Department of Biology, Ghent University, Krijgslaan 281 S8, 9000 Ghent, Belgium
VIB-UGent Center for Plant Systems Biology, Technologiepark 927, B-9052 Ghent, Belgium
Department of Plant Biotechnology and Bioinformatics, Ghent University, Technologiepark 927, B-9052 Ghent, Belgium
Complex multicellular life has evolved from unicellular organisms along at least five independent paths, giving rise animals, plants, fungi, red algae and brown algae, respectively. Asymmetric divisions are key in this process as a means to create diverse cell types. At the molecular level, the uneven distribution of molecular components such as mRNA, proteins or organelles determines a polarisation vector. A subsequent cell division along this polarization vector results in two daughters cells which are no longer equivalent and receive different cell fates.
In recent years, insights in the molecular mechanisms determining cell polarisation and asymmetric cell division have mainly emerged from investigations on animals. With exception of yeasts, the mechanisms of polarity establishment are much less well established in other groups of eukaryotes. While many model cell polarity systems from land plants lend them to the research of polarity signalling, polarity is often either predetermined (e.g. pollen) or the cells are enclosed in the surrounding tissue (e.g. zygotes), which renders them less suitable for studies examining polarity establishment. Conversely, eggs of brown algae are released in the surrounding seawater as radial symmetric spheres and polarity is established after fertilization. Since the middle of the 19th century, there has been interest in the gametes, zygotes and embryos of fucoid brown algae (Fucus and Silvetia) and a considerable effort has been invested in the study of their gamete recognition and fertilisation, cell wall biosynthesis and polarity acquisition. One asset of these organisms is their broadcast spawning nature. Gamete releases can be experimentally induced resulting in large populations of synchronously developing embryos. In addition, the zygotes are also large (60–100 µm) and therefore easy to (micro-)manipulate.
In fucoids the mechanism of cell polarisation has been well established. Upon fertilization the apolar egg cell develops a cell wall and starts the polarization process. The sperm entry site and subsequently the light direction provide cues for establishing the polarization vector de novo and both determine the direction and sense of the polarisation axis simultaneously.Unfortunately, it is very hard to culture fucoid brown algae. Hence, they are less amenable to genetic and molecular studies. Similarly the development of the cultivable, but isogamous, brown alga Ectocarpus siliculosus as a successful model for genomic research has been accompanied by a shift towards the developmental research questions related to life cycle and fertilisation biology, but away from polarity establishment.
Figure 1. The polarization process in fucoid zygotes. A. Fertilized zygote, showing a sperm induced F-actin patch. B. Zygote polarizing according to the light direction. C. Polarized zygote showing a photopolarized F-actin patch, polarized endomembrane cytoskeleton and polarized MT cytoskeleton. D. Asymmetrically divided embryo with a thallus and rhizoid cell. Adapted from Bogaert et al. 2013.
Since many years, the Phycology Group of Prof. Dr. Olivier De Clerck is studying diverse biological aspects of the brown alga Dictyota for which laboratory culture are more easy to maintain. Fascinated by plant developmental biology I approached Prof. Dr. Olivier De Clerck to inquire for a master dissertation. Coincidentally while studying the mechanism of gamete recognition in Dictyota in the summer of 2007, Prof. Dr. Olivier De Clerck observed that the spherical egg cells deform into elongated rugby ball shaped spheroids after fertilization. Propelled by this finding a master dissertation subject was discussed over some Belgian beers and we teamed up with Prof. Dr. Tom Beeckman of the Root Development group VIB – PSB in Ghent. Early development of the Dictyota embryo quickly turned out to be different from Fucus and the collaboration ultimately matured in a PhD fellowship funded by the FWO (Research Foundation – Flanders).
Figure 2. Female gametophyte of Dictyota dichotoma in the field (credits: H. Verbruggen).
I vividly remember the excitement, observing that the egg cells of Dictyota elongate minutes after addition of the male gametes. Within three minutes a population of spherical egg cells was transformed into a homogenous population of rugby ball shaped cells, with the elongation observable under the stereomicroscope. From that moment, it was clear that the direction of the elongation must have been predetermined; no process could be so fast to determine an elongation direction after fertilisation in those mere seconds. This was later corroborated by TEM sections showing a heterogenous organisation of the organelles suggesting the presence of a preformed axis in the unfertilized eggs. During a research stay in the lab of Dr. Susana Coelho and Dr. Mark Cock at the SBR Roscoff I managed to visualize the autofluorescence of the chloroplasts of eggs being fertilized and provided in vivo evidence that the preformed axis indeed represents the elongation axis.
It is peculiar that this notion has been left unnoticed for so many years given the many efforts of John Lloyd Williams in the late 19th century describing the periodicity of gamete release and the parthenogenetic development of Dictyota dichotoma. Prof. Dr. Dieter G. Müller, best known for his ground breaking work on Ectocarpussiliculosus, his early work also concentrated on Dictyota dichotoma describing the release periodicity of the female gametophytes in laboratory conditions. Only later we heard that Prof. DG Müller observed in the 60s the very same elongation of the eggs, while working for the developmental biologist Prof. Dr. Lionel F. Jaffe. The observation was received with much excitement and Jaffe suspected fundamental differences with the fertilisation response of fucoids, but a technical difficulty in the culturing withheld them from following up this observation. Most probably priorities must have laid in detangling the fertilisation and polarisation of the zygote of Fucus at that time.
Figure 3. Chloroplast autofluorescence showing the preformed axis predicts the elongation axis. Adapted from Bogaert et al. 2017.
The notion to walk in the footsteps of two of the most iconic early developmental biologists working on brown algae was most exciting to us. Quickly we realized that the elongation had implications for the way the cell polarize. While the egg cell has a predetermined elongation axis the direction of the polarisation vector, (aka ‘the sense of the polarization vector’) is not predetermined. In other words, it is not decided which side is going to develop into the leaf-like upper part or the lower root-like part of the alga. This was best illustrated by the fact that while the elongation direction was unresponsive to the direction of the light, which of the two poles will develop into the rhizoid is determined by the direction of the unilateral light. Moreover, toluidine blue O staining – a marker for permanent fixation of the rhizoid pole in fucoids – was only observed hours after fertilization. Therefore determination of the polarisation vector in Dictyota is inherently a two-phased process where initially the direction of the polarisation vector is determined and only later also the sense of the polarization vector.
Figure 4. A scheme comparing zygote polarization in Dictyota,Fucus and Arabidopsis. Timing of the two phases of polarisation are denoted in blue. Red dashed line represents direction of the polarisation vector (without sense). Red line with arrow denotes a polarisation vector with both direction and sense. Adapted from Bogaert et al. 2017.
These two phases are not only completed with a different timing, they also rely on at least two different mechanisms because they depend on two different cues. While the direction is maternally determined, the sense of the polarisation axis is determined by the light direction. Due to the different timing, the two phases even occur in different life stages: the direction in the polarisation vector in the oogonium and the sense is determined hours after fertilization in the diploid sporophyte. In animal systems it is however well known that developmental processes during the first cell divisions are under control of the transcriptome transcribed before egg arrest because de novo transcription (zygotic genome activation) is postponed. This is termed the maternal-to-zygotic transition. The broadcast spawning nature makes it relatively easy to yield large populations of cells, because we did not have to rely on microdissection techniques and FACS techniques. Therefore with some moderate effort we could obtain three biological replicate libraries of mRNA of gametes, zygotes and asymmetrical divided embryos. This allowed us to show that there is a large degree of zygotic genome activation taking place already during the first cell cycle, showing that the next generation is indeed acquiring developmental control over the cell polarisation.
I am thrilled with this publication, because it once more shows that research on oogamous brown algae is not from the past.The availability of an oogamous broadcast spawning brown alga that can be easily cultured and releases its gametes into the external medium opened up the opportunity for developing it into a research model complementary fucoids. This publication however shows that different and complementary insight can be gained from Dictyota early development.
References
Bogaert KA, Arun A, Coelho SM, De Clerck O (2013) Brown algae as model for plant organogenesis. In Methods in Molecular Biology: Plant Organogenesis (De Smet I ed). Humana Press. Springer Protocols, Heidelberg, 959: 97-125.
Bogaert KA, Beeckman T, De Clerck O (2017) Two-step cell polarisation in algal zygotes. Nature Plants3: doi:10.1038/nplants.2016.221.
Organisers: Michael Way, Elizabeth Chen, Margaret Gardel and Jennifer Lippincott-Schwartz Date: 21 – 24 May 2017 Location: Southbridge Hotel & Conference Center, Massachusetts, USA
Speakers:
Anna Akhmanova (Utrecht University, The Netherlands)
Daniel Billadeau (Mayo Clinic, USA)
Anthony Bretscher (Cornell University, USA)
Gaudenz Danuser (University of Texas Southwestern Medical Center, USA)
Cara J. Gottardi (Northwestern University Medical School, USA)
Kathleen J. Green (Northwestern University Medical School, USA)
Erika Holzbaur (University of Pennsylvania, USA)
Johanna Ivaska (University of Turku, Finland)
Tomas Kirchhausen (Harvard Medical School, USA)
Sophie Martin (University of Lausanne, Switzerland)
Mark Peifer (University of North Carolina, USA)
Jenny Russinova (Ghent University, Belgium)
Erik Sahai (The Francis Crick Institute, UK)
Giorgio Scita (IFOM Foundation & University of Milan, Italy)
David Stephens (University of Bristol, UK)
William Trimble (Sick Kids Research Institute, Canada)
Kristen Verhey (University of Michigan, USA)
Gia Voeltz (University of Colorado, USA)
Kenneth M. Yamada (NIH/NIDCR, USA)
The current issue of Development – our Special Issue on Organoids – features a collection of review- and research-based articles focusing on organoids. Here are some of the highlights. Happy reading (and thanks to everyone who contributed)!
Organoids: a Special Issue
In her Editorial, Melissa Little provides an overview of the entire contents of the Special Issue, highlighting some of the important findings and major themes therein.
The hope and the hype of organoid research
In their Spotlight article, Meritxell Huch andJuergen Knoblich, together with Matthias Lutolf and Alfonso Martinez-Arias, discuss the exciting promise of organoid technology, as well as the current limitations and what it will take to overcome them.
Ethical issues in human organoid and gastruloid research
Megan Munsie, Insoo Hyun and Jeremy Sugarman summarize some of the important ethical issues associated with research involving human organoids and other complex self-organized structures. Read their Spotlight article.
The physics of organoids: a biophysical approach to understanding organogenesis
Svend Dahl-Jensen and Anne Grapin-Botton highlight some interesting applications of physics in organoid research, from computational modeling of organoid biology to understanding mechanical aspects of organ development. Read their Spotlight article.
Using brain organoids to understand Zika virus-induced microcephaly
In their Spotlight article, Hongjun Song, Guo-li Ming and colleagues summarise the latest advances in using cerebral organoids to model Zika virus infection and the resulting pathology.
Pluripotent stem cell-derived organoids: using principles of developmental biology to grow human tissues in a dish
This Development at a Glance poster article by Heather McCauley and James Wells summarises how knowledge gained from developmental biology can be used to guide human in vitro organogenesis, and discusses the potential applications of this technology.
Translational applications of adult stem cell-derived organoids
Jarno Drost and Hans Clevers discuss how adult stem cell-derived organoids can be used to model human diseases, create personalized cancer therapies and further efforts in regenerative medicine. Read the Primer article.
Embryoids, organoids and gastruloids: new approaches to understanding embryogenesis
In their Review article, Mijo Simunovic andAli Brivanlou discuss the basic physical and biological principles that underlie the self-organization of embryonic stem cells into organoids, and how this informs human development.
Lung organoids: current uses and future promise
Brigid Hogan and colleagues explore the latest advances in both adult and embryonic stem cell-derived lung organoid culture, and discuss how these systems can be used to understand homeostasis and regeneration. Read their Review.
Dissecting the stem cell niche with organoid models: an engineering-based approach
In their Review, Zev Gartner and co-workers highlight how organoids have been used to model and characterize stem cell-niche interactions and how new engineering approaches enable systematic study of the stem cell niche.
PLUS:
This Special Issue also contains a number of Research Reports, Research Articles and Techniques & Resources Articles – click here for a full listing!
We seek a highly motivated postdoctoral fellow to spear head research investigating the genomic basis of transcriptional specificity of cell signaling during the directed differentiation of human pluripotent stem cells into the digestive and respiratory organoids. The goal is to understand how different transcriptional programs are activated by the same cell signaling pathways, at different times in development and disease. You will join a multidisciplinary team in the Zorn Lab, Division of Developmental Biology at Cincinnati Children’s Hospital Research Foundation.
Qualified applicants will have a PhD with peer review research publications, a demonstrated expertise in genomic analysis, and a keen interest to develop an independent research program in the area of genomics, development and stem cell biology.
Please submit your application to aaron.zorn@cchmc.org with the following information: a cover letter, statement of interest, and CV with contact details for 3 referees.
Closing Date: May 31st, 2017
The Cincinnati Childrens Hospital Medical Center, and the University of Cincinnati are Affirmative Action/Equal Opportunity Employers. Qualified women and minority candidates are especially encouraged to apply.
Department/Location: Wellcome Trust – Medical Research Council Cambridge Stem Cell Institute
Reference: PS11650
Closing date: 31 March 2017
Collaborative PhD Studentship with Astra Zeneca
Elucidating cellular behaviours and potential oncogenicity utilising in vitro organoid culture system Project Description
Department name: Cambridge Stem Cell Institute, University of Cambridge Supervisors: Primary: Dr Joo-Hyeon Lee (University of Cambridge) and Dr Mick Fellows (AstraZeneca)
Assessment of cell transformation (i.e. the acquisition of malignant characteristics in morphology, growth control or function) has proven problematic to model in vitro. The soft agar cell transformation assay has been validated, but only in a limited number of immortalised or mixed population embryonic cells and the specificity of the assay has been questioned. Furthermore, the genetic basis of cell transformation in this model has not been fully delineated (Harvey et al 2015, Creton et al 2012). The use of genetically and phenotypically stable organoids derived from single human stem cells (Schwank et al 2013) should provide a better model for cell proliferation and transformation.
Organoids have the advantage of 3D morphology, being able to be derived from several major organs and can be engineered to demonstrate proliferative effects by introducing a cell type specific ki67 fluorescent protein to tag. Organoid stability is also dependent on several growth factors and inhibitors e.g. EGF, Wnt, BMP inhibitor, TGFb inhibitor. Growth following withdrawal of these factors, which are known pathways implicated in cancer genesis, will be indicative of cell phenotypic and/or transformative changes. It is also proposed that this model could be used to investigate mechanisms of pro-oncogenicity and also the potential off-target effect of genome editing by CRISPR/Cas9, for which there is currently an unmet need for in vitro assays to assess concerns around inappropriate editing in oncogenes.
The project will develop organoids from a variety of tissue sources, including organoids with the ki67 tag, and assessment of proliferative and morphological changes following withdrawal of growth factors after treatment with reference carcinogens and following transfection with specific and promiscuous guide RNA and associated Cas9 protein. Comparative data will be generated from alternative in vitro methodologies to analyse cell transformation e.g. the soft agar assay. Reference carcinogens will be analysed in this assay along with the potential of CRISPR/Cas9 editing to induce cell transformation (which has not been previously assessed). The student will also use next generation sequencing technologies to identify specific genes involved in morphological changes in both cell and organoid cultures. Success will provide new and more relevant model for understanding and assessment of morphological changes and carcinogenesis.
Eligibility and Funding
This studentship covers 4 years’ UK/EU tuition fees (see below for EU eligibility requirements) and a maintenance stipend.
Fully-funded PhD studentships (3 years) for projects in Mathematical/Computational Biology are available at the University of Southampton (Mathematical Sciences). The PhD projects will be supervised by Drs Philip Greulich and Ben MacArthur and will involve mathematical and computational modelling of stem cell dynamics in biological tissues, in particular related to cancer and development. The candidate should have, or should expect, an upper second-class degree or higher, and a genuine interest in biological problems.
An outline of two potential PhD project topics can be found here:
Hanieh Falahati, Lewis–Sigler Institute for Integrative Genomics, Princeton University.
Eric Wieschaus, Howard Hughes Medical Institute, Department of Molecular Biology, Princeton University.
The whole universe is moving toward disorder; this is the second law of thermodynamics in simple terms. Yet, living organisms have found a way to keep themselves organized by spending energy. Cells need this organization in order to provide specialized microenvironments for different cellular functions. Two types of intracellular organizations can be distinguished in biological systems: The first type is membrane-bound organelles such as ER, or lysosome. The composition of these organelles is maintain by spending energy and active transport of molecules across a membrane. The second type of intracellular organization comes from membrane-less organelles such as nucleoli, histone locus bodies, and stress granules that are high-concentration assemblies of different proteins and RNAs. A puzzling question is that without a membrane, how are cells able to form and maintain these organelles.
A historical perspective
The question of how membrane-less organelles form immediately captured my attention the first time I heard about it in an introductory meeting with my doctoral advisor, Prof. Eric Wieschaus. Of course this question has fascinated many scientists since the initial observation of the nucleolus, the quintessential membrane-less organelle, in the 18th century. In 1898, Montgomery performed a very comprehensive study of the nucleoli of different cell types, hand-drawn in 346 figures (a sample is shown in Fig. 1), and concluded that the nucleolus forms when a substance from the cytoplasm enters the nucleus and is deposited there “in the form of masses of varying dimensions, which may be either globular or irregular in shape, according as they are fluid or viscid in consistency”. He further describes that nucleoli form via “coalescence of numerous small portions of nucleolar substance”, consistent with its fluidity (Montgomery 1898). However, this model faded away with the advancement in cell biology and genetics which showed that the nucleolus forms around ribosomal DNA repeats and is the site of active transcription and processing of ribosomal RNA, and ribosomal biogenesis. By the end of the 20th century, nucleolus was commonly thought to form actively, in the process of making ribosomes, until a number of influential works from Hyman and Rosen groups redirected the focus to the liquid nature of the membrane-less organelles (Brangwynne et al. 2009; Li et al. 2012). Based on this liquid property, it was proposed that formation of membrane-less organelles is a spontaneous liquid-liquid phase separation (LLPS), similar to the separation of oil from water.
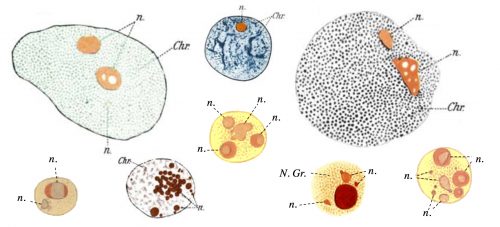
Figure 1. Hand drawing of nuclei with nucleoli (n) of different cell types drawn by Montgomery in 1898. N. Gr.: nuclear granule. Chr.: Chromatin.
By the time I joined the Wieschaus lab at Princeton, two main hypotheses existed for the formation of membrane-less organelles: 1. The LLPS model, in which the components of these organelles assemble spontaneously in a thermodynamically-driven process due to their favorable intermolecular interactions, and form a liquid phase; 2. The active assembly model which suggests that these organelles form as a result of an active process, which happens inside the cells or inside the organelles. Such active processes are reactions carried out by enzymes that couple these reactions to an energy source such as ATP. The evidence for the LLPS model comes from experiments showing that many of the proteins which localize to the membrane-less organelles have the physical properties to phase separate in vitro. However, in most cases the phase separation is only observed at concentrations much higher than cellular levels. Therefore, a pressing question is whether the physical properties of these components would also drive their spontaneous assembly in vivo, or active processes are responsible for their congregation. Accordingly, we decided to develop an in vivo test that would allow us to distinguish between the LLPS and the active mechanism, and apply it to study nucleolus assembly as a model for the formation of membrane-less organelles.
Working out a path – How to test the phase separation model in vivo?
Developing such an in vivo test was not trivial, since compared to in vitro systems, living cells are far more limited in the ways they can be manipulated. But considering the expertise of my advisor, Eric Wieschaus, in manipulating Drosophila embryos, his lab was the best place to do this type of research. It was not the only reason for us to study this question in fly embryos: Development in Drosophila starts without a nucleolus, and the nucleolus forms at a particular developmental stage (Video 1). This allows us to manipulate the system and study its effect on the nucleolus assembly. In addition, nucleolus forms during the 13 synchronous divisions at the beginning of development, therefore at each time-point there are multiple nuclei at the exact same developmental stage, which helps with statistics. Finally, during these 13 division the nuclei share the cytoplasm, which means that all the nuclei have access to the same concentration of most molecules. These features make fly embryos an ideal system for studying nucleolus assembly.
Video 1. Nuclei of D. melanogaster embryos start development without a nucleolus, and form the nucleolus (bright foci) for the first time at nuclear cycle 13.
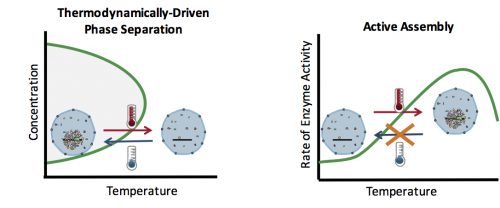
How can you test whether the formation of an organelle in vivo is a thermodynamically driven LLPS, or an active assembly process? In general, LLPS processes are affected by two factors: concentration and temperature (Fig. 2). Increasing the concentration results in an increase in the size of assemblies: The more oil you add to the water, the bigger the oil droplets. However, active processes are also affected similarly with concentration: The higher the concentration of the reactants, the more products (assemblies) are formed. Therefore, changes in the concentration cannot distinguish between an LLPS and an active assembly. On the contrary, temperature often has a differential effect on these two processes: In general, lowering the temperature causes more condensation and enhances phase separation, but slows down active enzymatic reactions. This means that by quantifying the effect of various temperatures on the assembly formation, one can distinguish between an LLPS and an active assembly model.
Figure 2. (Left) Phase separation processes are affected by concentration and temperature. Increasing the concentration and decreasing the temperature results in the formation of larger assemblies. (Right) Active processes are carried out by enzymes which couple them to an energy source. Therefore, the rate of these processes is determined by the rate of enzyme activity, which is reduced at lower temperatures.
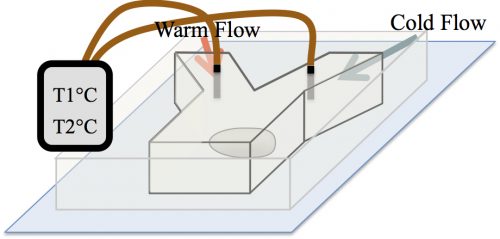
At the time, our strategy to examine the effect of temperature on the formation of assemblies was received with skepticism among the community. The main concern was whether in the temperature range tolerable by biological systems, an effect would be detectable. A reassurance, however, was an exquisite work by Sarah Veatch, who showed that plasma membrane vesicles isolated from living cells exhibit phase transitions between 15 and 25°C (Veatch et al. 2008). Subsequently, we employed a microfluidic device that allowed us to control the temperature of a fly embryo between 6 and 31°C (Fig. 3). We used this microfluidic device to examine the effect of temperature on the first time formation of assemblies, and also to test the reversibility of assembly formation, for six different nucleolar proteins.
Figure 3. A microfluidic device was used to control the temperature of an embryo in a range of 6 to 31°C by flowing in warm or cold water.
The unexpected result of two independent mechanisms
Much to our surprise, the results of our test showed that two independent mechanisms govern the assembly of different subsets of nucleolar proteins: While four of the studied proteins depict properties of LLPS, the two others are recruited actively. At lower temperatures, the phase separating proteins assemble at an earlier developmental time, and dissolve upon heating (Video 2). The latter is consistent with the reversibility of thermodynamic processes. On the contrary, the formation of active proteins is inhibited at low temperatures, and is irreversible, an exclusive property for active processes.
Video 2. The nuclei of intact fly embryos are subjected to temperature changes in the surrounding fluid. As the temperature is shifted from low to high, the phase separated proteins dissolve, as can be seen in the disappearance of the bright spots.
rDNA: Come together, right now, over me
Our unexpected result that two independent mechanisms govern nucleolus assembly in vivo raised a new question: How are these two different mechanisms coordinated to form a single organelle? In a previous paper, we had shown that transcription of rDNA dictates the spatiotemporal precision in the formation of assemblies by the phase separating proteins (Falahati et al. 2016). Interestingly, we were also able to show that rDNA is necessary for the recruitment of the two actively assembling proteins. This suggests that rDNA can function as a coordinator between the two independent mechanism. In addition, it highlights the fact that in vivo, even the assembly of phase separating proteins is regulated by active processes such as transcription.
We are very enthusiastic about the publication of this paper as it is a significant advancement in the field both from a methodological and mechanistic standpoint. We took an interdisciplinary approach and developed an invaluable tool for closing the gap between the current knowledge of the in vitro self-assembly and the formation of membrane-less organelles in vivo at its full complexity. From a mechanistic perspective, the large number of proteins studied here allowed us to unravel the unappreciated complexity in the formation of intracellular assemblies, as our results show the presence of at least two independent mechanisms for the recruitment of nucleolar proteins. We are confident that the method and the results introduced in this paper will set a framework to better understand how normal or pathological intracellular assemblies form.
Our latest monthly trawl for developmental biology (and other cool) preprints. See June’s introductory post for background, and let us know if we missed anything
Another month, another bumper crop of preprints covering everything from calcium waves in the fly to the first mutant ant line, plant superhero genes to rat embryonic stem cells. Plus, as usual, a whole bunch of tools and resources (including a 3D printed micropipete!), and right at the end some enigmatic bacterial structures.
The preprints were hosted predominantly on bioRxiv, though we also found a handful on arXiv and PeerJ.
Multiple roles of PIWIL1 in mouse neocorticogenesis. Barbara Viljetic, Liyang Diao, Jixia Liu, Zeljka Krsnik, Sagara H. R. Wijeratne, Ryan Kristopovich, Marina Dutre-Clarke, Matthew L. Kraushar, Jimin Song, Jinchuan Xing, Kevin Chen, Mladen-Roko Rasin
Laser ablation tomography image of a nodal root from a maize plant, from Bucksch, et al.
Morphological plant modeling: Unleashing geometric and topological potential within the plant sciences. Alexander Bucksch, Acheampong Atta-Boateng, Akomian Fortune Azihou, Mathilde Balduzzi, Dorjsuren Battogtokh, Aly Baumgartner, Brad Binder, Siobhan Braybrook, Cynthia Chang, Viktoriya Coneva, Thomas DeWitt, Alexander Fletcher, Malia Gehan, Diego Hernan Diaz Martinez, Lilan Hong, Anjali Iyer-Pascuzzi, Laura Klein, Samuel Leiboff, Mao Li, Jonathan Lynch, Alexis Maizel, Julin Maloof, RJ Cody Markelz, Ciera Martinez, Laura Miller, Washington Mio, Wojtek Palubicki, Hendrik Poorter, Christophe Pradal, Charles Price, Eetu Puttonen, John Reese, Ruben Rellan-Alvarez, Edgar Spalding, Erin Sparks, Chris Topp, Joseph Williams, Daniel Chitwood
Tex19.1 Restricts LINE-1 Mobilisation in Mouse Embryonic Stem Cells. Marie MacLennan, Marta Garcia-Canadas, Judith Reichmann, Elena Khazina, Carmen Salvador-Palomeque, Abigail Mann, Paula Peressini, Laura Sanchez, Christopher J Playfoot, David Read, Chao-Chun Hung, Ragnhild Eskeland, Richard R Meehan, Oliver Weichenrieder, Jose Luis Garcia- Perez, Ian R Adams
Sagittal sections of mandibular molars from Feng, et al.
Functional Annotation of Chemical Libraries across Diverse Biological Processes. Jeff S Piotrowski, Sheena C Li, Raamesh Deshpande, Scott W Simpkins, Justin Nelson, Yoko Yashiroda, Jacqueline Barber, Hamid Safizadeh, Erin Wilson, Hiroki Okada, Abraham Gebre, Karen Kubo, Nikko Torres, Marissa LeBlanc, Kerry Andrusiak, Reika Okamoto, Mami Yoshimura, Eva DeRango-Adem, Jolanda van Leeuwen, Katsuhiko Shirahige, Anastasia Baryshnikova, Grant W Brown, Hiroyuki Hirano, Michael Costanzo, Brenda Andrews, Yoshikazu Ohya, Hiroyuki Osada, Minoru Yoshida, Chad L Myers, Charles Boone
TOWARDS COORDINATED INTERNATIONAL SUPPORT OF CORE DATA RESOURCES FOR THE LIFE SCIENCES. Warwick Anderson, Rolf Apweiler, Alex Bateman, Guntram A Bauer, Helen Berman, Judith A Blake, Niklas Blomberg, Stephen K Burley, Guy Cochrane, Valentina Di Francesco, Tim Donohue, Christine Durinx, Alfred Game, Eric Green, Takashi Gojobori, Peter Goodhand, Ada Hamosh, Henning Hermjakob, Minoru Kanehisa, Robert Kiley, Johanna McEntyre, Rowan McKibbin, Satoru Miyano, Barbara Pauly, Norbert Perrimon, Mark A Ragan, Geoffrey Richards, Yik-Ying Teo, Monte Westerfield, Eric Westhof, Paul F Lasko
Uncharacterized bacterial structures revealed by electron cryotomography. Megan J Dobro, Catherine M Oikonomou, Aidan Piper, John Cohen, Kylie Guo, Taylor Jensen, Jahan Tadayon, Joseph Donermeyer, Yeram Park, Benjamin A Solis, Andreas Kjaer, Andrew I Jewett, Alasdair W McDowall, Songye Chen, Yi-Wei Chang, Jian Shi, Poorna Subramanian, Cristina V Iancu, Zhuo Li, Ariane Briegel, Elitza I Tocheva, Martin Pilhofer, Grant J Jensen
A postdoctoral research fellow position is available in the laboratory of Gregor Andelfinger at the CHU Sainte Justine Research Center, Montréal, Québec, Canada. Our work focuses on the genetic origins of cardiovascular diseases in the young and includes both genomics and functional approaches. In this project, the successful candidate will apply genomic and molecular biology approaches to develop disease models. Postdoctoral fellows will have a unique opportunity to do interdisciplinary research at all interfaces of translational medicine and publish their results in high impact journals.
The candidate will work in collaboration with all lab members and is expected to independently perform human and mouse genetic studies, as well as in vitro molecular analyses. Experience with a wide variety of techniques (developmental studies, histology, confocal microscopy, cell culture assays, RNAseq) is desired. Experience with genome editing techniques (CRISPR) and/or and bioinformatics analysis is an asset.
The successful candidate must have a recent Ph.D. degree in life sciences, molecular cell biology, or a related field. We are seeking highly motivated and well published researchers with excellent oral, written, and interpersonal communication skills.
Montreal is a vibrant bilingual city with a high quality of life. With a greater metropolitan area of 3.5 M, Montreal offers all scientific and cultural benefits of a large North American agglomeration with a European flair and beautiful surroundings.
The Andrew Yoo lab at WUSTL is looking for a lab manager/technician. The Yoo lab uses microRNA-mediated direct reprogramming of fibroblasts to neurons to study aging and neurodegenerative diseases.
![[Scheme - Fucus cell polarisation]](https://thenode.biologists.com/wp-content/uploads/2017/03/figure1-500x131.png)
![[habitus Dictyota]](https://thenode.biologists.com/wp-content/uploads/2017/03/img_0190-500x375.jpg)
![[chlorooplast distribution - elongation]](https://thenode.biologists.com/wp-content/uploads/2017/03/figure2-500x186.png)
![[scheme Dictyota, Fucus, Arabidopsis]](https://thenode.biologists.com/wp-content/uploads/2017/03/fig3-500x511.png)
 (3 votes)
(3 votes) (No Ratings Yet)
(No Ratings Yet) This
This  Jarno Drost and
Jarno Drost and  n their
n their  Brigid Hogan and colleagues explore the latest advances in both adult and embryonic stem cell-derived lung organoid culture, and discuss how these systems can be used to understand homeostasis and regeneration. Read their
Brigid Hogan and colleagues explore the latest advances in both adult and embryonic stem cell-derived lung organoid culture, and discuss how these systems can be used to understand homeostasis and regeneration. Read their  In their
In their