Graduate Peer Group: How to Lead When You Are Not in Charge
Posted by Tomer Stern, on 29 October 2015
Itamar Harel¹* and Tomer Stern²*
¹ Department of Biological Regulation, Weizmann Institute of Science, Rehovot, 76100, Israel
Present address: Department of Genetics, Stanford University, Stanford, CA 94305, USA
² Department of Molecular Genetics, Weizmann Institute of Science, Rehovot, 76100, Israel
* Correspondence: itamarharel1980@gmail.com (IH), tomer.stern@weizmann.ac.il (TS)
–
Starting early: How can we make more out of graduate school?
In the long and winding road of a scientific career, graduate school is a benchmark and a springboard for days to come and how high we “jump” clearly depends on a variety of factors. Yet, it seems that some of them are entirely unrelated to our experimental results, nor taught in the many years we spend on the bench, in the lecture hall, or in front of our computers. How can we to improve the guidance of our mentors to better fit our personal needs? How should we initiate and maintain a successful collaboration? What can we do to improve the presentation of our work in meetings or publications? Is it possible to be productive graduate students without letting it take over our lives? How should we find a new lab for our rapidly approaching postdoc? According to the physicist and author Evelyn Fox Keller (Keller, 1996), there is an invisible equation stating that scientific = logical and therefore emotional = non-scientific. Does this leave less room for ‘soft skills’ (See box A) in science? Much is discussed about ‘The Researcher’s Toolkit’ as the sum of methods and techniques practiced by a scientist, although it appears that far more is needed in order to become a successful and happier one.
On February last year, we attended a scientific meeting that was held on the peaceful shores of the Dead Sea. After a long day of lectures has ended, we enjoyed an informal lecture by Prof. Uri Alon from the Weizmann Institute of Science (WIS), who presented several topics related to nurturing scientists (Alon, 2009). It was then that we first heard about the ‘Young PI Forum’ (Milo and Schuldiner, 2009) . The idea behind this group, explained Prof. Alon, is to nurture young principal investigators (PI’s) and help them face the various challenges of the first steps in their independent careers. “Why not start earlier?” we thought; but are we putting the cart before the horse? Clearly, dilemmas concerning topics such as mentoring or hiring research students are not relevant to us, yet. Thinking about this possible adventure, several questions occupied our thoughts, such as are there any advantages for starting early? what are the unique requirements of graduate students, when compared to undergraduate students, postdocs or young PIs (Milo and Schuldiner, 2009), and how will these in turn reflect on such a peer group?
Every step in our scientific training has its own features. During graduate studies, we begin to shape our identity as independent scientists and our voice is starting to be heard. We gradually learn how to lead projects, initiate collaborations, read and criticize the scientific works of others and, finally, decide if and where to train as postdocs. On the other hand, in many aspects we are not yet fully ‘ in charge’. We are still significantly dependent on the guidance, approval and support of our supervisors and we have many questions about the academic world and our place in it. Identifying these unique characteristics has enabled us to start designing our own peer group, in terms of goals, needs, format and especially the case studies for our discussions. Looking back on more than a year of activity, we can say that the ‘WIS Graduate Peer Group’ has helped us find inspiring solutions, a fresh point of view and great support in the traditional, yet slightly forgotten concept of “the power of the group”.
–
More than just a group of friends: A DIY recipe
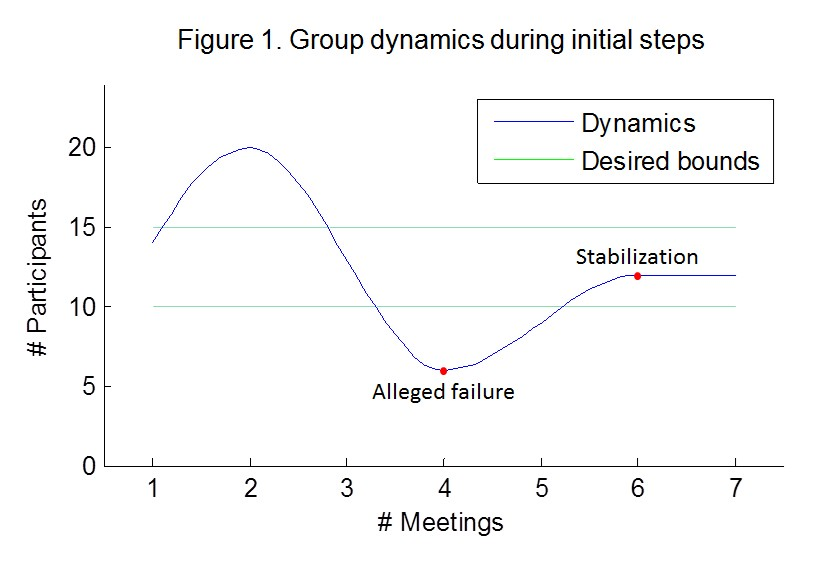
Our main objective was to establish a group with an intimate and interactive atmosphere, enabling a dialogue over the various challenges and dilemmas we encounter during graduate school. As this is uncharted terrain, initiating this activity in graduate school may pose a variety of challenges; the main one is of course creating and maintaining a committed and steady discussion group. As this type of activity is unfamiliar to graduate students, therefore, when we started ‘recruiting’ students around campus, reactions ranged from suspicion, skepticism and fear of sharing, to enthusiasm and excitement. Prepare yourselves for the possibility that in the beginning, there might be meetings attended by no more than a handful of people. Knowing this in advance will help to alleviate the frustration from the alleged failure and will keep you going until the group starts to stabilize (Fig. 1). When such a group performs its first steps, it is perfectly natural that some participants will not relate to this type of activity and will clear their seats for new members. Eventually, the group will converge to its final size and composition. Additional challenges were acquiring the consent of scientists to attend as guest speakers, and getting faculty financial support for funding of management courses, refreshments, etc. Notably, we enjoyed the friendly advice and opinion of experienced researches at our institute, especially in our first steps. Therefore, we recommend looking for researchers from your faculty with which you can share some of your questions and challenges.
After extensive trial-and-error during the last year, we wish to present a few ground rules we found to be useful. Several features of the group, such as the recommended number of participants, format of meetings (e.g. discussing dilemmas vs. teaching a skill) and the role of the coordinators, were inspired by known models of discussion groups (Klein and Lande-Diner, 2011; Milo and Schuldiner, 2009). Therefore, we will focus mainly on the lessons we have learned from our own experience and the adjustments we made accordingly.
Confidentiality: Although it is not unique to our group, this is the most important guideline we would like to offer. In this forum, members share their private thoughts and intimate experiences; therefore, maintaining absolute discretion among members is critical. It is necessary to stress this point and to create an atmosphere that will promote gradual building of trust.
Participants: The meetings require intimacy and therefore it is preferred that the number of participants will range between 10-15 students. Consequently, inviting every graduate student in the faculty is not advisable, as more applicable for the young PI forum (Milo and Schuldiner, 2009). In order to obtain a wide spectrum of opinions, try to diversify the group in terms of stages of graduate studies, gender etc. Furthermore, since personal dilemmas raised in discussion often involve lab mates or supervisors, we would advise to avoid the participation of multiple members from the same lab.
Guest lecturers: Every third meeting we invited a guest speaker, who can be a group leader, an expert in a certain field, or any other person of interest. We found that an open and informal discussion between graduate students and PIs was extremely fruitful, enabling us to discuss issues we typically wouldn’t feel comfortable raising in front of our own advisors. These meeting were dedicated to a wide variety of topics, ranging from professional skills, such as “How to write a good scientific paper” with Prof. Eran Segal, to nurturing ‘soft skills’ by getting acquainted with the “The impostor syndrome” with Dr. Maya Schuldiner (see box B). In other meetings, we had an opportunity to discuss “The passion in science making” with the Nobel laureate Prof. Ada Yonath, and learn about how scientific paradigms rise and fall (Kuhn, 1996) in a meeting with Prof. Yadin Dudai.
Online forum: A private online forum enables members to continue the dialogue after the session, keep records and upload materials relevant to recently discussed topics. In addition, the forum informs members about upcoming meetings.
–
From a dilemma to a solution – a few case studies
During the passing year, we discussed fascinating dilemmas and topics, which we would like to share as case studies.
‘Managing up’: An honest confession: as graduate students, we like to complain about our advisors, and quite often. Our mentors are either rarely around or they burden us with frequent meetings, scarcely guide us or limit our independence, overly strict and organized or unpredictable and chaotic – you name it. How hard can it be for a world leader in cancer research to handle ’straightforward‘ tasks such as mentoring, nurturing us to become independent scientists and, on top of these, have perfect social skills? Is that really too much to ask?! We often tend to assume that to be an advisor is to miraculously acquire all the skills of advising. But when we think of it, our supervisors are just like us, only several years ahead. Discussing this topic in our peer group has yielded interesting practical advices, along with valuable support. Based on a few case studies, we would like to offer some suggestions on how to make the most of the challenging “student- advisor” relationship.
Our discussions related to personal interactions and communication, constructive feedback and criticism, effective meetings with the supervisor etc. We found that in order to relieve ‘communication tight spots’, we first needed to define what we wanted. In other words, instead of expecting our advisors to know what we lack, we should merely ask for what we need. This is sometimes easier said than done, but it’s worth the try. For instance, while asking for comments on a written paper, one student described criticism by the supervisor as vague remarks muttered on the way to the coffee machine, whereas another student mentioned that the returned manuscript seemed like ‘pages over pages of a tick-tac-toe tournament’ with red ink. Can this be improved? For example, if we wish for specific feedback on the ‘Results’ section, sending the entire text and asking for an opinion might be counterproductive. However, it would be useful to meet beforehand and discuss the general direction before submitting the complete work to our advisor. This brings us to our next topic, properly organized meetings.
Our mentors are busy; no surprise there, but this only means that we ought to be more persistent. Regular meetings with our supervisors are pivotal to the success of our studies for obvious reasons, such as getting good advice, better focus and avoiding unnecessary mistakes. By ‘meeting’ we mean one-on-one sessions; group meetings or random discussions are no substitute. Furthermore, the more familiar our supervisors are with our work, the more likely they are to be interested in it, to further read, think about and present it, as well as to learn how we think and work. Therefore, describe to yourself what you want to achieve from an upcoming meeting and come prepared. We found that a simple and well-defined target may work better than an overwhelming flood of topics and data.
How can we lead when we are not in charge? Or in other words, how can we ‘manage up’? (Kearns and Gardiner, 2011). To most of us, there comes a point in our graduate studies when we realize that the responsibility for our research, lies mostly on our shoulders, along with the enjoyment of practicing it. Our supervisors possess a range of good and poor qualities. We have to benefit from what they have to offer; and when we are not content, learn how to guide them to better fit our needs, or find alternatives elsewhere (such as consulting mentors that are not our PIs). The qualifications and traits that will help us become world leaders in cancer research are not necessarily those that will make us good mentors. However, exercising these often-neglected aspects during graduate school, for instance by guiding other research students, may provide an early start for acquiring and improving mentoring skills.
‘Searching for a postdoc position’: Choosing a lab for postdoctoral research is tough, really tough, and probably one of the most significant and difficult science-related decision we have to make. As graduate students, we have spent the last several years gaining expertise in our field. We know by now what the leading labs and central paradigms are; no doubt, valuable knowledge. But what if we change direction? There are so many questions to answer in a very short time. Which topic should I choose, and how do I do it when there are so many I hardly know anything about? How do I compose a list of potential labs and make my visits as effective as possible? This decision is very personal, as it is based on one’s personality and scientific interests. Nevertheless, discussing this topic in our meetings has proved highly useful. First, realizing that we were not alone and that it was just as hard for everybody else was comforting. Furthermore, it has enabled us to identify common ground rules that can help us all on this route. These helpful tips have been drawn from the personal experience of members that were searching for a postdoc position at that time, as well as from the experience of visiting young PIs. In the following section, we will share some of our collective dilemmas and the insights we have gained by discussing this complicated decision.
Choosing the topic for a postdoc is a rare opportunity for scientists to completely change their field of research. We found that toward the end of our graduate studies, it was useful to step out of our niches and to expose ourselves to a wide range of topics unrelated to our research. For instance, although most of us have by then accumulated the required credit points, taking some courses (even as a free auditor) was a great way to start. Reading articles and going to lectures and meetings outside our field is also recommended. Once we had several directions in mind, we consulted with researches in relevant fields (in our university or in neighboring ones) regarding potential postdoc labs. We prepared our own list of candidate labs, as well as asked for advice and recommendations regarding others we might have missed.
There are many other personal aspects to consider besides the research topic. For instance, whereas some of us preferred a big and established lab, others looked for a smaller lab likely run by a younger PI (both options have great advantages). Other considerations, such as where you (or your spouse) would want to live, and even seemingly trivial factors such as the climate, may be taken into account. We tried to inquire as much as possible about the labs before we applied; former or current postdocs from the lab or the department may provide valuable information. When we wrote our letters of application, we made sure they reflected the fact that we had carefully studied the lab. If you don’t get an answer, don’t take it personally; we learned that persistence could be rewarding, as sending an additional mail or have a mutual acquaintance introduce you can make all the difference.
The ‘tour’ itself can be quite stressful. From our experience, visiting more than four or five labs in a single trip might be a bit overwhelming; if possible, it is better to make two trips instead. Keeping that in mind, in order to facilitate this costly endeavor, try to look for funding sources, such as traveling grants, and ask for assistance from the labs you are visiting. In preparing your talk, don’t forget that your crowd might not be familiar with your scientific discipline. Therefore, try to present your work as a story that would be interesting and accessible to a broad audience and try to rehearse your talk in front of a mixed crowd as well.
The chemistry with your future PI is perhaps the most important aspect to consider – don’t compromise! This was emphasized repeatedly by postdocs and young PIs that were guest speakers in our group. Furthermore, there are important practical matters that you should discuss with your future mentor, such as financial support, grants, your family etc. Make sure to ask the lab-members questions about issues that are important to you, preferably outside the lab or over lunch. What are the PI’s strengths and weaknesses? How available is he/she? Are there collaborations among the group members? What are the expected working hours? As long as you ask politely, it is completely OK. Listen carefully to criticism you hear and don’t ignore it, since it is likely to be true. Finally, don’t forget that although you will try to impress people in your visit, they should impress you just as much.
Looking for a postdoc position is hard, and doing it alone and without guidance is even harder. Although this might feel like preparing for a military operation, don’t let these challenges spoil the fun and don’t forget to enjoy it. It is essential to keep in mind that the postdoc period is a great opportunity to grow, scientifically as well as personally. If we don’t enjoy the journey by fulfilling our scientific curiosity, expressing our creativity and originality, then what are we doing it for?
–
Conclusion and next steps
Graduate school is a wonderful period, during which we ask questions and obtain the tools needed to reveal the secrets of nature. This period is also well known for its diverse range of challenges, stresses and frustrations. It was fascinating to realize how common these are among our colleagues. As scientists, studying the literature or discussing our results with our peers seems practical and almost obvious. We think that a similar approach can and should be applied to ‘non-scientific’ aspects of our everyday life as graduate students. Along the same line, this concept may very well be relevant for earlier academic stages, each with its own characteristics. One of our main goals in forming the ‘Graduate Peer-Group’ was to address these somewhat neglected aspect of our education as young scientists; Such as getting familiar with relevant psychological theories (e.g. ‘the imposter syndrome’), techniques for personal interactions (e.g. soft skills, ‘managing up’), practical information and sharing of experience (e.g. finding the right place for postdoc training), combined with the support of our peers and friends.
The described set-up was previously shown to be successful for young PIs. With the proposed modifications, we show its great potential for graduate students. We hope our experience will serve as an example, demonstrating the central role a peer group can play during graduate school and assist in initiating it. Excitingly, by word of mouth, the Graduate Peer-Group idea has spread. Currently, similar groups are being established in several places, including the Sheba medical center and the Tel-Aviv University. We believe that such programs should be encouraged and supported, possibly as an integral part of the syllabus. In case of high demand, multiple peer groups can operate in parallel at the same institution, ideally with formal assistance, such as management courses for the coordinators. Initiating such programs will provide a source of knowledge and support, and prepare us for a future independent career, when we will guide our own students.
Our experience in the peer group also demonstrated the importance of soft skills. Classic “hard science” disciplines, such as medicine, science and engineering, are often bound to traditional professional behavior models. In some aspects, these models might find themselves at odds with the diverse interpersonal interactions required in today’s workplaces. We think that developing soft skills through educational and professional enrichment activities, such as the peer-group also puts us responsible for confronting our current challenges as graduate students, as well as to our own career development. Therefore, addressing and improving these skills in graduate school should not be considered as ‘too early’ – but rather ‘just on time’. Finally, mastering these ‘non-scientific’ aspects of our everyday life means that, by relieving unnecessary frustrations, more time and energy would be devoted to creative science – making us happier as well as better scientists.
–
Box A – Soft skills:
This sociological term, also known as “people skills”, refers to a set of personal traits and social abilities, allowing someone to effectively interact with other people. Soft skills relate to a person’s “EQ” (Emotional Intelligence Quotient), and considered essential to both personal and professional success by complementing hard skills which is more associated with a person’s IQ. In a job-related environment as well as in the scientific world, soft skills are valuable (and can be nurtured and improved) in a variety of aspects (Heckman and Kautz, 2012), such as scientific presentations, conflict resolution and collaborations, in building a successful research team etc’.
–
Box B – The imposter syndrome:
The impostor syndrome (Clance and Imes, 1978; Felder, 1988; Laursen, 2008) is a psychological phenomenon in which people, despite their evident competence and recognition by their peers, are incapable of internalizing their own success and accomplishments. This syndrome is not limited to scientists or the scientific community, and those with it feel as frauds which do not deserve the success they have achieved. Interestingly, any proof of success is regarded as sheer luck, timing, or as a result of misleading others into thinking they are more intelligent and competent than they believe themselves to be. The term was coined by clinical psychologists Pauline Clance and Suzanne Imes in 1978, and although not an official psychological disorder, it has been discussed in of many books and articles by psychologists and educators.
–
–
REFERENCES
Alon, U. (2009). How to choose a good scientific problem. Molecular cell 35, 726-728.
Felder, R. (1988). Impostors everywhere. Chemical Engineering Education 22, 168-169.
Heckman, J.J., and Kautz, T. (2012). Hard evidence on soft skills. Labour Economics.
Kuhn, T.S. (1996). The structure of scientific revolutions (University of Chicago press).
Laursen, L. (2008). No, You’re Not an Impostor. Science Careers.
 (2 votes)
(2 votes) (5 votes)
(5 votes) My name is Helena, and for the next few months I’ll be helping to run the Node, while Cat concentrates on other projects to make the Node even better! I am Spanish, but I moved to the UK five years ago to study Biochemistry. I am currently doing my PhD in Alfonso Martinez Arias’ lab at the Department of Genetics in Cambridge: I use mouse embryonic stem cells to study patterning in the embryo, and to try and elucidate how different signalling molecules coordinate this patterning during gastrulation. My PhD programme (the
My name is Helena, and for the next few months I’ll be helping to run the Node, while Cat concentrates on other projects to make the Node even better! I am Spanish, but I moved to the UK five years ago to study Biochemistry. I am currently doing my PhD in Alfonso Martinez Arias’ lab at the Department of Genetics in Cambridge: I use mouse embryonic stem cells to study patterning in the embryo, and to try and elucidate how different signalling molecules coordinate this patterning during gastrulation. My PhD programme (the



 This post is part of a series on a day in the life of developmental biology labs working on different model organisms. You can read the introduction to the series
This post is part of a series on a day in the life of developmental biology labs working on different model organisms. You can read the introduction to the series