Two postdoc positions (1x bioinformatics and 1x wet lab) are available at University College London. Our lab studies how cell fate choice and proportioning is robust and reproducible, despite the fact that extensive cell-cell variation is generated by noise and stochasticity, which can cause heterogeneous gene network activity. Therefore to understand how cell fate choice and patterning are achieved during development and regeneration, we are trying to determine:
(1) How are the gene networks that regulate differentiation affected by noise and stochasticty?
(2) Do differentiating cells all follow the same path towards the differentiated state?
Until recently, however, it has been impossible to follow the behaviour of entire gene networks in individual cells, or to follow their temporal changes in activity in individual cells as they differentiate along different linages. Single cell gene expression analysis, together with novel computational reconstruction of gene network dynamics provides this opportunity. Therefore, in this project you will use powerful new single cell RNA-sequencing approaches to follow changes in gene expression over time in individual differentiating cells.
You will join a multidisciplinary team led by Professor Chris Thompson, following our move to UCL in September 2017. This work builds upon our recent discovery of a novel robust developmental circuitbased on an ultradian cell fate oscillator. The approaches used in the lab include transcriptomics, functional genomics, molecular genetics, live cell imaging, and mathematical modeling.
Key Requirements
Candidates with extensive experience of using either computational genomic approaches (Position 1) or wet lab approaches (Position 2) to understand the molecular basis of gene networks will be considered. You should currently hold or be about to obtain a PhD in Computational, Cell, Molecular or Developmental Biology.
The post funded by the Wellcome Trust and is available for 18 months in the first instance (with possibility of extension).
(Grade 7, Salary (inclusive of London allowance) £34,635 – £41,864 per annum)
To learn more about our work visit: https://thethompsonlab.wordpress.com
For any queries regarding the application process, please contact Biosciences Staffing on biosciences.staffing@ucl.ac.uk and quote the reference 1670534
Hello readers of the Node! My name is Sarah and I’m delighted to meet the Node readership as the new intern on the Node, hopefully I will be communicating and interacting with many of you over the next three months.
Please allow me to introduce myself and tell you a little bit about myself. I am currently undertaking a BBSRC PhD in Dr Thomas Pratt’s research lab at the University of Edinburgh. A component of this programme is a three-month internship to allow us to experience careers outside the traditional academic job trajectory and I am fortunate enough to undertake my internship with the Node. My background is developmental neurobiology, with my PhD focusing on understanding the concert of genes that act to control human foetal cerebral cortex development and how disruptions to this can contribute to neurodevelopmental disorders.
For years now I have been fascinated by the incredible complexity of all aspects of development and how the multitude of processes acts flawlessly in concert to generate the complete organism. The Node is a place that encompasses the vast spectrum of development; from the computational modelling, single cell studies, to complete organisms into one space for biologists to learn, explore new avenues of research and simply enjoy the wonder of it all. I am so excited to be a part of this online community for all researchers and I hope I can contribute to making it a more enjoyable resource for you all.
Faculty of Health and Medical Sciences, University of Copenhagen
Background
The Novo Nordisk Foundation Center for Stem Cell Biology – DanStem has been established as a result of a series of international recruitments coupled with internationally recognized research groups focused on insulin producing beta cells and cancer research already located at the University of Copenhagen. DanStem addresses basic research questions in stem cell and developmental biology and has activities focused on the translation of promising basic research results into new strategies and targets for the development of new therapies for cancer and chronic diseases such as diabetes and liver failure. Find more information about the Center at http://danstem.ku.dk/.
Job Description We are looking for a postdoctoral candidate with a strong developmental biology/cell signaling background and/or a biological image processing background to study the cellular and molecular mechanisms of lumenogenesis and tubulogenesis and their role in balancing cell proliferation and differentiation of multipotent progenitors in the developing pancreas. The candidate is expected to use both in vivo (mouse) and in vitro (human pluripotent stem cells) experimental models. The position is for 2 years with possible extension. The employment is planned to start December 1st 2017 or upon agreement with the chosen candidate.
Qualifications
The candidate is required to hold a PhD degree in stem cell/developmental/cell biology/biological image processing. A few years of postdoctoral experience in the same areas is a merit. A candidate with biological background should also have hands on experience in mouse genetics, micro dissection of mouse embryonic organs, human pluripotent stem cell culture and differentiation, live-cell imaging, quantitative image analysis. A candidate with image processing background should have experience with segmentation and tracking in large 3D datasets and an interest in learning to work with the biological techniques described above. Finally, we are looking for applicants with a good record of peer reviewed scientific publications and grant writing skills.
Employment Conditions The terms of employment are set according to the Agreement between the Ministry of Finance and The Danish Confederation of Professional Associations or other relevant professional organization. The position will be at the level of postdoctoral fellow and the basic salary according to seniority is 32.700-34.400 DKK/month. A supplement could be negotiated, dependent on the candidate´s experiences and qualifications. In addition a monthly contribution of 17.1% of the salary is paid into a pension fund. Applicants that are recruited from abroad may be eligible for a special researcher taxation scheme. In all cases, the ability to perform the job will be the primary consideration, and thus we encourage all – regardless of their personal background and status – to apply.
Foreign applicants may find the following links useful: www.ism.ku.dk (International Staff Mobility) and www.workingconditions.ku.dk.
Application Instruction The application must be submitted in English, by clicking on “Apply online” below. Only online applications will be accepted. The closing date for applications is 23.59pm, November 1st 2017.
The application must include:
Cover letter detailing the basis on which the applicant scientific qualifications meet the requirements for this position.
Curriculum vitae.
List of references (full address, incl. email and phone number)
Diplomas – all relevant certificates.
List of publications.
Application procedure
After the expiry of the deadline for applications, the authorized recruitment manager selects applicants for assessment on the advice of the Appointments Committee. All applicants are then immediately notified whether their application has been passed for assessment by an expert assessment committee. Selected applicants are notified of the composition of the committee and each applicant has the opportunity to comment on his/her assessment. You may read about the recruitment process at http://employment.ku.dk. DanStem and The University of Copenhagen wish to reflect the diversity of society and welcome applications from all qualified candidates, regardless of personal background.
Founded in 1479, the University of Copenhagen is the oldest university in Denmark. With 37,000 students and 9,000 employees, it is among the largest universities in Scandinavia and one of the highest ranking in Europe. The University consists of six faculties, which cover Health and Medical Sciences, Humanities, Law, Science, Social Sciences and Theology. Part of the International Alliance of Research Universities (IARU), and among Europe’s top-ranking universities, the University of Copenhagen promotes research and teaching of the highest international standard. Rich in tradition and modern in outlook, the University gives students and staff the opportunity to cultivate their talent in an ambitious and informal environment. An effective organisation – with good working conditions and a collaborative work culture – creates the ideal framework for a successful academic career.
Molecular biological approaches to understanding the complex gene networks regulating development often resemble attempts to understand a Monet painting by looking at it up close. We find the links within a disorganized array of brush strokes and shapes but may miss the big picture. Our recent study is an example of how one must sometimes take few steps back to admire the water lilies.
Why do we study inner ear development?
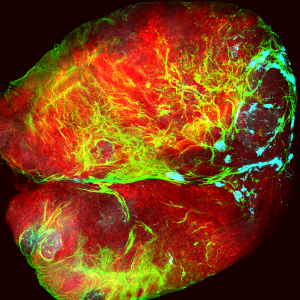
The subject of our investigation is a vestibular sensory organ—the utricle (Fig. 1). Our ability to perceive accelerations and to orient ourselves in three-dimensional space relies upon the operation of this and four other vestibular organs within the inner ear (Utricle and Saccule balance and equilibrium). Despite the differences in their functions, the vestibular organs as well as the hearing organ, or cochlea, are populated by the same type of mechanosensitive receptors—the hair cells. Mammals develop a specific number of these cells during embryonic development; no additional hair cells are formed throughout life, and once lost, these sensory receptors cannot be restored naturally. Hair cells are vulnerable to many environmental and health hazards that are prevalent in industrialized countries: loud noises, certain antibiotics, and anticancer drugs. As a result, deficiencies of hearing and balance often occur owing to the irreversible loss of hair cells. Regeneration of the sensory receptors represents a promising approach for treating these disorders. However, understanding the molecular basis of sensory organ development is a necessary first step towards making this treatment a reality.
Figure 1. In this preparation of the sensory portion of the utricle, hair cells are labeled in green. Supporting cell nuclei and a subset of the hair cell nuclei are labeled in red.
The original idea for our work
The size of each of the inner ear sensory organs, such as the utricle, shows miniscule variation, suggesting a tight control of growth. Whereas many molecular pathways that stimulate sensory cell proliferation during early development have been identified, the growth control mechanism that arrests proliferation in these organs has not been understood. By combining computational modeling and molecular biology we sought to investigate this mechanism in the utricle (Gnedeva et al., 2017).
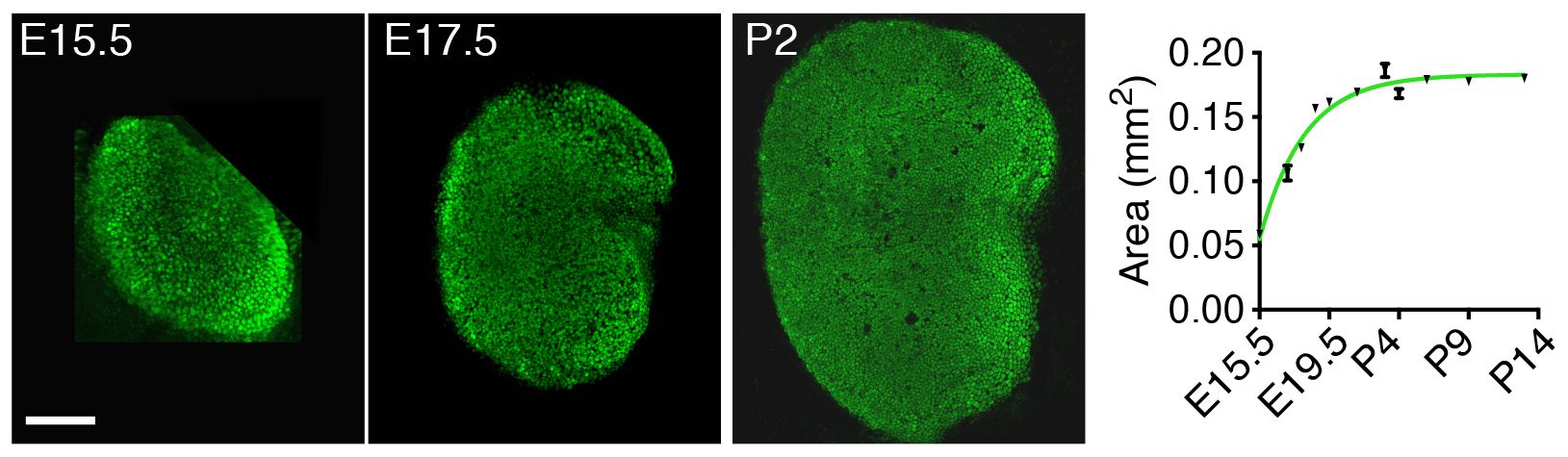
Instead of looking first at the molecular, “up close” level by investigating the gene-expression changes accompanying development of the utricle, we decided to take a few steps back and to apply a macroscopic, engineering approach to understand the mechanisms controlling its growth. By measuring the changes in area of the utricle during development starting at embryonic day 15.5 (E15.5), we found that the organ’s growth slows as the utricle approaches its final size by postnatal day 2 (P2) (Fig. 2). In other words, the rate of the organ’s growth correlates negatively with its size. This observation suggested a self-regulatory mechanism of epithelial growth and implied a negative-feedback mechanism—a previously unexplored possibility of growth control in sensory organs of the inner ear.
Figure 2. In this preparation of the sensory portion of the utricle, supporting cells are labeled in green (left panels). A decrease in the slope of the curve representing the area of the utricle as a function of time (graph on the right) demonstrates that the growth rate decreases as the organ approaches its final size.
Surprisingly, in biological systems only two major self-regulatory negative feedback mechanisms are known to regulate organ growth. The first was discovered when in vitro studies demonstrated that many cell types produce morphogens that suppress their own proliferation. One of the first examples of such molecules is a member of the TGF-β family of morphogens—myostatin, which is produced by skeletal muscles to inhibit their growth. Many homologous molecules have since been discovered to control growth in various other tissues.
The second mechanism of growth control is related to mechanical constraints. It has long been observed that cells cease to proliferate in culture when they fill up the available space. The molecular basis of this phenomenon, called contact inhibition of cell growth, has recently been discovered. In brief, cell-cell contacts, as well as changes in pressure and tissue stiffness, can activate the Hippo signaling pathway. The Hippo kinase cascade subsequently targets the transcriptional cofactors Yap and Taz, resulting in their degradation; this in turn inhibits cell growth and proliferation.
In our work we created morphogen-limited and elasticity-limited computational models to incorporate each of the two growth control mechanisms and to explore which one of them is more dominant in regulating cell proliferation in the utricle.
What did we discover?
Because the models included the negative-feedback control of growth, they could both accurately reproduce the observed pattern of utricular area expansion. To distinguish between the models, we developed an ex vivo culture system that allowed for manipulation of the elastic force independently of the morphogen concentration in the system.
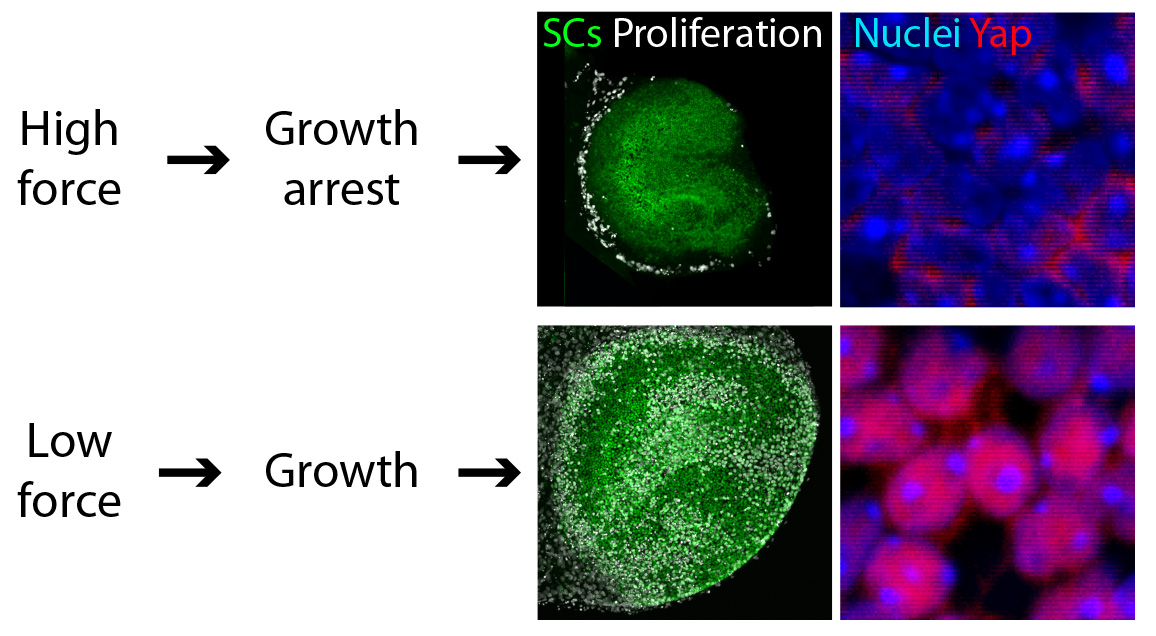
We found that reducing the mechanical constraints normally present during development led to a striking expansion of utricular area (Fig. 3). This demonstration that elastic force retards the growth of the utricle suggested the involvement of the Hippo signaling cascade. Our experimental data confirmed this prediction: the excessive growth observed in low-stiffness conditions was triggered by the accumulation and nuclear translocation of Yap protein—the direct target of Hippo signaling (Fig. 3).
Figure 3. Force controls cell proliferation in the utricle. In these preparations of the sensory portions of utricles (top panels), supporting cells (SCs) are labeled in green and proliferating cells are labeled in white. The supporting cell nuclei of growth-arrested utricles, which have been subjected to mechanical constraints, show limited cytoplasmic Yap staining (bottom left panel). In contrast, an actively growing utricle with low mechanical constraints demonstrates nuclear Yap localization (bottom right panel).
We confirmed that the pattern of cell proliferation normally observed in the developing utricle can be explained by the distribution of the forces in the tissue, as predicted by the model, and accords with the expression pattern of Yap protein. Importantly, we also found that the role of Yap is not limited to development; nuclear Yap translocation is also induced during the limited regeneration observed in the utricle. Because cell proliferation is a first step necessary for recovery of the inner ear sensory functions, we predict that altering Hippo signaling might induce regeneration. The strength of this approach, as opposed to gene therapy, is that the kinases in the Hippo pathway can be targeted pharmacologically. Although further investigation of the role of Hippo signaling in the inner ear is required, biochemical manipulation of this pathway might aid in the recovery of hearing and balance after the loss of hair cells.
In conclusion
Methodological advances during the past few decades have progressively specialized and separated the branches of biological research. Although this specialization has led to great progress in our understanding of living matter at a deeper level, the separation has come at a cost. It is increasingly clear that a cross-disciplinary approach can restore a broader perspective and facilitate the investigation of complex biological systems. Our work constitutes another example of the benefits of such an approach. A combination of mathematical and biological methodologies has allowed us to uncover a growth control mechanism regulating the development of sensory epithelia in the inner ear. Our hope is that building upon our original model, for example by adding the processes of cell differentiation and cell death, can yield an understanding of other aspects of inner ear development, such as the mechanism of pattern formation and regeneration in the system.
All the illustrations are adopted from the original article:
Gnedeva K.*, Jacobo A.*, Salvi J., Petelski A., and Hudspeth A. J. Elastic force restricts the growth of the murine utricle. eLife in press (2017). DOI: 10.7554/eLife.25681
* These authors contributed equally to the manuscript
We have openings for 2 motivated postdoctoral fellows with strong Drosophila molecular genetics experience and interest in integrative strategies to join our team.
Project 1. miRNA biology in the nervous system. Although miRNAs are often thought to mediate “fine-tuning”, we revealed many examples of profound defects in neural miRNA mutants (e.g. Garaulet J Neuroscience 2016, Sanfilippo Development 2016, Sun PLoS Genetics 2015, Garaulet Developmental Cell 2014). We uncovered other developmental and behavioral defects in neural miRNA mutants and utilize genetic, neurobiological and genome engineering strategies to precisely characterize their critical in vivo biologies.
Project 2. siRNA biology in the testis. What is endogenous RNAi utilized for? Surprisingly, there is still little clear biology. Building on our recent appreciation that Drosophila RNAi mediates adaptive gene regulation and productive spermatogenesis (e.g. Wen Molecular Cell 2015), we currently explore the evolution and function of RNAi systems across the Drosophilid phylogeny. We discovered networks of rapidly evolving RNAi substrates in the testis, and are using genomics and CRISPR/Cas9 in non-model fruitfly species to reveal critical roles of RNAi that cannot be seen in D. melanogaster, and may underlie speciation.
Funded positions with housing and medical benefits are available immediately. Please provide CV, motivation letter and references to Eric Lai, laie@mskcc.org.
Hello, Community! In July, I revealed to the world my attempts at scientific illustration. Aidan (The Node’s Community Manager) took a keen interest in my work and was very kind to share it on social media. Here is my long-promised post about my biology art (which can be viewed at www.embryosafari.com).
Before I delve into the illustrations, let me introduce some background information about myself:
1st Instar (Science). Like many of us, I have been enthralled by biology since an early age. The specific focus of this fascination fluctuated through the years (from paleontology, through molecular virology to immunology), but I finally settled on developmental biology (in October 2006, to be precise), as I saw it was the science which tackled the most wonderful of all subjects: how bodies form (and evolve). Eventually, this lead to my PhD in developmental genetics in the labs of Anne-Gaelle Borycki and Phil Ingham, where I studied the expression of an ECM (extracellular matrix) gene using mouse, chicken and zebrafish embryos.
2nd Instar (Art). Similar to my fascination with science, I have enjoyed art (particularly clay modelling and pencil sketching) for as long as I can remember. As a child, I used to grab a block of clay and start sculpting (with arguable success) sci-fi creatures and characters that had grabbed my imagination on screen. Unlike science, I have never had any formal training in the arts (I don’t count art classes in primary school as such). In my later years, when I was already studying embryos, I frequently resorted to clay modelling when I needed to understand the three-dimensional anatomical organisation of a body region or organ. Subsequently, this “hobby” evolved into a digital form, the results of which I present below.
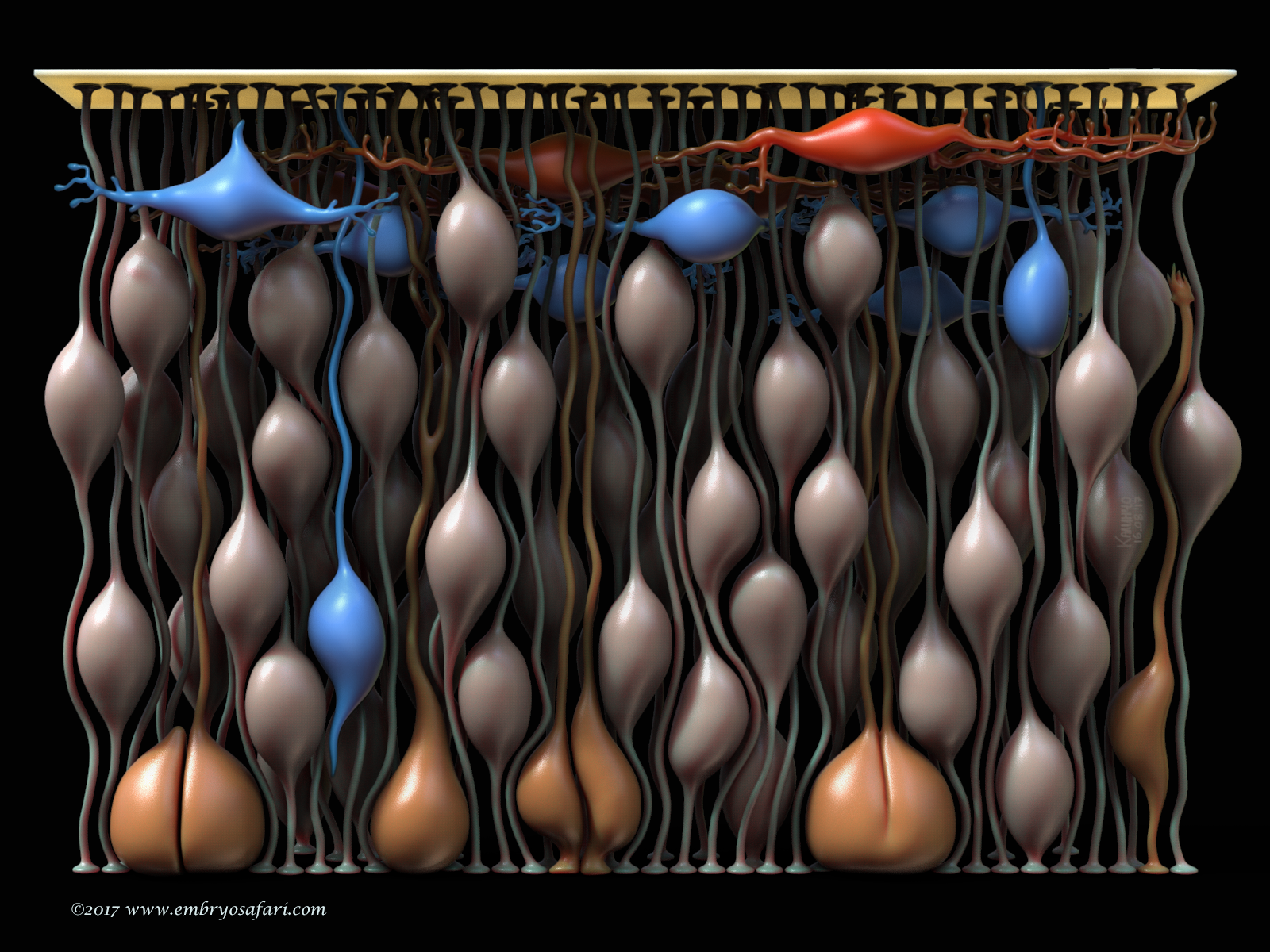
“Building The Brain”
This work represents the cytoarchitecture of the murine telencephalon at E10.5. The goal was to show the cell types at the beginning of neurogenesis. The fleshy (salmon) colour indicates the neuroepithelial cells (NECs) whose nuclei migrate “up and down” the thickness of the epithelium, undergoing INM (interkinetic nuclear migration). Mitoses occur close to the ventricle (the apical side of the epithelium), where the mitotic cells are indicated in orange-ish. What fascinates me is the fact that most NECs, even the mitotic ones, remain anchored to the pial basal lamina (the bright yellow plate on the top) via these long, thread-like basal processes. The first neurons in the telencephalon, those of the preplate, are indicated in red (Cajal-Retzius cells) and blue (prospective subplate neurons). Initially, this model was part of a bigger story which aimed to illustrate gyrification (cortical folding) in mammals, but it was too ambitious (one day I’ll go back to it!).
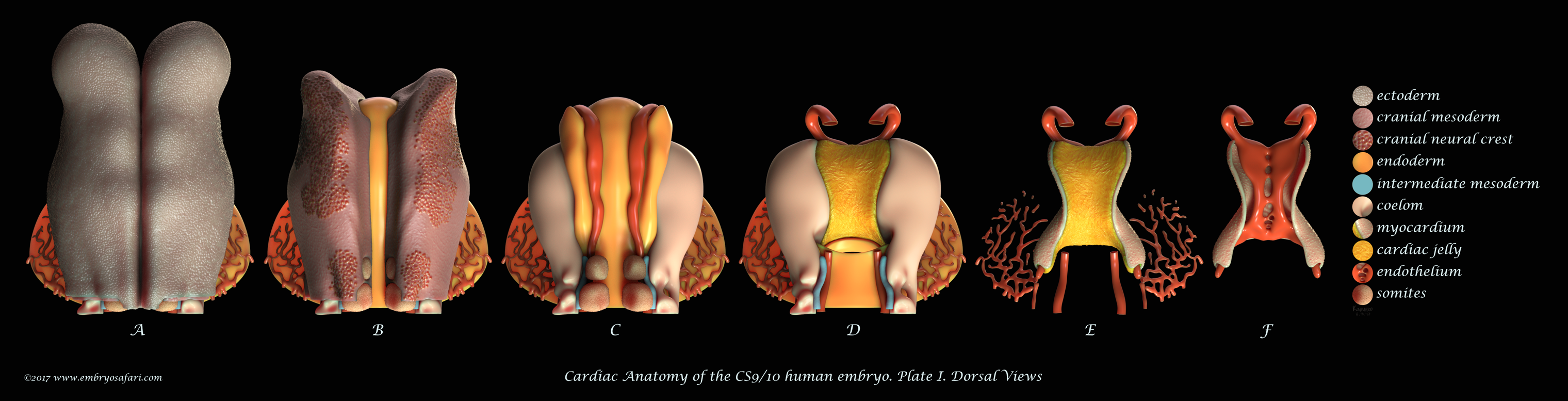
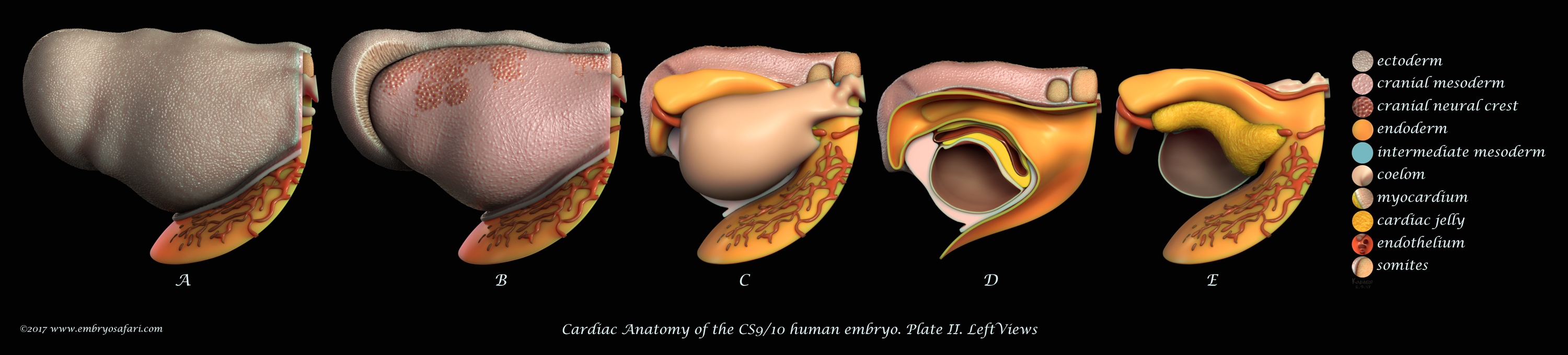
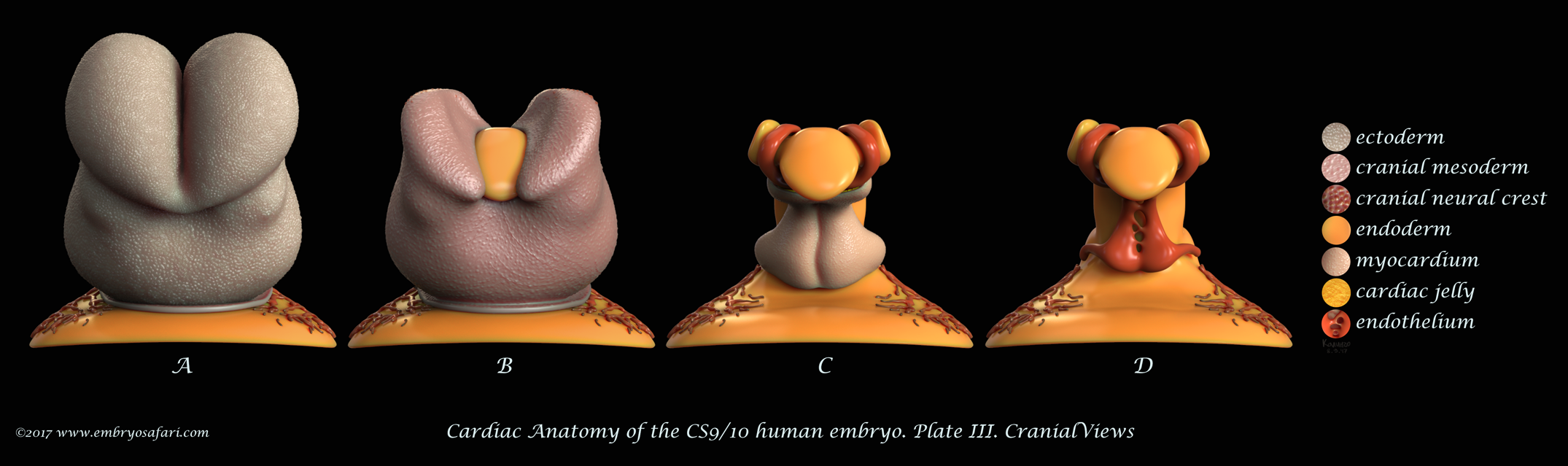
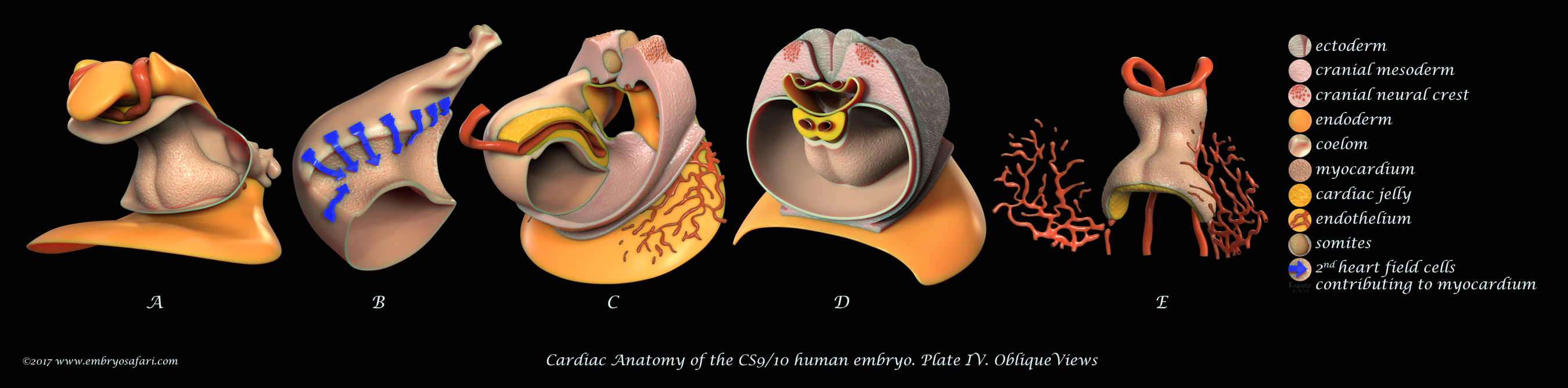
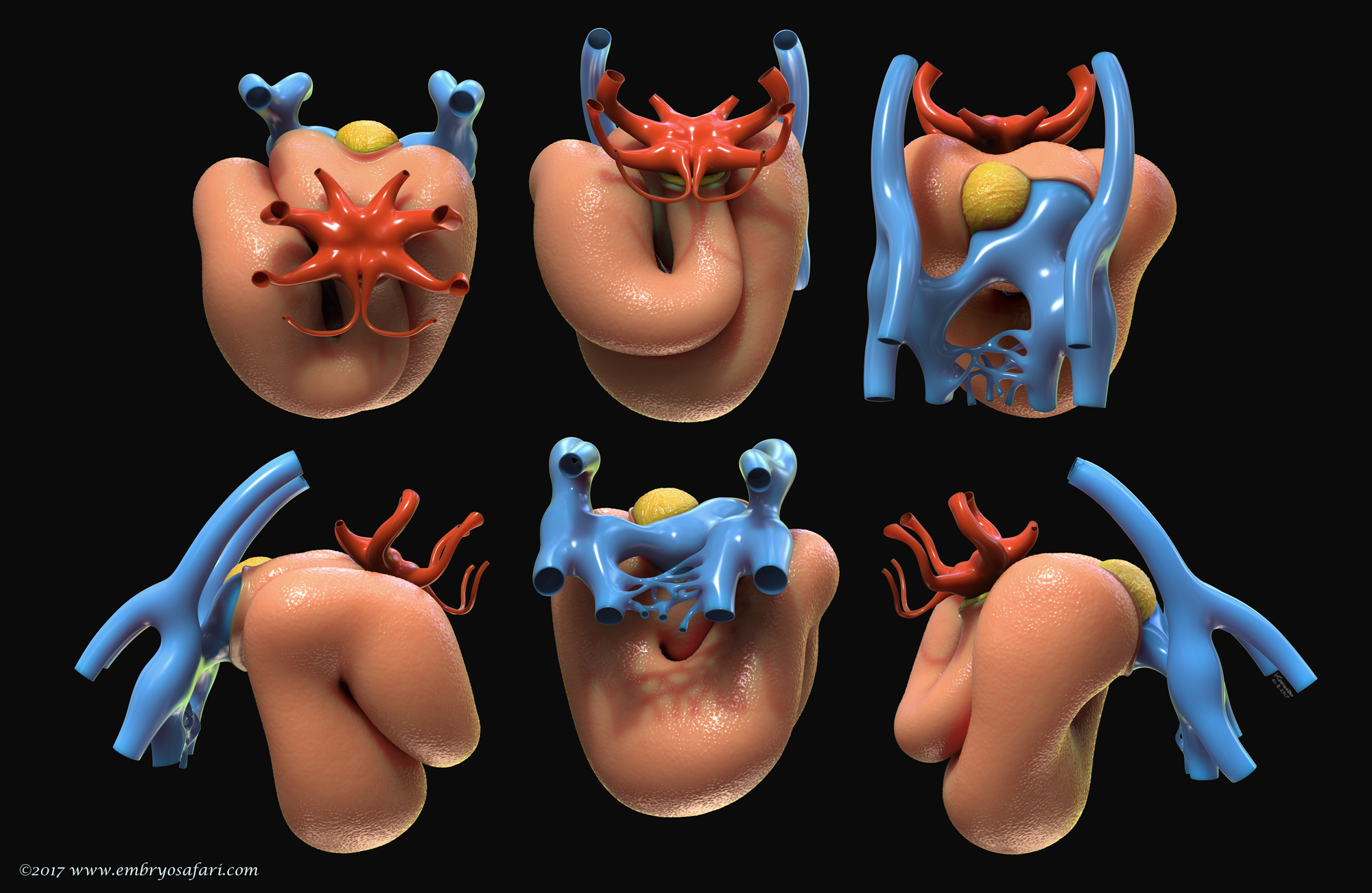
Rarely there is a subject more irresistible than the embryology of our own species. Without any anthropocentric bias, human developmental anatomy is simply beautiful! Of all human embryology subjects, I consider brain, cardiac and craniofacial development most challenging in terms of morphological understanding. And because I enjoy such kind of challenges, I decided to tackle first cardiac anatomy, for which I knew the least. Ideally, I dream of creating illustrated plates for all Carnegie Stages, for both external and internal cardiac morphology, as well as for the major cellular and genetic events in cardiac development. Only time will show how feasible such an endeavour is. For now, I present illustrations on Carnegie Stages 9/10 and 12.
These plates illustrate the anterior half of the human embryo at a morphological stage between CS9 and CS10, viewed from different angles. This stage is interesting for it shows the fusion of the initially bilateral endocardial tubes (visible in Figure J on Plate I), and the presence of a distinct pericardial cavity (anterior coelom), shown in Figures C, D (Plate I) and in A-D (Plate IV). The abundant cardiac jelly, visible in Figure E (Plate I), and in Figures C and D (Plate IV), fills the space between the fusing endocardial tubes and the incipient myocardium. Importantly, myocardial progenitors in the second heart field add up to the growing myocardium (shown as blue arrows in Figure B on Plate IV). In addition to the heart, I spent some time sculpting the closing neural tube, the first two pairs of somites, the septum transversum, and the cranial neural crest (each cell was individually painted, as seen in Figure B on Plate I). For detailed examination, please view the images at full screen mode, or visit https://www.embryosafari.com/gallery.
This illustration represents the external morphology of the embryonic human heart at Carnegie Stage 12, viewed from different angles. The pericardial sac has been removed to expose the myocardium. The focus here is on the spatial relationship between the myocardium (salmon colour), the cardiac jelly (in yellow) and the vessels at the venous pole (blue). At this site, the second heart field continues supplying the myocardium with cardiomyocyte progenitors via the dorsal mesocardium (the cut edge in the top 3rd figure, left to right). The red colour indicates the vessels at the arterial pole of the heart, where the highly dynamic pharyngeal arch arteries form and regress in a quick, craniocaudal succession.
At the end, I would like to thank all curious, luminous and hardworking beings of science, without whom developmental biology wouldn’t have bloomed into its current state.
We are looking for a postdoctoral candidate with a strong developmental biology/cell signaling background and/or a biological image processing background to study the cellular and molecular mechanisms of lumenogenesis and tubulogenesis and their role in balancing cell proliferation and differentiation of multipotent progenitors in the developing pancreas.
Background:
The Novo Nordisk Foundation Center for Stem Cell Biology – DanStem has been established as a result of a series of international recruitments coupled with internationally recognized research groups focused on insulin producing beta cells and cancer research already located at the University of Copenhagen. DanStem addresses basic research questions in stem cell and developmental biology and has activities focused on the translation of promising basic research results into new strategies and targets for the development of new therapies for cancer and chronic diseases such as diabetes and liver failure. Find more information about the Center at http://danstem.ku.dk/.
Job Description:
We are looking for a postdoctoral candidate with a strong developmental biology/cell signaling background and/or a biological image processing background to study the cellular and molecular mechanisms of lumenogenesis and tubulogenesis and their role in balancing cell proliferation and differentiation of multipotent progenitors in the developing pancreas. The candidate is expected to use both in vivo (mouse) and in vitro (human pluripotent stem cells) experimental models. The position is for 2 years with possible extension. The employment is planned to start December 1st 2017 or upon agreement with the chosen candidate.
Qualifications:
The candidate is required to hold a PhD degree in stem cell/developmental/cell biology/biological image processing. A few years of postdoctoral experience in the same areas is a merit. A candidate with biological background should also have hands on experience in mouse genetics, micro dissection of mouse embryonic organs, human pluripotent stem cell culture and differentiation, live-cell imaging, quantitative image analysis. A candidate with image processing background should have experience with segmentation and tracking in large 3D datasets and an interest in learning to work with the biological techniques described above. Finally, we are looking for applicants with a good record of peer reviewed scientific publications and grant writing skills.
Employment Conditions:
The terms of employment are set according to the Agreement between the Ministry of Finance and The Danish Confederation of Professional Associations or other relevant professional organization. The position will be at the level of postdoctoral fellow and the basic salary according to seniority is 32.700-34.400 DKK/month. A supplement could be negotiated, dependent on the candidate´s experiences and qualifications. In addition a monthly contribution of 17.1% of the salary is paid into a pension fund. Applicants that are recruited from abroad may be eligible for a special researcher taxation scheme. In all cases, the ability to perform the job will be the primary consideration, and thus we encourage all – regardless of their personal background and status – to apply.
For further information contact Professor Henrik Semb, henrik.semb@sund.ku.dk
Foreign applicants may find the following links useful: www.ism.ku.dk (International Staff Mobility) and www.workingconditions.ku.dk.
Application Instruction:
The application must be submitted in English, by clicking on “Apply online” below. Only online applications will be accepted. The closing date for applications is 23.59pm, November 1st 2017.
The application must include:
Cover letter detailing the basis on which the applicant scientific qualifications meet the requirements for this position.
Curriculum vitae.
List of references (full address, incl. email and phone number)
Diplomas – all relevant certificates.
List of publications.
Application procedure:
After the expiry of the deadline for applications, the authorized recruitment manager selects applicants for assessment on the advice of the Appointments Committee. All applicants are then immediately notified whether their application has been passed for assessment by an expert assessment committee. Selected applicants are notified of the composition of the committee and each applicant has the opportunity to comment on his/her assessment. You may read about the recruitment process at http://employment.ku.dk. DanStem and The University of Copenhagen wish to reflect the diversity of society and welcome applications from all qualified candidates, regardless of personal background.
Apply online
Link for general info on the recruitment process: http://employment.ku.dk/faculty/recruitment-process
Founded in 1479, the University of Copenhagen is the oldest university in Denmark. With 37,000 students and 9,000 employees, it is among the largest universities in Scandinavia and one of the highest ranking in Europe. The University consists of six faculties, which cover Health and Medical Sciences, Humanities, Law, Science, Social Sciences and Theology.
Applications are invited for the new position of Postdoctoral Scientist in Quantitative Biology of Cell Fate and Tissue Dynamics.
We seek a highly motivated, proactive individual who will benefit from the exceptional WIMM research environment and the recently purpose-built MRC WIMM Centre for Computational Biology, which brings together experts in analysis and modelling of large biomedical datasets. Working under the supervision of Dr Ed Morrissey, you will play a key role in developing mathematical and statistical methods to interpret intestinal stem cell data. You will work in collaboration with the experimental biology group of Dr Doug Winton, based at CRUK-Cambridge Institute. Your work will include developing analytical tools and resources to better understand stem cell dynamics within the intestinal tissue, both in healthy tissue and in the early stages of carcinogenesis.
You will have a PhD in a quantitative discipline (e.g. computational biology, physics, statistics, engineering or mathematics), with experience of working in a UNIX/LINUX environment. You will be proficient in the use of at least one programming language e.g. c++, python, R or MATLAB. A prior publication record with evidence of key involvement in work and a strong interest in applying modelling to biological questions are also essential.
The position, available immediately, is funded by the Wellcome Trust and is fixed-term for 2 years.
Applications for this vacancy are to be made online. To apply for this role and for further details, including a job description and selection criteria, please click on the link below:-
Tumour (red) covered by collagen (green), which is being deposited by cells of the immune system (cyan). (Image: Mariana Muzzopappa, IRB Barcelona)
Researchers from the Development and Growth Control Laboratory at IRB Barcelona have identified the cell types and molecular mechanism responsible for the unlimited growth potential of epithelial tumours (carcinomas) and demonstrated that the growth of these tumours is independent of its microenvironment. “In epithelial tumours caused by chromosomal instability or loss of cell polarity, the interaction between two tumour cell populations drives malignant growth,” explains Marco Milán, ICREA Research Professor and head of the laboratory.
Published in the journal Proceedings of the National Academic of Sciences (PNAS) as a cover story of its 35 Issue, the study analyses solid tumours of epithelial origin in the fruit fly Drosophila melanogaster. “We have induced tumour development in two ways—by generating genomic instability and the loss of cell polarity. We have validated the causal relation between these two conditions—which are frequently observed in carcinomas—and the development of tumours,” explains Mariana Muzzopappa, first author of the study and postdoctoral fellow in the Development and Growth Control Lab.
Independent of the tumour microenvironment
To study the effect of the microenvironment on tumour development, the researchers examined tumour growth in the absence of adjacent cell populations, such as cells of the immune system or mesenchymal cells, which can act as a niche by supplying tumours with growth factors. The scientists observed that the tumour continued to grow in the absence of these two cell types.
Furthermore, they demonstrated that “the growth of epithelial tumours is dependent on activation of the JNK stress signalling pathway and that this pathway is intrinsically activated in the tumour, regardless of its microenvironment,” highlights Marco Milán.
Interactions between two tumour cell populations
The researchers have identified two functionally distinct cell populations within the tumour—one that proliferates and ones that does not—upon which internal growth mechanisms depend. “JNK is activated in a group of non-proliferating cells, namely those that show the highest degree of chromosomal instability or that have lost polarity. JNK triggers the expression of growth factors and makes those cells still in the epithelium to go on proliferating. The continued proliferation of these cells leads to an increase in chromosomal instability and the loss of epithelial polarity in the tumours. Consequently, the number of cells expressing growth factors rises. These cross-feeding interactions explain the unlimited growth potential of these epithelial tumours,” says Muzzopappa.
The mechanism of JNK activation differs depending on the tumour. “We have observed that tumours derived from chromosomal instability are induced by oxidative stress caused by ROS (reactive oxygen species), which triggers JNK. The mechanism in tumours that arise from the loss of cell polarity differs,” explains Marco Milán.
The results of this study shed further light on the causal relationship between chromosomal instability, loss of epithelial polarity and tumorigenesis and open new avenues for the search of therapeutic targets.
This study was supported by the Ministry of Economy, Industry and Competitiveness (MINECO) through ERDFs “A way to build Europe” and “la Caixa” Foundation PhD programme, that supports Lada Murcia PhD student.
Reference article:
Mariana Muzzopappa, Lada Murcia and Marco Milán
Feedback amplification loop drives malignant growth in epithelial tissues
Positions for Students (MSc and PhD) and Postdoctoral Research Fellows in Cardiovascular Genetics
Positions are available in the laboratory of Dr. Gregor Andelfinger at the CHU Sainte Justine Research Center, Montréal, Québec, Canada at MSc, PhD and postdoctoral level. Our work focuses on the genetic origins of cardiovascular diseases in the young and includes both genomics and functional approaches. In these projects, the successful candidates will apply genomic and molecular biology approaches to develop cell‐ and animal‐based disease models. Successful candidates will have a unique opportunity to do interdisciplinary research at all interfaces of developmental and translational medicine and publish their results in high impact journals.
We are looking for highly motivated and focused candidates who will work in collaboration with all other lab members. Team players with excellent communication skills who want to pursue a career in basic or translational cardiovascular research will be ideal candidates. We require expertise (post‐doctoral fellows) or willingness to learn (PhD and MSc students) a wide variety of basic and cutting‐edge techniques (histology, microscopy, developmental studies, Next Generation Sequencing including single cell, stem cell assays, CRISPR, and bioinformatics).
Applicants at MSc or PhD level must have excellent academic standing. Applicants for postdoctoral fellowships must have a recent Ph.D. degree and publications in life sciences, cell biology, or a related field.
The CHU Sainte Justine Research Center has recently moved into a brand‐new building and has all the required state‐of‐the‐art infrastructure and a dynamic international scientific community. Our Research Center is directly linked with Sainte Justine University Hospital Center, a very large tertiary pediatric center. The PI is affiliated with Université de Montréal, one of the largest universities in Canada and the largest francophone university in North America. Montreal is a vibrant bilingual city with a high quality of life. With a greater metropolitan area of 3.5 M, Montreal offers all scientific and cultural benefits of a large North American agglomeration with a European flair and beautiful surroundings.
Interested applicants are invited to provide the following documents to Dr. Gregor Andelfinger (gregor.andelfinger@recherche‐ste‐justine.qc.ca):
CV
Cover letter describing previous experience and career goals
 (No Ratings Yet)
(No Ratings Yet) (7 votes)
(7 votes)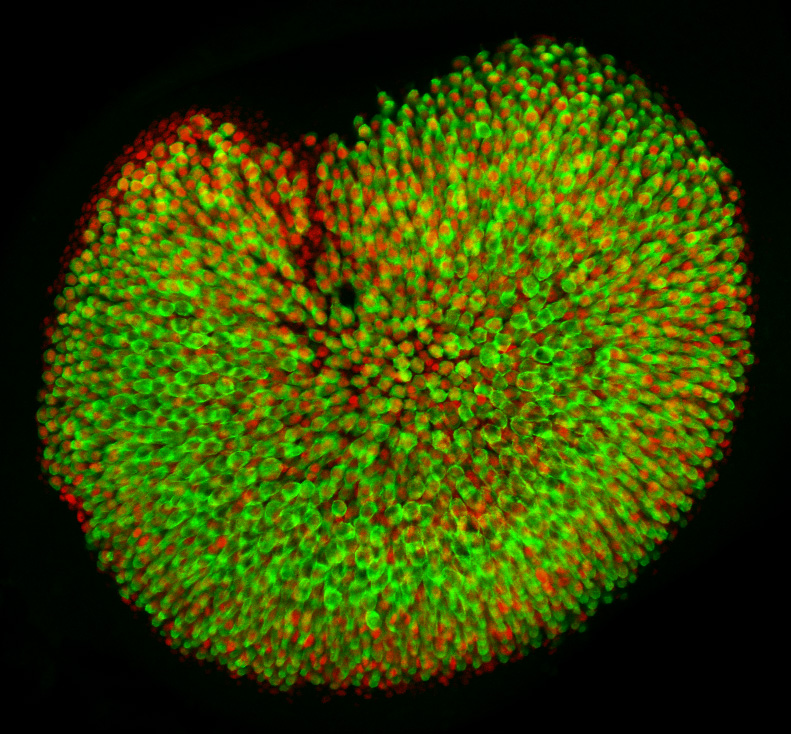
 (4 votes)
(4 votes)