In Development this week (Vol. 141, Issue 13)
Posted by Seema Grewal, on 24 June 2014
Here are the highlights from the current issue of Development:
Eye’s got rhythm
 In zebrafish, the circadian clock, which is the internal timekeeper that coordinates multiple cellular, physiological and behavioural processes with the external rhythmic environment, begins cycling very early in development. However, the functional relevance for embryonic and larval development of these early circadian oscillations is unclear. Here (p. 2644), Ricardo Laranjeiro and David Whitmore find that a number of important developmental regulators show rhythmic expression in a manner consistent with circadian regulation. In particular, they uncover strong circadian expression of the neural transcription factor Neurod, whose levels oscillate specifically in the photoreceptor layer of the retina. They further show that a number of other key regulators of retinal photoreceptor differentiation oscillate, but only after differentiation itself is essentially complete, implying that this rhythmic expression is unrelated to the known functions of these factors in cell fate specification. Instead, the authors propose that certain components of the phototransduction pathway – which also show cyclic expression – may be controlled by these developmental transcriptional regulators, suggesting an intriguing interplay between the circadian clock and key regulators of retinal differentiation and function.
In zebrafish, the circadian clock, which is the internal timekeeper that coordinates multiple cellular, physiological and behavioural processes with the external rhythmic environment, begins cycling very early in development. However, the functional relevance for embryonic and larval development of these early circadian oscillations is unclear. Here (p. 2644), Ricardo Laranjeiro and David Whitmore find that a number of important developmental regulators show rhythmic expression in a manner consistent with circadian regulation. In particular, they uncover strong circadian expression of the neural transcription factor Neurod, whose levels oscillate specifically in the photoreceptor layer of the retina. They further show that a number of other key regulators of retinal photoreceptor differentiation oscillate, but only after differentiation itself is essentially complete, implying that this rhythmic expression is unrelated to the known functions of these factors in cell fate specification. Instead, the authors propose that certain components of the phototransduction pathway – which also show cyclic expression – may be controlled by these developmental transcriptional regulators, suggesting an intriguing interplay between the circadian clock and key regulators of retinal differentiation and function.Keeping dendrites in check
 A key question in developmental biology is how different tissues maintain proportional growth during development. A striking example of this is the tiling of sensory dendrites across the body wall of theDrosophila larva: during early larval life, the neuronal dendrites extend to cover the entire body wall, without overlapping. As the larva grows further, tiling is maintained – meaning that the dendrites and the overlying epithelium grow proportionally (dendrite-substrate coupling). On p. 2657 Jay Parrish and colleagues investigate the mechanistic basis of this coupling, finding that the microRNA bantam (which they previously showed to be required in the epithelial body wall for proper scaling) regulates endoreplication of these epithelial cells. Inhibiting endoreplication by multiple means disrupts dendrite-substrate coupling such that dendrites overgrow. Moreover, they show that integrin expression in the epithelium is controlled by bantam and other regulators of endoreplication, and is in turn important for appropriate dendrite-epithelial contacts to be made and maintained for proportional growth. Thus, by coordinating cell growth (endoreplication) with epithelial cell-dendrite adhesion, coupled tissue growth can efficiently be achieved.
A key question in developmental biology is how different tissues maintain proportional growth during development. A striking example of this is the tiling of sensory dendrites across the body wall of theDrosophila larva: during early larval life, the neuronal dendrites extend to cover the entire body wall, without overlapping. As the larva grows further, tiling is maintained – meaning that the dendrites and the overlying epithelium grow proportionally (dendrite-substrate coupling). On p. 2657 Jay Parrish and colleagues investigate the mechanistic basis of this coupling, finding that the microRNA bantam (which they previously showed to be required in the epithelial body wall for proper scaling) regulates endoreplication of these epithelial cells. Inhibiting endoreplication by multiple means disrupts dendrite-substrate coupling such that dendrites overgrow. Moreover, they show that integrin expression in the epithelium is controlled by bantam and other regulators of endoreplication, and is in turn important for appropriate dendrite-epithelial contacts to be made and maintained for proportional growth. Thus, by coordinating cell growth (endoreplication) with epithelial cell-dendrite adhesion, coupled tissue growth can efficiently be achieved.
Sampling the SAM
 The shoot apical meristem (SAM) is the growing tip of the plant stem, from which a population of pluripotent stem cells generates all above-ground organs. The SAM is organised both in a central-to-peripheral manner, with the central zone containing the stem cells while their progeny differentiate in the peripheral zone, and in outer-to-inner layers that generate different cell types. These different zones and layers of the SAM are presumably defined and regulated by distinct (if overlapping) gene regulatory networks, and G. Venugopala Reddy and co-workers (p. 2735) set out to define the gene expression landscape of theArabidopsis SAM. They isolate multiple different cell populations from the SAM and perform a detailed transcriptomic analysis to compare the gene expression profiles of the various populations. From these data, the authors are able to identify specific characteristics of particular cell populations, which might provide insight into functional differences between different regions of the SAM. Importantly, the datasets provide a valuable resource for the community and should stimulate further research to better understand the complexity of cell states within SAMs.
The shoot apical meristem (SAM) is the growing tip of the plant stem, from which a population of pluripotent stem cells generates all above-ground organs. The SAM is organised both in a central-to-peripheral manner, with the central zone containing the stem cells while their progeny differentiate in the peripheral zone, and in outer-to-inner layers that generate different cell types. These different zones and layers of the SAM are presumably defined and regulated by distinct (if overlapping) gene regulatory networks, and G. Venugopala Reddy and co-workers (p. 2735) set out to define the gene expression landscape of theArabidopsis SAM. They isolate multiple different cell populations from the SAM and perform a detailed transcriptomic analysis to compare the gene expression profiles of the various populations. From these data, the authors are able to identify specific characteristics of particular cell populations, which might provide insight into functional differences between different regions of the SAM. Importantly, the datasets provide a valuable resource for the community and should stimulate further research to better understand the complexity of cell states within SAMs.Characterising developmental ‘super-repressors’
 DNA and histone methylation patterns correlate with – and define – transcriptional activity of the genome. In particular, DNA hypomethylation is associated with active chromatin and generally thought to be permissive for gene transcription. However, this rule is not globally applicable, and Shinichi Morishita, Hiroyuki Takeda and colleagues (p. 2568) now identify a particular class of hypomethylated domains (HMDs) in pluripotent cells of the medaka fish that are associated with strong gene repression. These HMDs are characterised by their large size and strong H3K27me3 deposition, and are referred to as large K27HMDs. Notably, they are most commonly found surrounding promoters of key developmental transcription factors that are under strong transcriptional repression. These HMDs shorten in mature cells, where the genes are expressed, due to DNA hypermethylation in these regions. Importantly, the authors find that a significant proportion of large K27HMDs are conserved between medaka and human stem cells. Together, these data define a genomic feature – the large K27HMD – that may be responsible for ensuring that key developmental transcription factors are kept strongly repressed in pluripotent cells.
DNA and histone methylation patterns correlate with – and define – transcriptional activity of the genome. In particular, DNA hypomethylation is associated with active chromatin and generally thought to be permissive for gene transcription. However, this rule is not globally applicable, and Shinichi Morishita, Hiroyuki Takeda and colleagues (p. 2568) now identify a particular class of hypomethylated domains (HMDs) in pluripotent cells of the medaka fish that are associated with strong gene repression. These HMDs are characterised by their large size and strong H3K27me3 deposition, and are referred to as large K27HMDs. Notably, they are most commonly found surrounding promoters of key developmental transcription factors that are under strong transcriptional repression. These HMDs shorten in mature cells, where the genes are expressed, due to DNA hypermethylation in these regions. Importantly, the authors find that a significant proportion of large K27HMDs are conserved between medaka and human stem cells. Together, these data define a genomic feature – the large K27HMD – that may be responsible for ensuring that key developmental transcription factors are kept strongly repressed in pluripotent cells.
Plus…
Amyloid precursor protein and neural development
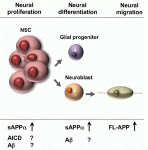 Interest in the amyloid precursor protein (APP) has increased in recent years due to its involvement in Alzheimer’s disease. Understanding the basic biology of APP and its physiological role during development thus will provide a better comprehension of Alzheimer’s disease. Here, Nicolas and Hassan present an overview of some of the key studies performed in various model organisms that have revealed roles for APP at different stages of neuronal development. See the Primer on p. 2543
Interest in the amyloid precursor protein (APP) has increased in recent years due to its involvement in Alzheimer’s disease. Understanding the basic biology of APP and its physiological role during development thus will provide a better comprehension of Alzheimer’s disease. Here, Nicolas and Hassan present an overview of some of the key studies performed in various model organisms that have revealed roles for APP at different stages of neuronal development. See the Primer on p. 2543
The roles and regulation of multicellular rosette structures during morphogenesis
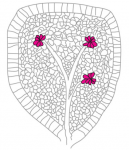 Multicellular rosettes have recently been appreciated as important cellular intermediates that are observed during the formation of diverse organ systems. Here, Nechiporuk and colleagues review recent studies of the genetic regulation and cellular transitions involved in rosette formation. They discuss and compare specific models for rosette formation and highlight outstanding questions in the field. See the Review on p. 2549
Multicellular rosettes have recently been appreciated as important cellular intermediates that are observed during the formation of diverse organ systems. Here, Nechiporuk and colleagues review recent studies of the genetic regulation and cellular transitions involved in rosette formation. They discuss and compare specific models for rosette formation and highlight outstanding questions in the field. See the Review on p. 2549
Heterogeneity and plasticity of epidermal stem cells
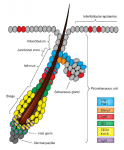 The epidermis is an integral part of our largest organ, the skin, and protects us against the hostile environment. Here, Jensen and co-workers discuss stem cell behaviour during normal tissue homeostasis, regeneration and disease within the pilosebaceous unit, an integral structure of the epidermis that is responsible for hair growth and lubrication of the epithelium. See the Review on p. 2559
The epidermis is an integral part of our largest organ, the skin, and protects us against the hostile environment. Here, Jensen and co-workers discuss stem cell behaviour during normal tissue homeostasis, regeneration and disease within the pilosebaceous unit, an integral structure of the epidermis that is responsible for hair growth and lubrication of the epithelium. See the Review on p. 2559
 (No Ratings Yet)
(No Ratings Yet) (1 votes)
(1 votes)



 (5 votes)
(5 votes)

{kind=link}
{kind=link}
{kind=link}