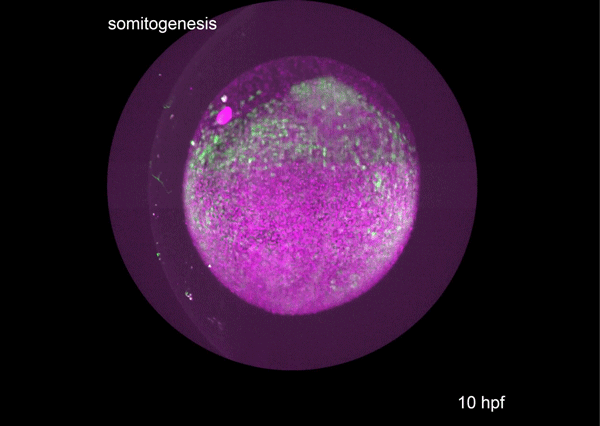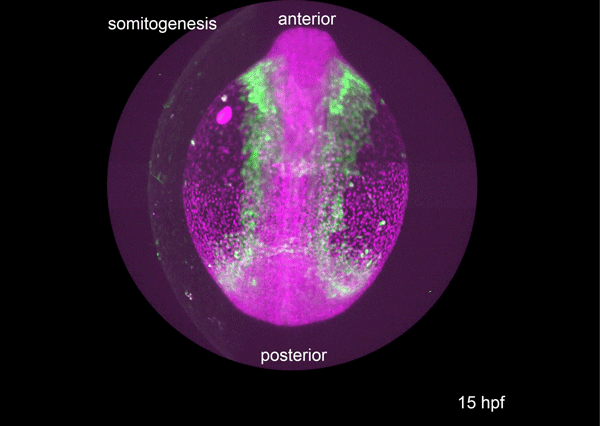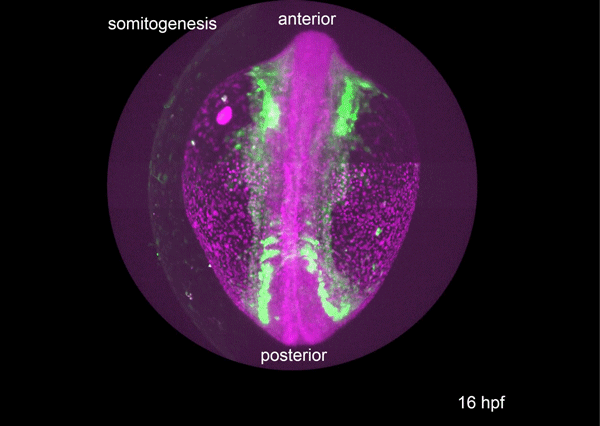
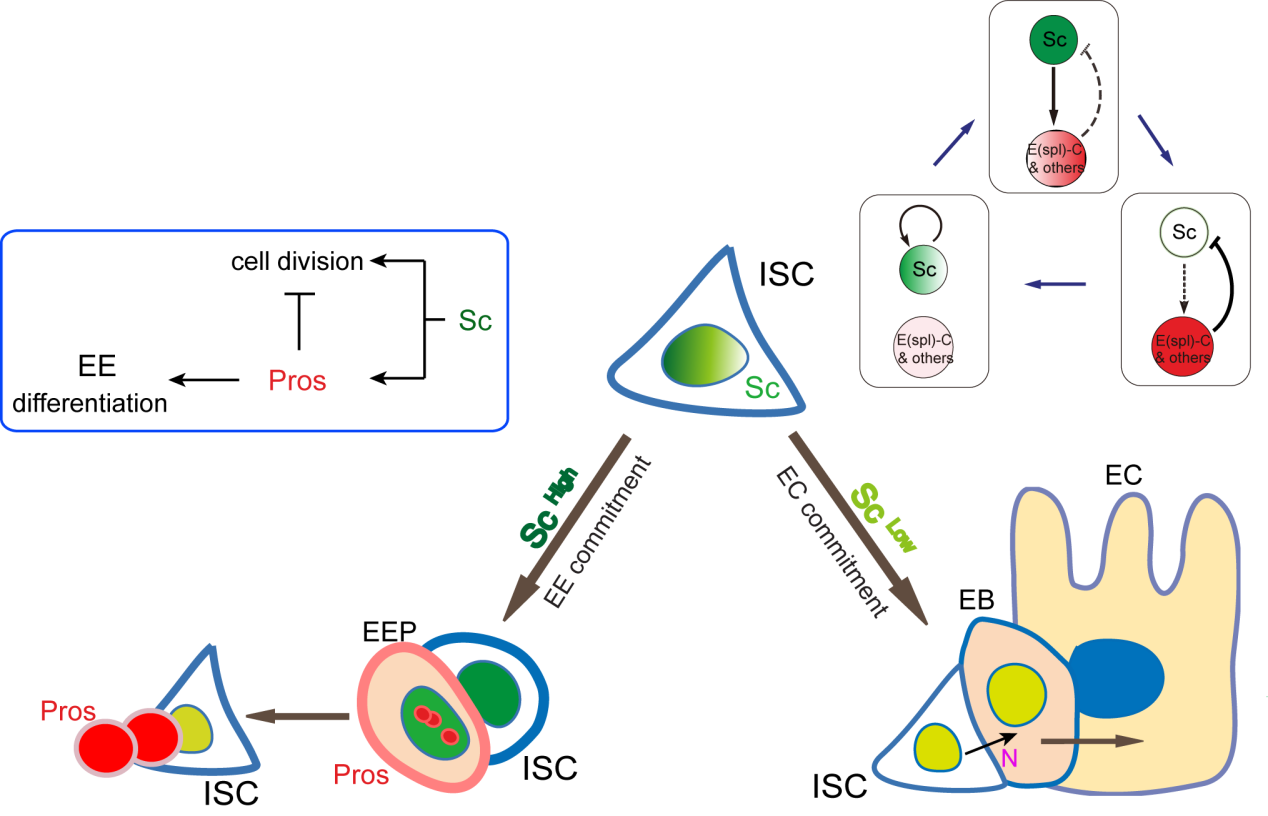
Figure 1. A graphic model to describe how ECs and EEs are respectively generated from ISCs. Notch-signaling-guided EC generation from ISCs acts as the default mode, while transient expression of Sc triggers EE generation from ISCs. Oscillatory expression of Sc in ISCs is achieved by transcriptional self-stimulation combined with a negative feedback regulation between Sc and E(spl) proteins & other Notch targets. During the generation of EEs, increased Sc expression induces asymmetric cell division that generates a new ISC and an EEP; residual Sc activity in the newly formed EEP is then able to induce one round of cell division and precisely generate a pair of EEs.
A cell fate is determined by a transiently expressed protein
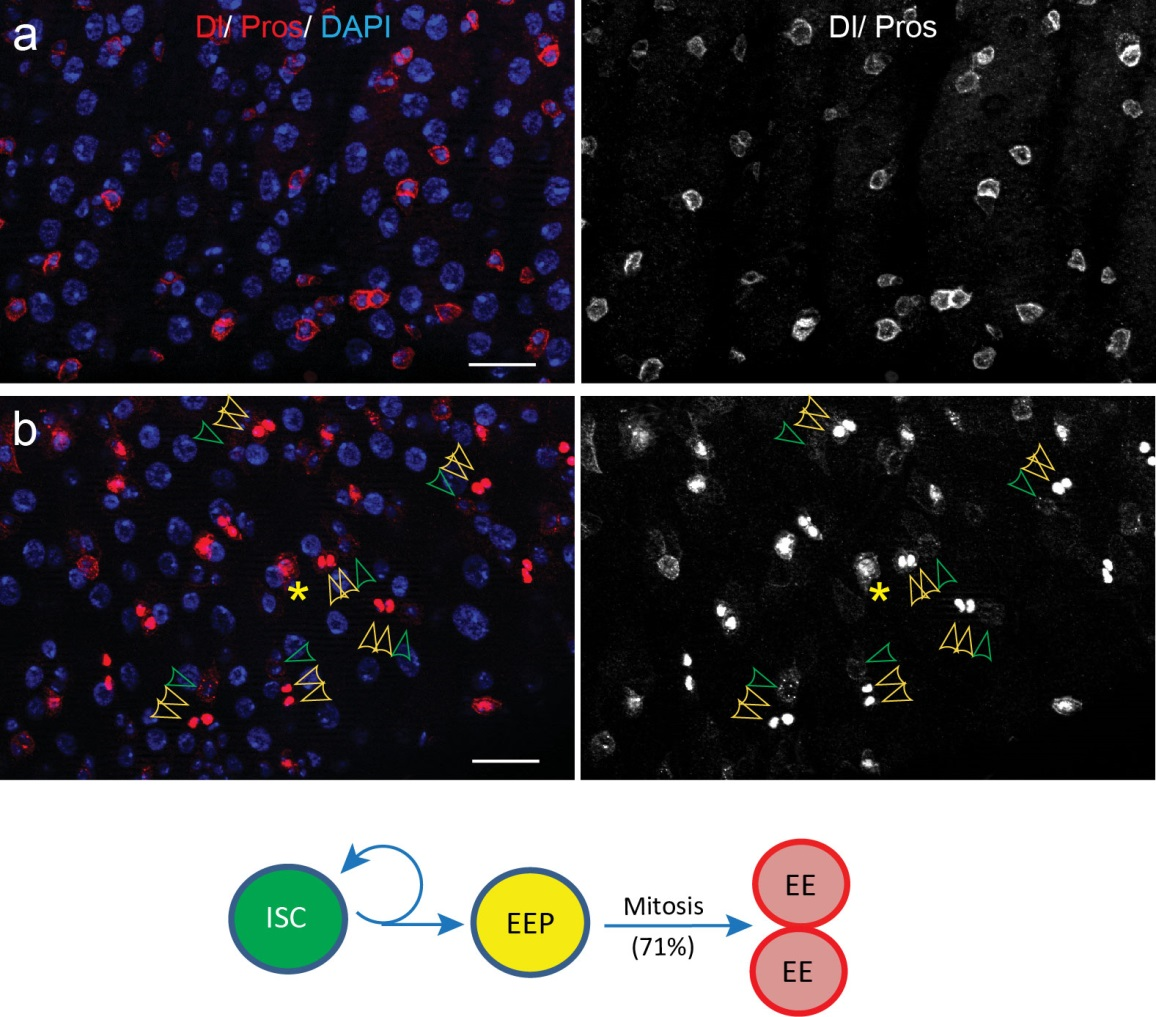
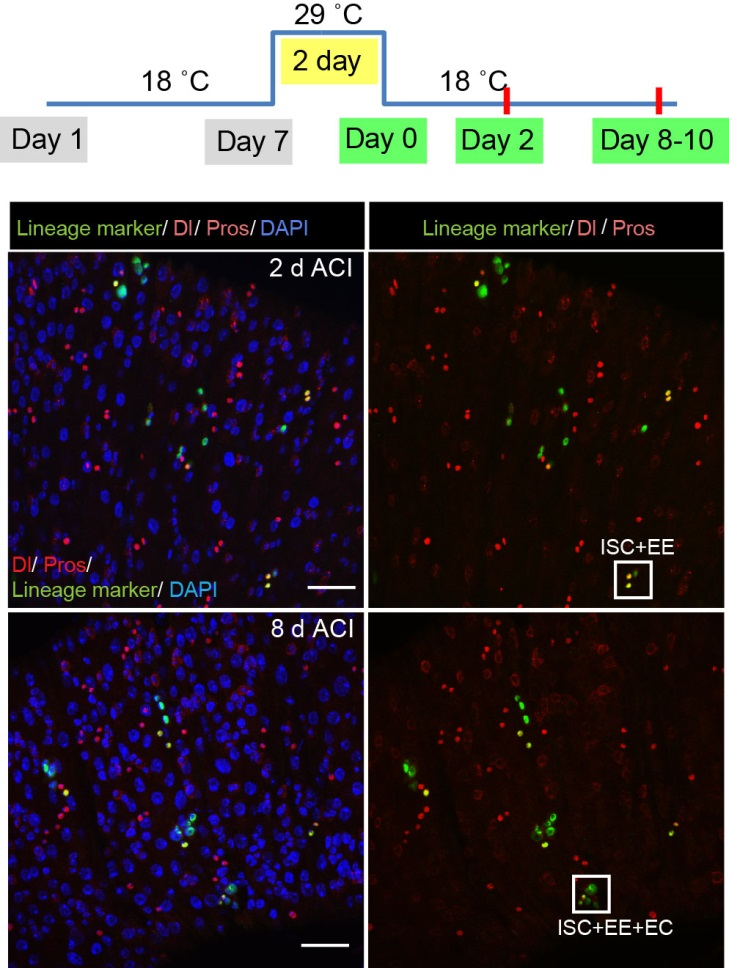
To better understand the process of EE specification in ISCs, we set up an EE regeneration assay and examined de novo EE regeneration. This assay was first beautifully set up by Na Xu and Pin Huang. Based on the finding that Sc is required for EE generation from ISCs, we temporally knocked down sc starting from the pupal stage, and this process produced flies with midguts lacking EE cells. We then used these EE-less midguts to examine the process of EE production by using temperature shift to re-introduce Sc expression in the midgut. With this assay, we discovered that (i) ISCs actually undergo an initial division to generate a new EE progenitor cell (EEP), and (ii) the EEP then undergoes one final round of cell division to produce a pair of EEs (Figure 2).
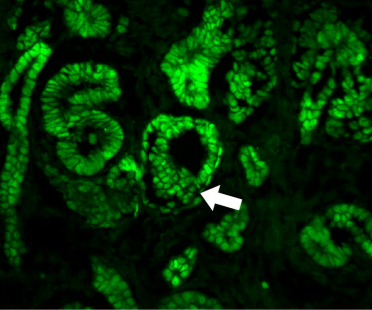
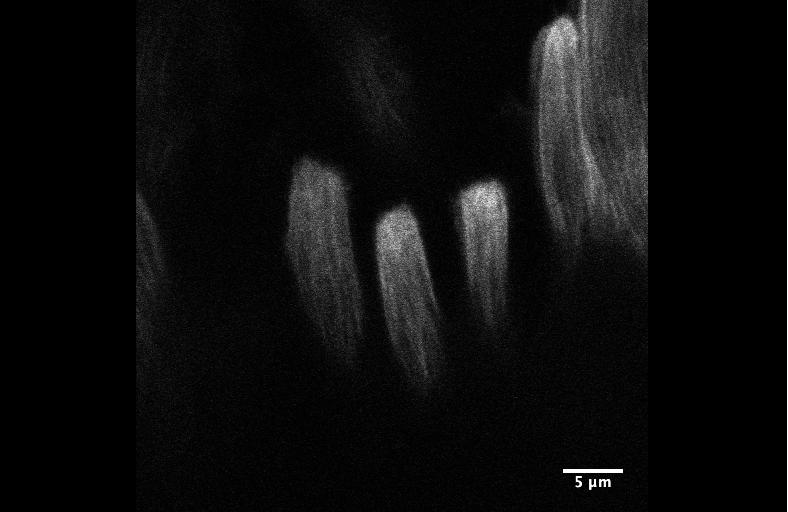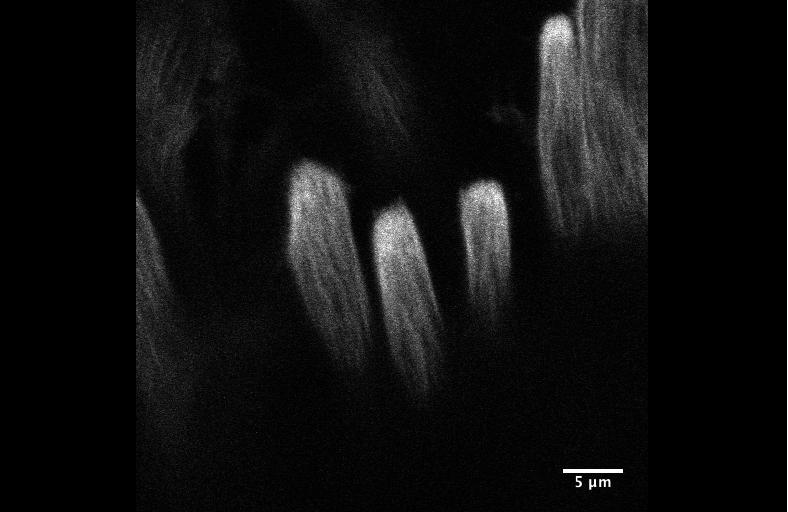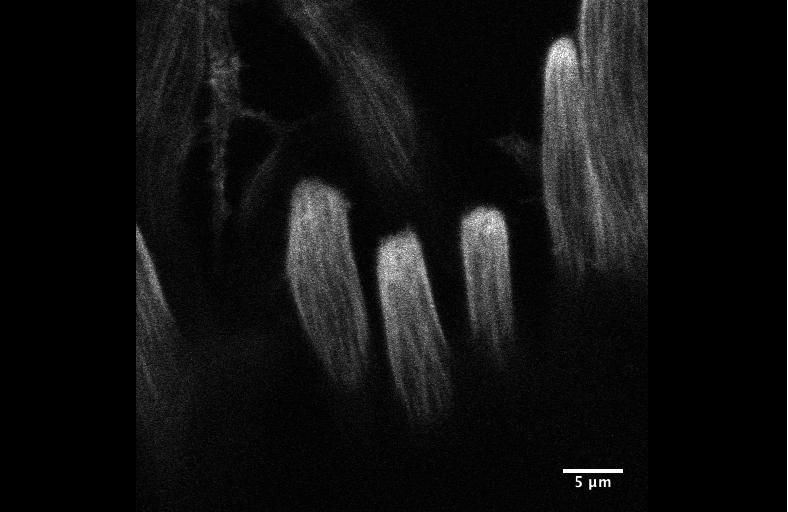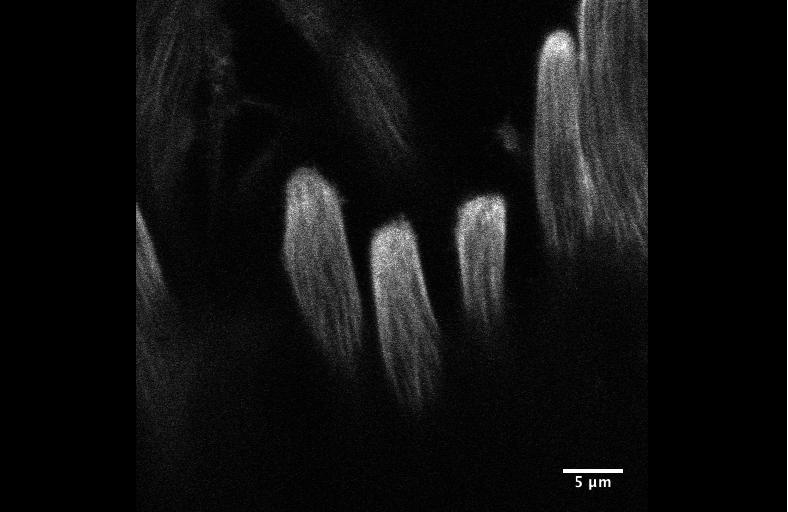
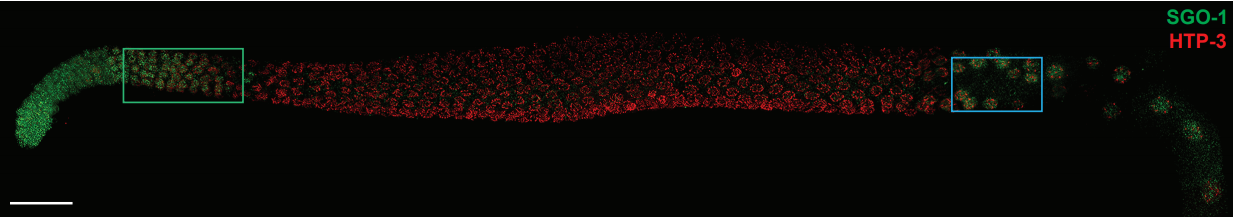
Figure 2. An EE-regeneration model reveals that ISCs self-renew during the generation of EE pairs. (a-b) Patterns of ISC (marked by anti-Dl, red on membrane) and EE cells (marked by anti-Pros, red in nuclear) during sc-RNAi mediated EE depletion (a) and the following EE regeneration (b). An ISC undergoes self-renewal before generating an EEP, and 71% of EEPs undergoes one round of mitosis to generate a pair of EEs, and the rest directly differentiate into a single EE.
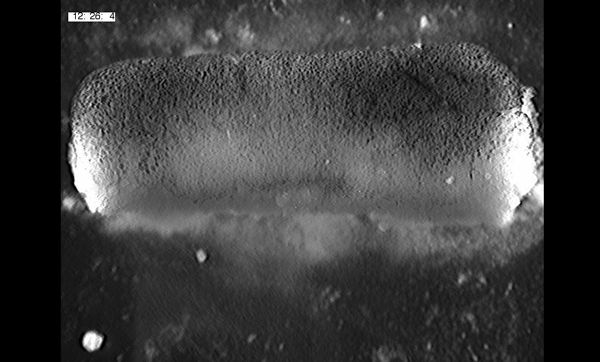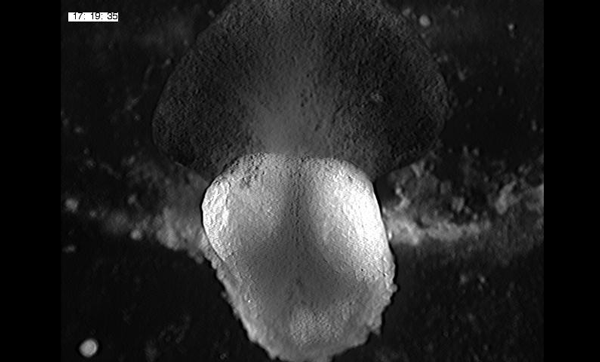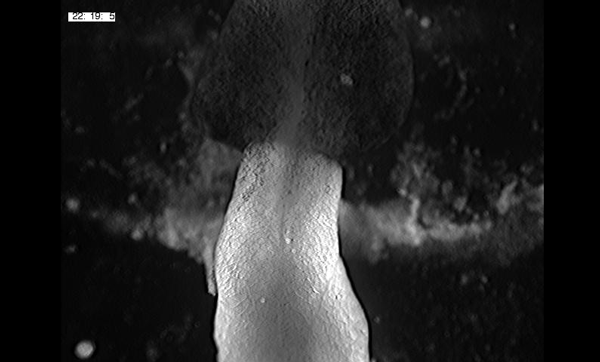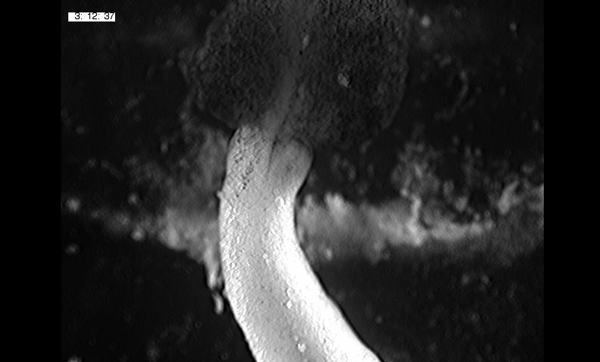
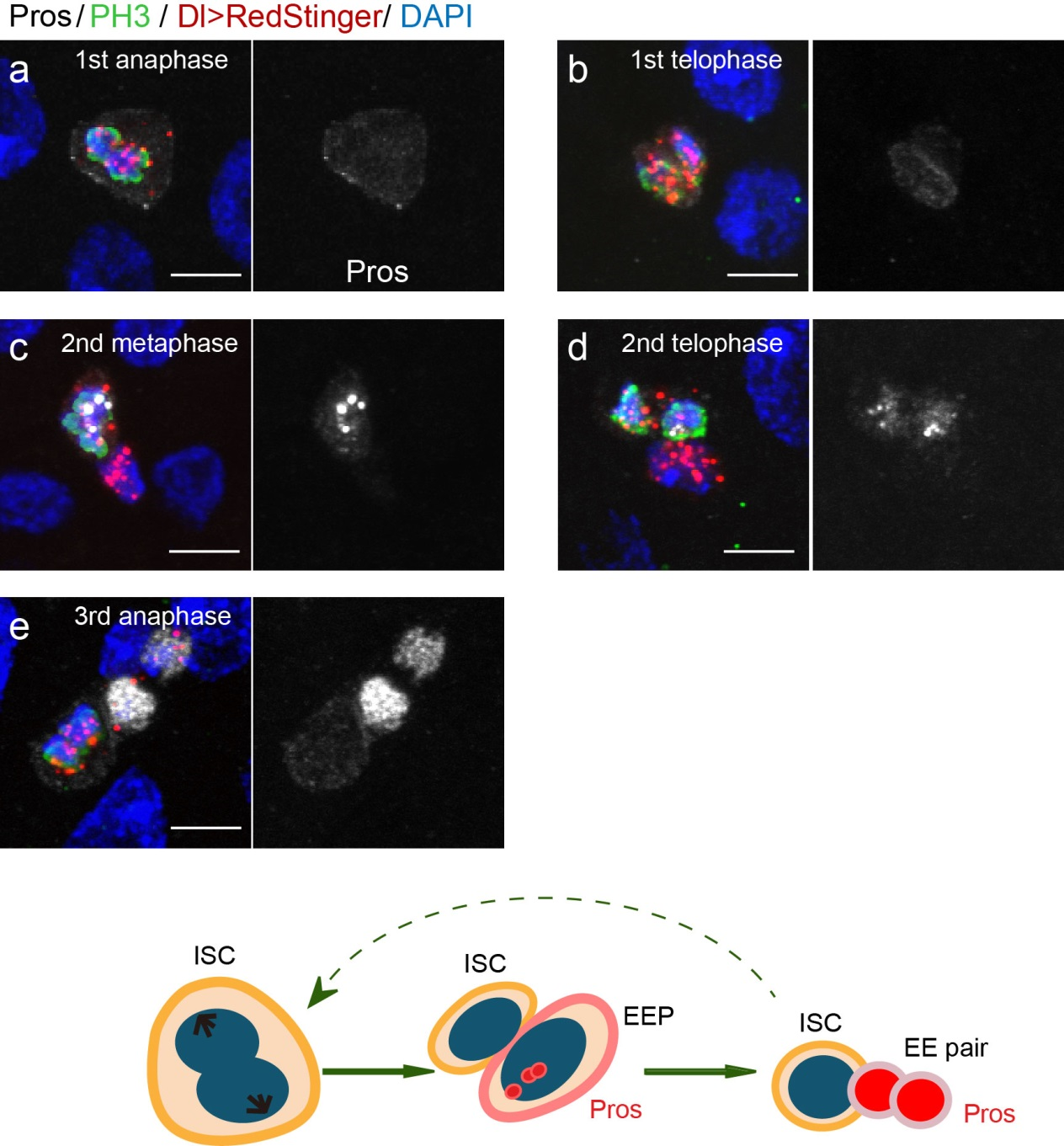
To further analyze this two-step cell division process, Chenhui Wang genetically overexpressed sc in ISCs and monitored the cellular events in a time-course experiment. Chenhui found that transient sc expression caused a rapid cell division response, and also induced expression of the EE-marker gene Pros, which is known as a potent cell-cycle inhibitor. These findings and subsequent experiments enable us to precisely define the regulatory circuitry that directs the formation of a pair of EEs from each ISC (Figure 1). Here a concern still exists that we have not given a “seeing is believing” results for cellular events of EE generation because we have not established long-term live imaging technique for fly midgut yet. To solve this problem, I expressed a UAS-RedStinger reporter in sc overexpression system. RedStinger is relatively stable and can serve as a lineage marker to trace the progeny of the originally marked ISCs. The number of cell divisions of the initially labeled ISCs could be deduced based on the mitotic marker PH3 and the number of RedStinger+ cells in a single cluster. In this experiment, I observed a tightly ordered process: The first cell division following sc overexpression occurred in ISCs (PH3+ in a one-cell clone), and at telophase of the first cell division, one of the two daughter cells began to show cytoplasmic Pros accumulation; the second cell division (PH3+ in a two-cell clone) always occurred in the Pros+ daughter cell, that is EEP; the third cell division (PH3+ in a three-cell clone) occurred again in ISCs. These observations suggest that EEs are generated from ISCs via two rounds of cell divisions: an asymmetric division of ISC to generate an EEP, and then the EEP division to produce an EE pair (Figure 3).
Figure 3. The process of sc-overexpression-induced EE generation from ISCs. (a-e) Expression of Dl>RedStinger (red), PH3 (green) and Pros (white) during sc-overexpression-induced mitosis. Sc induction in ISCs promotes asymmetric cell division that generates EEPs, which begin to show punctate nuclear Pros expression. Each EEP immediately divide once prior to terminal differentiation, yielding a pair of EEs.
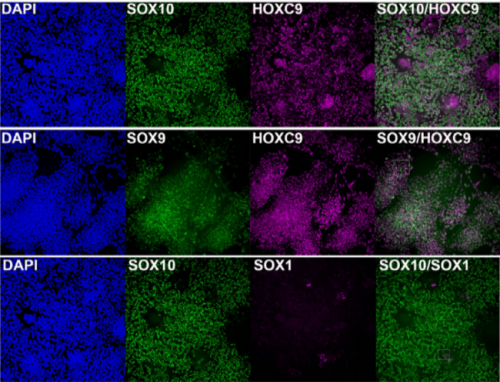
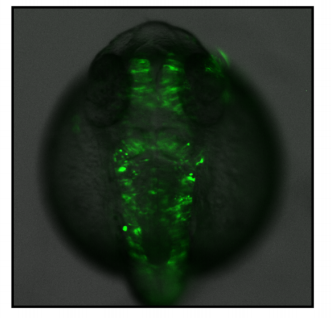
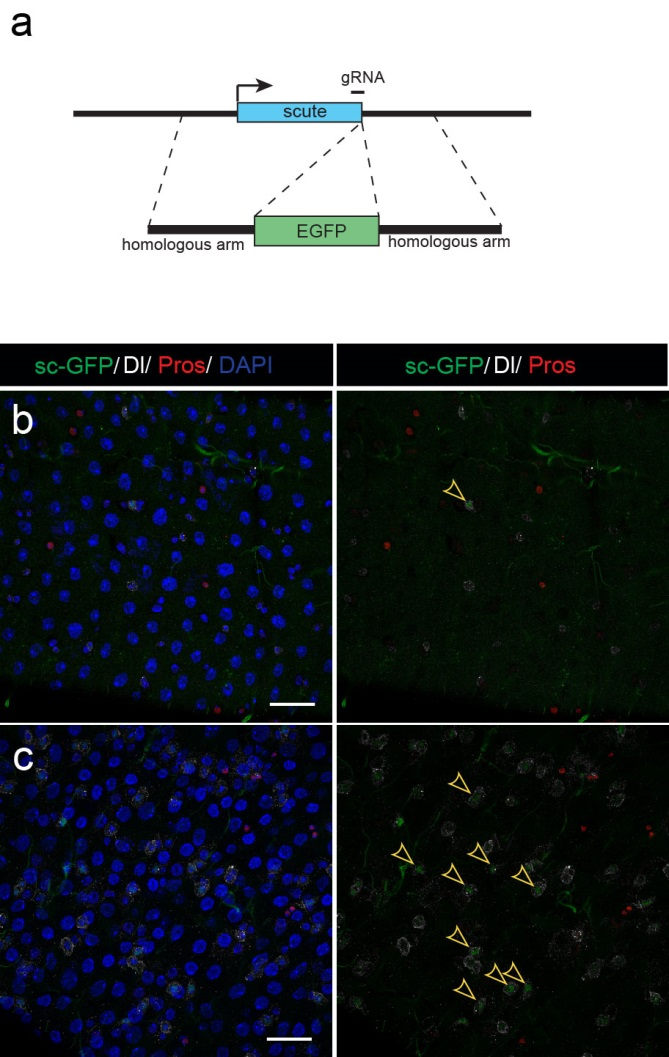
Next, to visualize the expression of Sc in midgut, Pin generated a green fluorescent protein (GFP) tagged line for Sc in collaboration with Zhongsheng Yu and Renjie Jiao from the Institute of Biophysics of the Chinese Academy of Sciences. In this line, the GFP was fused to 3’ of the Sc coding region. Initially we were a little bit disappointed as the GFP signal was too weak to visualize and all the researchers had to immunostain with anti-GFP antibody, which effectively amplified the Sc-GFP signal. Immunostaining results revealed that Sc-GFP could be observed in virtually all ISCs but the expression level is largely indistinguishable among ISCs. With improved microscopy technology, I managed to capture GFP signal in unstained samples and found that the Sc-GFP fusion protein is expressed at higher levels in ~15% of the ISCs (Figure 4). This result was exciting because it indicated that Sc may be expressed in a dynamic manner in ISCs, and in a snap shot, you may see a weak expression level in most ISCs, and increased expression levels in a small subset of ISCs.
Figure 4. Sc is expressed in a small subset of ISCs. (a) A diagram showing genomic information for C-terminal insertion of EGFP in sc gene region. (b-c) Expression of Sc-GFP (green), Dl (white) and Pros (red) in midgut of 5-7 day old flies.
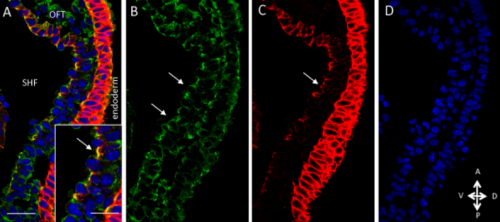
The next step was to test the cell lineage fate of these Schigh ISCs. The follow-up cell lineage tracing studies with a Sc-Gal4 line will help to do that, but there was no available Sc-Gal4 line at that time. Fortunately, from 23 (upstream and downstream) glass multiple reporter (GMR) enhancer-GAL4 lines generated for sc, I identified one GAL4 line that drove UAS-GFP expression in some diploid cells in the midgut epithelium. The density (also in ~15% ISCs), distribution, and individual variability of the GFP+ cells were largely similar to those of Sc-GFP+ cells, and about half of RFP reporter driven by this GAL4 line recapitulates Sc-GFP expression, suggesting that this GAL4 line is driven by the enhancer element for sc expression in the midgut. Cell lineage tracing studies with this GAL4 line revealed that the immediate daughter cells of Sc-GAL4+ ISCs were mainly EEs; however, these ISCs re-assume their default EC-producing fate once Sc expression is downregulated (Figures 1&5).
Figure 5. The cell lineage tracing results with the Sc-Gal4 line.
The knotty problem
With these exciting new observations, we inevitably faced a mechanistic question, “How does such transient upregulation of Sc in ISCs occur?” This question comes like a boss in video games, and has always been difficult to tackle. Studies over the decades on proneural genes have revealed that the AS-C genes in the neural cell lineages are regulated by highly-complex-cis-regulatory regions, and these regulatory regions are considered to constitute an integrating device for multiple signaling regulators and chromatin factors. Firstly came to our minds was to avoid such “net” and to set out from the reported signals that regulate EE specification in Drosophila midgut. Previous studies suggest that the Slit molecules secreted from EEs activate the Robo2 receptors of ISCs to prevent EE generation, thereby establishing a negative feedback to coordinate EE production with tissue demand. However, Sc expression pattern was unaltered in Robo2 mutants, in which the excessive EE phenotype was prominent. Considering Robo2 activation in ISCs is not sufficient to prevent EE production from ISCs, this mechanism appears to be a modulator rather than a key component in the EE fate decision process. Thus, I had to go back to hit the core of the question, the transcriptional control of As-c genes.
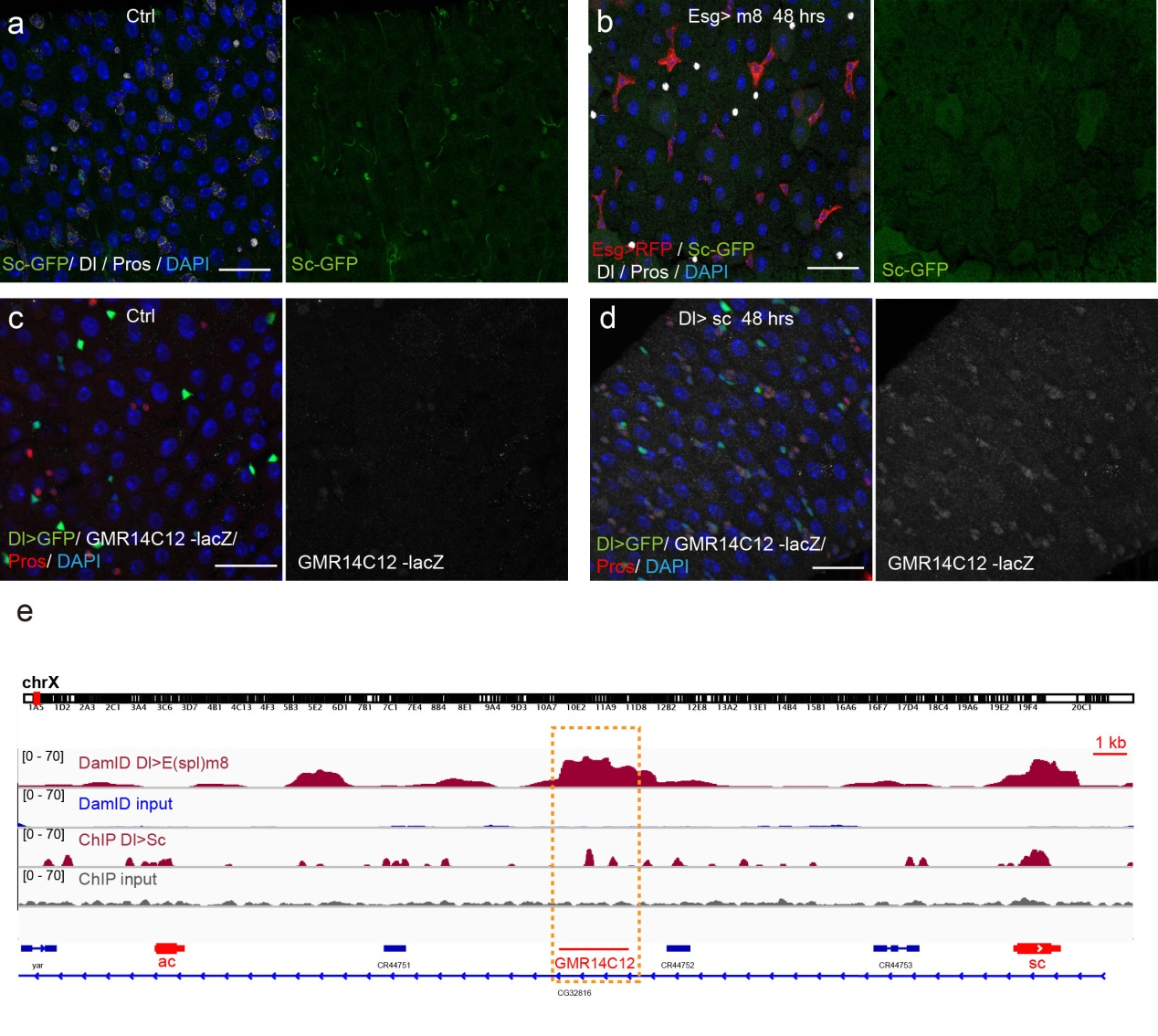
Previous studies on early Drosophila development have suggested reciprocal regulatory relationships between AS-C genes and the enhancer of split complex (E(spl)) genes, which are known as the Notch target genes. Inspired by these reports, I screened a number of candidate reporters for individual E(spl) genes, and identified a single reporter, m8-lacZ, which showed a weak, but similar expression pattern to Sc in wild type guts. To characterize the regulatory relationship between Sc and E(spl)m8, I transiently overexpressed sc in ISCs, and surprisingly saw robust upregulation of m8-lacZ expression in all ISCs. Notably, co-expressing Notch-RNAi did not prevent the upregulation of m8-lacZ expression caused by sc overexpression, suggesting that E(spl)m8 expression is independent of Notch activity in ISCs. To test whether such regulatory relationship similarly applies to other E(spl) genes, I sorted out sc-overexpressed ISCs for mRNA profiling by RNA-seq analysis. Strikingly, in addition to m8, many other E(spl) genes, including m4, m6, m7, mγ, and mδ were strongly upregulated upon sc overexpression. By combining genetic assays and ChIP-seq analysis, we showed Sc could bind to the enhancer regions of many E(spl) genes, and directly upregulate these E(spl) genes in ISCs (Figure 6).
It’s then instinctive to consider whether these E(spl) genes, also known as neural fate repressors, would in turn negatively regulate Sc expression. By combining genetic assays and targeted DamID analysis using a E(spl)m8-Dam fusion line, we showed that E(spl)m8 suppresses sc expression by directly binding to the enhancer region of sc. The direct two-way regulation between Sc and E(spl)m8 form a typical negative feedback regulatory loop, which may explain the transient activation pattern of Sc in ISCs (Figure 6).
The question still has half part unanswered, “how does sc initially build up?” Searching for other transcriptional activators, like other bHLH activators as reported, would make this question a “chick and egg” issue. Interestingly, Sc has been reported to transcriptionally self-stimulate itself, which acts as an essential mechanism for proneural protein accumulation during sensory organ development. To test whether self-stimulation of Sc also occurs in ISCs, we constructed LacZ transcriptional reporter for sc using the Sc-Gal4 enhancer fragment that we had identified. This lacZ reporter was barely detectable in WT midgut epithelium, but effectively induced in ISCs when sc was transiently induced. ChIP-seq data analysis also revealed two Sc binding peaks within this Sc-Gal4 enhancer region (Figure 6). Thus, Sc is able to stimulate its own transcription directly by binding to sc enhancer. Together, our results suggest that two feedback regulatory loops control the transient upregulation of Sc in ISCs prior to EE fate commitment. There is a transcriptional self-stimulation loop that allows Sc to gradually build up and eventually reach a high level to induce EEP specification, and there is a negative feedback regulation loop between Sc and E(spl) genes that returns sc expression back to the baseline level (Figure 1).
Figure 6. Regulatory feedback loops control Sc expression in ISCs. (a-b) Overexpression of m8 rapidly reduced sc-GFP and Dl expression in all ISCs. (c-d) GMR14C12-lacZ (LacZ reporter for Sc-Gal4 line) was nearly undectable in normal midgut epithelium. Overexpression of sc in ISCs led to GMR14C12-lacZ expression in progenitor cells and newly formed EEs. (e) DamID analysis for E(spl)m8 and ChIPseq analysis for Sc in ISCs revealed binding activities for both E(spl)m8 and Sc at the GMR14C12 region.
The beginning of the end
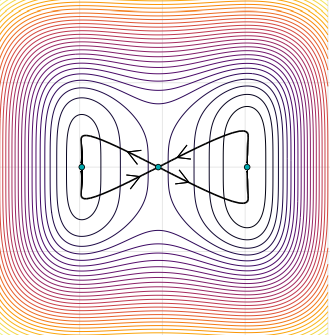
Given that negative feedback is a common mechanism underlying biochemical oscillations in virtually all organisms, the feedback loops between Sc and E(spl) genes could plausibly be the driver of an oscillatory expression pattern for Sc in ISCs; in theory such oscillatory expression could potentially serve as an internal timer for periodic production of EEs from ISCs. This clock mechanism would be similar to what is known about the circadian clocks, a biological research field that was recently honored with the 2017 Nobel Prize for Physiology or Medicine. We are obviously very excited about the findings and potential implications. However, this is just a tip of iceberg, future cellular and molecular analysis, likely in combination with in vivo live imaging work will allow further testing and refining of the oscillation model proposed in our study, and such experiments will determined whether and how any internal timer is regulated by certain endogenous and/or environmental cues, and whether the oscillation model is generally applicable in other tissue stem cells, including that in humans.
Finally, I want to say that I am very fortunate and grateful to be a part of such a wonderful research team and work on such an exciting project. This work would not be possible without the contribution and help from our past and current lab members, especially Na Xu, Chenhui Wang, and Pin Huang, as well as informaticians Huanwei Huang and Tao Cai at NIBS. I especially want to thank my mentor Dr. Xi for his great guidance and trust, as well as his helpful advice on the writing of this article. As you can imagine, in addition to the “high” moments when the exciting results were first observed, I also had many upset and head-scratching moments during the course of this study. These experiences have endowed me a lot on how to explore, to observe, to cooperate, to write, and to persevere. I believe that no matter how hard it seems like, if you continue to stay focused and think hard, great things may eventually happen, in an instant.
A few days back over dinner at a CNV gathering, Theresia Gutmann from the Coskun lab casually told me about her PhD work. In collaboration with the Rockefeller University NYC, Theresia had visualized the changing conformation of the human insulin receptor upon insulin binding (paper). I made a sketchnote summarizing their discovery of a conformational switch that could explain how the insulin receptor transforms information about extracellular ligand binding into an intracellular activity to react by taking up glucose!
A postdoc position is available in the Lehoczky Lab (Brigham and Women’s Hospital/Harvard Medical School). The lab is focused on understanding the molecular basis of mouse digit tip regeneration, with the ultimate goal of teasing apart the genetic pathways necessary for this process. For more information about the lab see LehoczkyLab.org
Applicants with a strong background in regenerative biology, genetics, developmental biology, and/or molecular biology are encouraged to apply. Prior experience with mouse genetics is preferred. Experience with RNAseq analysis is a plus.
Interested candidates should provide: 1) cv, 2) a brief letter detailing your interest in the lab and relevant past research experience, and 3) contact information for three references who can comment on your research
Application materials and any questions regarding the position should be sent to Jessica: jlehoczky@bwh.harvard.edu
Our latest monthly trawl for developmental biology (and other cool) preprints. Let us know if we missed anything.
On February 20th, The Company of Biologists launched preLights, a community-led preprint highlighting service. A panel of early career researchers (the ‘preLighters’) select and comment on recent preprints that caught their eye, and encourage preprint authors to answer any questions about the work that they had. So far it looks great, and the developmental biology content has been especially good (see the dedicated subjectcategory). We’d love to know what you think: you can contact the team via the site or the Twitter feed.
The idea behind the site was influenced in part by this list – as it got longer and longer (reflecting increased preprint usage), we were wondering how else we could encourage and promote the discussion of preprints, and the preLights idea took form. Rest assured that this list will live on, at least until the point at which it gets impossibly long!
And here’s the list – all the developmental biology I could find, plus relevant and cool other preprints thrown in for good measure.
The preprints were hosted on bioRxiv, PeerJ, andarXiv. Use these links to get to the section you want:
Axial progenitors after 8 days of differentiation, from Frith, et al.’s preprint
Human axial progenitors generate trunk neural crest cells. Thomas J.R. Frith, Ilaria Granata, Erin Stout, Matthew Wind, Oliver Thompson, Katrin Neumann, Dylan Stavish, Paul R Heath, James O.S. Hackland, Konstantinos Anastassiadis, Mina Gouti, James Briscoe, Valerie Wilson, Mario R Guarracino, Peter W Andrews, Anestis Tsakiridis
Zebrafish embryogenesis from Hess, et al.’s preprint
A conserved regulatory program drives emergence of the lateral plate mesoderm. Christopher Hess, Karin Dorien Prummel, Susan Nieuwenhuize, Hugo Parker, Katherine W. Rogers, Iryna Kozmikova, Claudia Racioppi, Sibylle Burger, Eline C. Brombacher, Alexa Burger, Anastasia Felker, Elena Chiavacci, Gopi Shah, Jan Huisken, Zbynek Kozmik, Lionel Christiaen, Patrick Mueller, Marianne Bronner, Robb Krumlauf, Christian Mosimann
Hedgehog signaling controls progenitor differentiation timing during heart development. Megan Rowton, Andrew D. Hoffmann, Jeffrey D. Steimle, Xinan Holly Yang, Alexander Guzzetta, Sonja Lazarevic, Chul Kim, Nikita Deng, Emery Lu, Jessica Jacobs-Li, Shuhan Yu, Erika Hanson, Carlos Perez-Cervantes, Sunny Sun-Kin Chan, Kohta Ikegami, Daniel J. Garry, Michael Kyba, Ivan P. Moskowitz
Neutralizing Gatad2a-Chd4-Mbd3 Axis within the NuRD Complex Facilitates Deterministic Induction of Naive Pluripotency. Nofar Mor, Yoach Rais, Shani Peles, Daoud Sheban, Alejandro Aguilera-Castrejon, Asaf Zviran, Dalia Elinger, Sergey Viukov, Shay Geula, Vladislav Krupalnik, Mirie Zerbib, Elad Chomsky, Lior Lasman, Tom Shani, Jonathan Bayerl, Ohad Gafni, Suhair Hanna, Jason Buenrostro, Tzachi Hagai, Hagit Masika, Yehudit Bergman, William J. Greenleaf, Miguel A. Esteban, Yishai Levin, Rada Massarwa, Yifat Merbl, Noa Novershtern, Jacob H. Hanna
Adaptive Reduction of Male Gamete Number in a Selfing Species. Takashi Tsuchimatsu, Hiroyuki Kakui, Misako Yamazaki, Cindy Marona, Hiroki Tsutsui, Afif Hedhly, Dazhe Meng, Yutaka Sato, Thomas Stadler, Ueli Grossniklaus, Masahiro M. Kanaoka, Michael Lenhard, Magnus Nordborg, Kentaro K. Shimizu
Firefly genomes illuminate parallel origins of bioluminescence in beetles. Timothy R Fallon, Sarah E Lower, Ching-Ho Chang, Manabu Bessho-Uehara, Gavin J Martin, Adam J Bewick, Megan Behringer, Humberto J Debat, Isaac Wong, John C Day, Anton Suvorov, Christian J Silva, Kathrin F Stanger-Hall, David W Hall, Robert J. Schmitz, David R Nelson, Sara Lewis, Shuji Shigenobu, Seth M Bybee, Amanda M Larracuente, Yuichi Oba, Jing-Ke Weng
10 Aquilegia species species from Filiaut, et al.’s preprint
The genome of the water strider Gerris buenoi reveals expansions of gene repertoires associated with adaptations to life on the water. David Armisen, Rajendhran Rajakumar, Markus Friedrich, Joshua B Benoit, Hugh M Robertson, Kristen A Panfilio, Seung-Joon Ahn, Monica F Poelchau, Hsu Chao, Huyen Dinh, HarshaVardhan Doddapaneni, Shannon Dugan-Perez, Richard A Gibbs, Daniel ST Hughes, Yi Han, Sandra L Lee, Shwetha C Murali, Donna M Muzny, Jiaxin Qu, Kim C Worley, Monica Munoz-Torres, Ehab Abouheif, Francois Bonneton, Travis Chen, Li-Mei Chiang, Christopher P. Childers, Andrew G Cridge, Antonin JJ Crumiere, Amelie Decaras, Elise M Didion, Elizabeth Duncan, Elena N Elpidina, Marie-Julie Fave, Cedric Finet, Chris GC Jacobs, Alys Jarvela, Emily J Jennings, Jeffery W Jones, Maryna P Lesoway, Mackenzie Lovegrove, Alexander Martynov, Brenda Oppert, Angelica Lilico-Ouachour, Arjuna Rajakumar, Peter N Refki, Andrew J Rosendale, Maria Emilia Santos, William Toubiana, Maurijn van der Zee, Iris M Vargas Jentzsch, Aidamalia Vargas Lowman, Severine Viala, Stephen Richards, Abderrahman Khila
In vivo CRISPR-Cas gene editing with no detectable genome-wide off-target mutations. Pinar Akcakaya, Maggie L. Bobbin, Jimmy A. Guo, Jose Malagon Lopez, M. Kendell Clement, Sara P. Garcia, Mick D. Fellows, Michelle J. Porritt, Mike A. Firth, Alba Carreras, Tania Baccega, Frank Seeliger, Mikael Bjursell, Shengdar Q. Tsai, Nhu T. Nguyen, Roberto Nitsch, Lorenz Mayr, Luca Pinello, Mohammad Bohlooly-Y, Martin J. Aryee, Marcello Maresca, J. Keith Joung
Multiple laboratory mouse reference genomes define strain specific haplotypes and novel functional loci. Jingtao Lilue, Anthony G Doran, Ian T Fiddes, Monica Abrudan, Joel Armstrong, Ruth Bennett, William Chow, Joanna Collins, Anne Czechanski, Petr Danecek, Mark Diekhans, Dirk-Dominic Dolle, Matt Dunn, Richard Durbin, Dent Earl, Anne Ferguson-Smith, Paul Flicek, Jonathan Flint, Adam Frankish, Beiyuan Fu, Mark Gerstein, James Gilbert, Leo Goodstadt, Jennifer Harrow, Kerstin Howe, Mikhail Kolmogorov, Stefanie Koenig, Chris Lelliott, Jane Loveland, Richard Mott, Paul Muir, Fabio Navarro, Duncan Odom, Naomi Park, Sarah Pelan, Son K Phan, Michael Quail, Laura Reinholdt, Lars Romoth, Lesley Shirley, Cristina Sisu, Marcela Sjoberg-Herrera, Mario Stanke, Charles Steward, Mark Thomas, Glen Threadgold, David Thybert, James Torrance, Kim Wong, Jonathan Wood, Fengtang Yang, David J Adams, Benedict Paten, Thomas M Keane
Reproducible big data science: A case study in continuous FAIRness. Ravi K Madduri, Kyle Chard, Mike D’Arcy, Segun C Jung, Alexis Rodriguez, Dinanath Sulakhe, Eric W Deutsch, Cory Funk, Ben Heavner, Matthew Richards, Paul Shannon, Ivo Dinov, Gustavo Glusman, Nathan Price, John D Van Horn, Carl Kesselman, Arthur W Toga, Ian Foster
A NIH-funded postdoctoral position is available as early as April 1st in the laboratory of Katherine Fantauzzo in the Department of Craniofacial Biology at the University of Colorado Anschutz Medical Campus to study the in vivo dynamics of PDGFR dimer-specific formation and the resulting effects on gene expression and cell activity during mouse craniofacial development. This project will utilize an array of complementary approaches such as bimolecular fluorescence complementation, mass spectrometry and conditional mutagenesis in the mouse embryo, with the ultimate goal of providing therapeutic directions aimed at the treatment of human birth defects such as cleft lip and palate. We are seeking highly motivated, creative and interactive applicants with the ability to work independently. Preference will be given to applicants with a strong background in biochemistry and/or mouse developmental biology who published a first-author paper as a result of their Ph.D. work. More information about our group and research interests can be found on our laboratory website (http://www.fantauzzolab.org). Interested candidates should apply through the CU careers website (https://cu.taleo.net/careersection/2/jobdetail.ftl?job=12799&lang=en) with a letter of interest, a curriculum vitae and contact information for three professional references.
How do cells give rise to the functional architecture of the brain? This is no longer a neuroscience-only question. Indeed, it is a cellular, genetic, developmental, mechanical, and material problem that requires experts from all of these disciplines working together to understand how the brain works! Yet, from this architectural design perspective, it is very hard to unite the leaders in these distinct fields of research to find an answer to this complex problem. Excitingly, this was exactly what happened at the Company of Biologists workshop entitled “Thinking beyond the dish: taking in vitro neural differentiation to the next level” organized by Madeline Lancaster and Denis Jabaudon. As some of the early-career researchers invited to participate, we each provide our perspectives on this amazing workshop and we are extremely grateful to the organizers and the Company of Biologists for putting this together.
GUILLERMO GOMEZ Group Leader, Centre for Cancer Biology, SA Pathology and the University of South Australia
Being myself a mechano-biologist, I was very excited about the brain organoids developed by Madeline Lancaster and Jürgen Knoblich and I became more curious on what would be next, what is the future of this technology, something that seems is becoming closer to science fiction. This was the key layout for this meeting: Thinking “beyond” the dish…., which by far exceeded my expectations.
The meeting was small but excellent. We had great talks on how we can create different types of materials to manipulate almost all class of its properties, and now, more excitingly, doing it precisely in space and time, to control cell behavior. We had also geneticists who show how single cell transcriptomics allows the creation of “expression trees” that link all the different cells that form these minibrains and described the genetic network architectures that contribute to the robustness of brain development in the early mammalian embryo (so everyone looks similar during gestation) but which then diversifies when we become more mature (so everyone looks different later). It was also really exciting how using this technology now it is possible to establish neural circuits based on organoids and also how these could contribute, for example, to the restoration of brain tissue to improve recovery in brain cancer patients after the resection of the tumor.
But where do we go now? We discussed it a lot through this meeting and my feeling is that we are still far from being able to integrate all these aspects because of its complexity and some limitations of the approach, to be able to make entire brains in the dish. But we are seeing the light on this technology to understand in a more physiologically relevant setting the fundamentals of the brain architecture and how is it affected in different type of diseases, for which strong interdisciplinary interactions are crucial. This meeting has seeded the grounds to be able to do it and gave me the chance to meet the leaders in these interdisciplinary areas, which has really fueled me with ideas and new perspectives about this problem. This is exactly what I needed at this very early stage of my independent career.
CRISTIANA CRUCEANU Postdoctoral Fellow, Max Planck Institute of Psychiatry, Munich, Germany.
I first became aware of the Company for Biologists workshops at the recommendation of one of my mentors, who suggested I would really enjoy the topic – and the format. She was right. If selected, I could have one of 10 coveted early-career scientist spots, and join 20 senior thought leaders in my field to learn about the latest research, discuss future directions, and potentially build collaborations. So I put in an application for the workshop entitled “Thinking beyond the dish: taking in vitro neural differentiation to the next level”, and got to attend from 4 – 7 February 2018. For a psychiatric geneticist looking to update our available in vitro model systems for the human brain using organoids, this seemed just perfect.
I was quite thrilled to test my research and ideas with a topically diverse audience, yet intimately focused on one important topic. How can cerebral organoids, one of the most promising developments of recent years, achieve the status of ‘workhorse of neuroscience’? I credit the organizers, Denis Jabaudon and Madeline Lancaster, for bringing together an eclectic group of scientists and engineers who covered the spectrum from neurodevelopment to bioengineering to psychiatry. The talks were wonderful for laying out the complex problem ahead, and the strides currently being made toward addressing it. The venue – a historical stately manor named Wiston house cradled between rolling green hills occupied by sheep (even in February!) – provided the perfect familiarity to foster discussion and exchange of ideas that would lead to collaborations worthy of speeding up discovery and innovation.
After 4 days of intense learning, exquisite meals, and stimulating discussions over drinks, I left inspired and motivated. I feel confident that cerebral organoids will be exponentially improved in the coming years, leading to tremendous advances of our understanding of uniquely-human brain development and its response to environmental perturbation. For a molecular biologist focused on understanding the brain and mental illness, this is an exciting time.
MUKUL TEWARY PhD Student, University of Toronto, Toronto, Ontario, Canada
When I read about this workshop being organized by the Company of Biologists on their website, I immediately sent in an application to attend it. My project deals with developing in vitro models of early development using bioengineering technologies. Although, my graduate studies have focused mostly on investigating the induction of mesendodermal tissues, our latest results have diverted my research interests toward studying the ectodermal lineages including the early specification of neural fates. Given that this workshop was focused on in vitro models of neural fates, and that it gave the attendees opportunities to network with the key opinion leaders in the field, I was extremely excited when I was given the chance to attend it.
This workshop far exceeded all my expectations. First and foremost, the organization of the workshop was exceptional! In terms of the content, I truly enjoyed hearing about the extent of progress that has been made in the field of neural organoids. The format that the organizers had chosen included a daily discussion group where the group discussed concerns that the field has and where they think the field is headed. As an early career scientist, I found these sessions incredibly valuable. Notably, one of the scientific concerns that seemed to be prevalent amongst the group was the variability between different pluripotent stem cell lines in generating the downstream organoids. An important aspect of our latest study deals with this very issue and I was very excited to hear that the key opinion leaders in the field are also looking into the same questions.
Overall, this workshop is one of the best meetings I’ve attended, and I would highly recommend these Company of Biologists workshops to everyone but especially to early career scientists.
MIKE FERGUSON MS student, Biomedical Engineering, Boston University, USA
Between the 3, or was it 5 or 6?, course meals, there was indeed great science being discussed! With armfuls (quite literally) of beers being consumed in the “cozy” backroom, great collaborations were set up well past midnight. Set in a victorian mansion, replete with a full time staff, including a most scholarly house historian, this workshop was truly an experience. If you ever wondered what it would be like to be a member of British high society like Barry Lyndon, look no further.
Biologists talking with engineers was the theme. Despite being an engineer and biologist myself, I was exposed to new ideas and ways of thinking nonetheless. For example, what is development? Is it self-assembly or self-organization? Is it special? The most brief of side conversations offered some of the most interesting ideas. Ideas that kept me thinking well after the workshop and have already caused me to take a fresh look at my own work, with good results.
Perhaps the most unique (and truly invaluable) aspect of the workshop was its laid back nature. Participants were encouraged to present unpublished data (non-disclosure was assured). In many ways, it was like a big informal lab meeting. For the young scientists, the workshop is a unique chance to make your name known and your ideas heard. It was a most interesting look into the future, for which I thank the organizers.
All in all, I left disappointed – this being my first real conference that I have attended, all future conferences and workshops are likely to pale in comparison. How many of them will have staff constantly offering you a cup of tea or coffee (an assortment of treats already laid out in the adjacent parlor)?
SAM NAYLER Postdoctoral Fellow, Department of Physiology, Anatomy and Genetics, University of Oxford, Oxford, UK
What do you get when you take a group of developmental biologists, chemists, bioengineers and neuroscientists away from their daily routines in the lab and put them together in a countryside manor near Brighton, England? The answer is not the punchline to a joke, but instead a seriously productive three days full of introspection and discussion of the pressing issues facing the respective field that unites the interested parties.
The Company of Biologists and conference organisers should be commended on their excellent approach to realizing the formula for a smooth and seamless meeting where inhibitions and impediments to open discussion are rapidly dissolved and a vibrant exchange of ideas and information takes their place. Despite the very formal setting, people were rapidly acquainted and exchanging information and ideas for collaboration. Combined with the fact that everyone presented, and everyone presented unpublished work, allowed for an open and frank forum for discussion. The exposure to fields outside one’s own was an excellent way to survey the current state of play in that field and germinate ideas for collaboration which were later crystallised over a drink in the bar or a walk (or run) in the woods.
There is a proverb that says it takes a village to raise a child. While there is a rich history of developmental biology, the emerging field that uses stem cell science to explore aspects of tissue formation is very much in its infancy, and indeed requires the specialist inputs from the ‘village’ as a whole. To be surrounded by bright, talented and enthusiastic bioinformaticians, biologists and engineers, all of whom have at least temporarily assembled as a village makes me very optimistic about the future of the field going forwards.
A PhD studentship is available in Dr. Claudia Barros’ group, Peninsula School of Medicine, Plymouth University, UK. The project is devoted to characterise the role of novel candidate genes in brain tumour initiation identified in the laboratory. It will focus on investigating pathways acting in mitochondria, the cells’ power plants, but it is not restricted to it. Drosophila will be used to examine candidate genes in the conversion of normal neural stem lineage cells into brain tumour initiation cells via gain and loss of function assays, providing a unique opportunity to unravel their role in live brains at single-cell resolution. Translation of findings will be performed using glioma tumour samples and patient-derived glioblastoma stem cell lines.
We are looking for a candidate with experience in genetics, molecular biology, immunohistochemistry, cell culture, biochemistry and/or confocal imaging. A high interest in neural stem cell and brain tumour biology is a must. Applications are invited from qualified graduates (1st class or 2:1, equivalent), and preferably a relevant Masters or extended laboratory research experience. International students must have an IELTS score of min 7.0 (with no less than 6.5 in any element).
For further information regarding the project, please contact Dr. Claudia Barros (claudia.barros@plymouth.ac.uk). Applications must be made in accordance with the details shown below.
Funding Notes:
The studentship is supported for 3 years and includes full Home/EU tuition fees plus a stipend of £14,553 per annum. The studentship will only fully fund those applicants who are eligible for Home/EU fees with relevant qualifications. Applicants required to cover overseas fees will have to cover the difference between the Home/EU and the overseas tuition fee rates. For the 17/18 academic year the difference in fee is £10350. If you are selected you will be required to provide financial assurances.
Shortlisted candidates will be invited for interview. We regret that we may not be able to respond to all applications. Applicants who have not received an offer of a place by end of May 2018 should consider their application has been unsuccessful on this occasion.
Skin is a fascinating organ that dynamically adjusts its size throughout our lifetimes and does so perfectly while, at the same time, acting as a robust bi-directional barrier protecting us from the insults of the outside world. Of particular interest is the epidermis, which is the outermost, barrier- and load-bearing layer of the skin. Every time we bend our fingers, undergo cycles of weight loss or gain, or are a subject to life-altering events, such as pregnancy, the epidermis has to shrink, expand, and withstand various amounts of tension while still maintaining integrity! From a point of view of an engineer, skin is a marvel – how can this be? What is the structure of this stunning tissue allowing it to perform the many functions that it has?
The epidermis is a highly ordered, multi-layered stratified epithelium. The bottom-most sheet is a tightly packed layer of proliferating stem cells attached to both the underlying matrix as well as to the cells of the terminally differentiated, postmitotic suprabasal layer. Upon differentiation, cells delaminate from the basal stem cell layer and move upwards, filling the many layers of the skin. During this upward motility, the cells need to maintain tight cell-cell junctions in order not to compromise the barrier function of this epithelium. When reaching the most superficial layer, the cells lose their nuclei and dye, forming the dead cornified layer, from where the cells are slowly sloughed off. This turnover is quite rapid, and the entire epithelium is fully renewed every 7-10 days.
The basal layer is the hotspot of most of this activity in the epidermis where some stem cells divide and continue residing within the layer while others differentiate, delaminate, move upwards, and eventually die. In the homeostatic state, epidermis must maintain a constant tissue size and this means a relatively constant basal cell number. Intuitively, the rate of proliferation then must equal to the rate of departure and delamination of the differentiating cells. On a population-level this behavior is completely logical, but the decision-making occurs on cellular level. Then, what are the drivers of cellular fate decisions within a tissue and how do cells coordinate their cell fate with their position? This is the exactly the question that we tackled in our recent publication! It turned out that it is all about differential cell and tissue mechanics!
Let the force guide you – how to robustly couple position with fate
It is intuitively apparent why one may think about external forces, such as compression and tension, in the context of the basal layer of the epidermis as it is an incredibly crowded and mechanically-loaded tissue. It is also instinctive to consider differential cellular tension in the context of cellular extrusion as we now from ample developmental biology literature that cell sorting is largely based on differential cortical tension. To put it simply, it would be more likely for a soft cell to squeeze upwards from a tightly jammed space than it would be for a stiff one. Thus, we started off with the very basic hypothesis that differential cell mechanics would be an excellent cue guiding cells to either remain within the crowded layer or change their position and move upwards. This was especially compelling in the context of highly mitotic areas with highly elevated lateral cellular compression and cell shape deformation. Our hypothesis was that crowding could alter cell shape and cortical tension. Thus, the first question that we asked was whether there was mechanical difference between cells that start differentiating versus ones that remain stem cells within the basal layer. Using atomic force microscopy we observed a biphasic evolution of cortical tension of differentiating cells, where initially upon differentiation cells indeed became softer as they made the decision to detach from the layer and, upon successful escape from the basal layer, they became stiffer, while at the same time changing their adhesion profiles. By measuring adhesion forces we further realized that the more differentiated the cell, the stronger the cell-cell adhesion forces. This was exciting as it suggested that indeed cell fate changes were accompanied by changes in cell mechanics. It further suggested that perhaps the differentiated cell was not only being squeezed out of the stem cell layer, it could also be partially pulling itself out using the strong cell-cell adhesion forces.
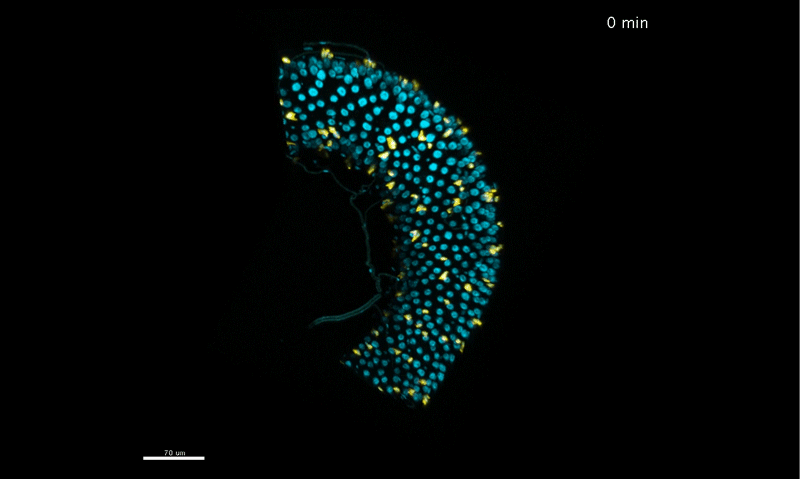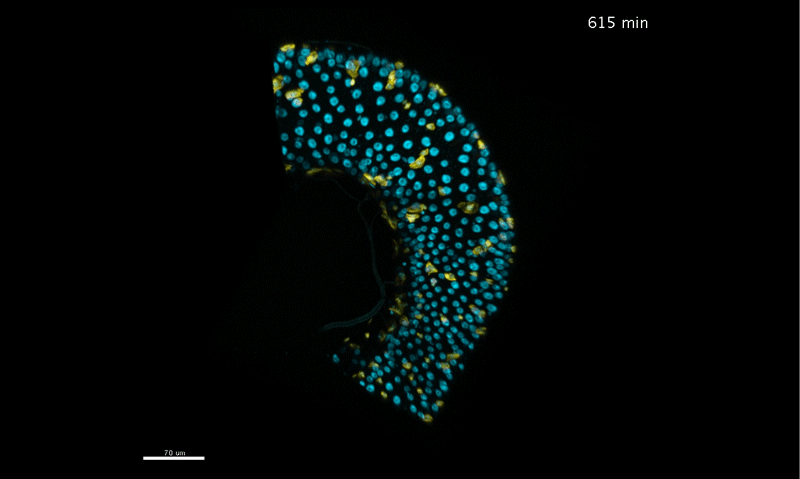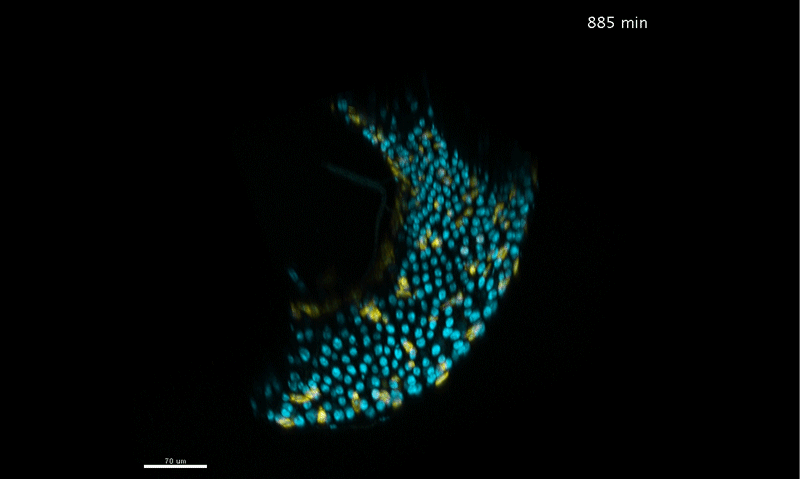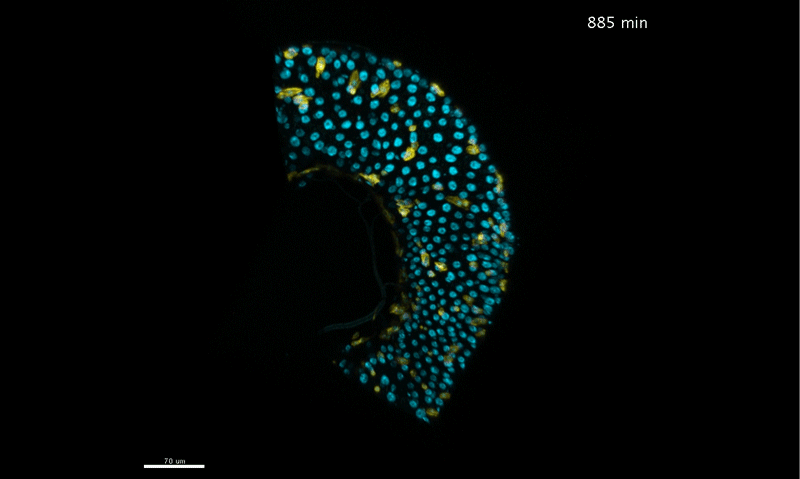
Time-lapse DIC video and smoothed velocity map of EPC monolayers after Ca2 + treatment. Motility vectors (left panel) and smoothed velocity maps (right panel) are shown. Asterisks demarcate examples of 2 dividing cells and arrowhead an example of a delaminating cell. Supplementary Video 3 from Miroshnikova, et al. 2018.
The path to finding answers: when physics meets cell biology
The next step was to ask the classical chicken-and-egg question of what comes first. Does the altered cell mechanics trigger differentiation or does differentiation trigger altered cell mechanics. To this end, we utilized a compression bioreactor as well as micro-patterned surfaces to precisely control crowding and cell shape anisotropy. The answers provided by these experiments made the engineers working on the project quite ecstatic as by artificially inducing crowding or restricting cell adhesion area and forcing cells to assume anisotropic shapes we were able to sufficiently to trigger differentiation. In addition, crowding was further, not surprisingly, sufficient to alter cell mechanics. So indeed, the stem cell layer crowding state seemed to control cell fate, and the cell fate change was coupled to a change in cell mechanics, providing an elegant mechanisms by which the tissue could monitor and react the changes in cell density as well as to couple cell fate to positioning.
As all of the initial observations were done using various in vitro manipulations, we subsequently turned to in vivo imaging of whole embryos to ensure that the in vitro principles are conserved and relevant to the state of the epidermis in vivo. Upon careful visual inspection of the in vivo epidermis, it was immediately apparent that the basal layer existed in a solid-like, jammed state. Interestingly, we subsequently noted that jamming coincided with the onset of delamination in our in vitro monolayers, implying that delamination of differentiating cells could be a specific feature of jammed epithelia. The biological effects of these imaging experiments were clear but their quantification was a bit more of a challenge that we overcame via a fruitful collaboration with Dr. Irene Wang and Dr. Martial Balland from Grenoble, France that allowed us to model tissue strains and monolayer flows in a quantitative manner.
Finally, it was important to test whether crowding-induced stochastic decrease of cortical tension coupled to increased cell-cell adhesion was sufficient to induce delamination. This was challenging, as any chemical or genetic perturbation would affect the mechanics of all cells, whereas we needed to induce mechanical heterogeneity. So to get an initial hint that we were really on to something relevant, we initiated an collaboration with Dr. Joerg Galle and Dr. Torsten Thalheim from Leipzig, Germany who had in the past beautifully simulated intestinal stem cell homeostasis in silico. They set out to simulate monolayer crowding, where increasing cell density through proliferation would, as shown by our experiments, reduce cell surface area, reduce cortical tension and increase cell-cell adhesion.
Time-lapse video of a 3D model epidermis simulation, side view is shown. Supplementary Video 5 from Miroshnikova, et al. 2018.
As a matter of fact, we were blown away by the results that the system maintained epidermal homeostasis at rates comparable to what had previously been observed using live imaging of intact epidermis of mice, and was completely buffered against changes in proliferation rates within the basal layer. Further, only cells with low cortical tension and high cell-cell adhesion delaminated, showing that this could indeed be an extremely robust mechanism to couple cell fate with position. We further went on to validate key aspects of the model by cell mixing experiments in epidermal organotypic cultures, where we observed that single E-cadherin-deficient cells, that showed lower cell-cell adhesion forces and high cortical tension, had severely compromised ability to delaminate, even if they had terminally differentiated.
Taken together, we conclude that individual cells tune their fates based on the activities of the neighboring cells, and by changing their mechanical properties as part of this fate switch, change their position within the tissue. This is quite remarkable, and we are now thinking about the implications of this phenomenon in terms of tissue pathologies. For example, recent findings by the lab of Dr. Phil Jones demonstrated that sun-exposed skin of healthy individuals contains clonal patches of cells with cancer-causing mutations such as mutations in Ras, but these clones co-exist with normal cells without any perturbations to the physiological functions or structure of the epidermis. Our skin is seems thus to be buffered against perturbations such as local differences in proliferative activity or differentiation rates.
I am grateful for having had the chance to work together in a fantastic team of engineers, physicists, biochemists, and cell biologists and am particularly proud to have been leading a study that employed such a wide spectrum of techniques from in vivo imaging of whole organisms to developing new quantitative algorithms and agent-based simulations to explore and understand the wonders of biology.
This editorial by Katherine Brown and Olivier Pourquié was published in Development on 22/02/18.
We are excited to announce the launch of preLights (https://prelights.biologists.com/), a new service from The Company of Biologists. preLights is a community platform for selecting, highlighting and commenting on recent preprints from across the biological sciences. With this service, we aim to help researchers discover the most interesting and relevant preprints from the growing archive of manuscripts deposited on bioRxiv and other preprint servers.
So what is the rationale behind preLights, and how will it work? While preprints have been around for a long time in the physics community, it was only when bioRxiv (https://www.biorxiv.org/) came on the scene in late 2013 that they began to take off in the biological sciences. Since then, we have seen an almost exponential growth in the number of preprints posted, as more and more researchers recognise the value in making their research available at an early stage, and as journals become increasingly open to considering papers that have already been posted on a preprint server. Here at Development, we quickly changed our policy on preprint deposition after the launch of bioRxiv, and have allowed authors to submit papers that were already available on preprint servers since Spring 2014. Since early 2016, all of the Company’s journals have been open to considering manuscripts deposited as preprints. Not only that, but we actively facilitate posting of submitted manuscripts through our bidirectional transfer portal with bioRxiv (see http://dev.biologists.org/content/news#biorxiv for more details). Overall, The Company of Biologists and Development see a synergy between preprint servers and the more traditional peer review and publishing model we operate. Preprints allow authors to get their work out quickly and readers to access the latest research, pre-publication peer review helps to ensure the rigour and quality of the work, and publication in a field-specific journal like Development helps the community find and digest the most relevant research in their area in an accessible format.
Since 2016, our community blog the Node has played an active role in helping developmental biologists find the most relevant preprints – providing a monthly round-up of the latest preprints in developmental biology and related fields (see https://thenode.biologists.com/tag/preprints/). This initiative has proved hugely popular: the post is generally our most-read post each month, and many people have told us how valuable they find the listing. Moreover, the list is getting longer each month, and is in danger of becoming unwieldy, even with the careful curation from our community manager. Given these trends, we started thinking about what more we could do to help people to navigate the ever-growing preprint literature – across the range of fields covered by the Company’s journals.
The preLighter community as of 22/02/18. Meet them all here
From these discussions, the idea of preLights was born. At its heart, preLights is a community of around 80 researchers, mainly (but not exclusively) postdocs and early stage PIs, many of whom have been nominated by our editors or editorial board members. Their research interests span the range of fields covered by our journals and beyond – from morphogenesis to neuroethology, from autophagy to cancer immunology. Our preLights team members will be selecting each month the preprint or preprints that they feel most worthy of comment, and will provide a personal perspective on why they have chosen each article. Through our online platform, we also hope to encourage other members of the community to comment on those preprints selected by our team, and to engage the authors of the selected preprints in the discussion as well – thus facilitating the exchange of ideas and opinions. Hopefully, this will also help authors improve and revise their papers as they make their way towards formal publication. Although the platform has been designed and will be hosted by The Company of Biologists, we see preLights as a community-run service, where we provide logistical support, but the content is driven by our team of selectors and the broader community.
Posts will be categorised and tagged by topic to make it easy for you to search for those preprints most relevant to you, and we’ll also highlight the most popular posts across all fields to give you a flavour of the latest work that’s attracting attention. We’re also planning to feature content from the preLights site in the journal, further helping the developmental biology community to discover the newest research in the field. And to reassure those of you who have become used to browsing the Node’s monthly list, we’re not planning to stop doing this. Finally, an additional aim is to support and promote our team of selectors – giving them the platform and profile to get their opinions ‘out there’ and helping them to expand their networks, both within their specific fields and beyond.
We are of course aware that commenting on preprints and papers has not really taken off in a big way in the biological sciences. preLights is therefore something of an experiment, but we’ve been hugely encouraged by the enthusiasm with which the idea was met by those we approached as potential contributors. While we hope that preLights will provide a venue where such discussion can happen more freely, we also believe that there is significant value in the selection and highlighting of a subset of preprints that will be of particular interest to our communities. Over time, we expect that the preLights site, and the team of selectors, will evolve – and we welcome your suggestions and feedback on how we can make it better. For now, though, we invite you to browse the first set of posts on preLights and to join in the discussion there. We hope you find this new initiative valuable!
 (4 votes)
(4 votes)
 (3 votes)
(3 votes) (No Ratings Yet)
(No Ratings Yet)