Here is June’s round-up of some of the interesting content that we spotted around the internet:
News & Research:
– Fly developmental geneticist Walter Gehring sadly passed away this month. His influential research career included the discovery of the homeobox genes.
– This last month saw new updates in the STAP case controversy- Haruko Obokata agreed to retract both papers, while a committee investigating the case has made recommendations that could potentially lead to the closing down of the RIKEN Centre for Developmental Biology.
– Jordan passed a law to control research and therapy using human stem cells derived from embryos, the first such legislation in the region. In Nature, Elena Cattaneo and Gilberto Corbellini wrote about their fight against pseudoscience in stem cell treatments in Italy.
– Mosaic, published by the Wellcome Trust, published a great piece this month about ageing and development.
– Model organisms at the MPI-CBG in Dresden are not just used for research but also to predict footbal scores- here are the scientific oracles for this World Cup! [in German] And if you are following the World Cup, here is how football compares with a PhD by PHD comics.
– And the Weigel lab (Max Planck Institute for Developmental Biology in Tübingen, Germany) continues the tradition of science-themed parodies of popular songs by releasing their ‘Weigel style’ video as part of their take over of the eLife twitter account:
Keep up with this and other content, including all Node posts and deadlines of coming meetings and jobs, by following the Node on Twitter.
Once upon a time, physicists got curious about the cytoskeleton. They characterised the cytoskeleton – using tools of soft matter, statistical and polymer physics – as a mesoscale material whose physical properties govern its dynamics. They showed that the cytoskeleton is an interconnected scaffold that, depending on the time scale, can behave like a shape-morphing gel (slow) or like an elastic polymer network (fast)1. Concurrently, biochemists showed how molecules interact to mediate the architecture of this scaffold. Together, biochemists and physicists have illustrated an emergent picture of comprehensive cytoskeletal function; they enabled a fuller comprehension of, for example, cell migration2, maintenance of cell shape3, and morphogenesis4. Essentially, the two lines of research linking the molecular scale to the polymer scale in the cytoskeleton have amplified one another in an entirely co-dependent manner.
Interestingly, if the cytoskeleton exists solely to maintain cell integrity, and drive migration and molecular traffic flow, one would think cytoskeletal polymers would begin and end at the cellular surface. However, they do not: they are vitally connected, through nesprins, lamins and the LINC complex5,6, to the nucleus and the genetic machinery of the cell7. Why is the genetic machinery of the cell coupled to forces from the outside world? This is an especially vexing question to ask of embryonic stem (ES) cells, given that they lack the Lamin A/C protein8 that largely provides the nucleus its mechanical integrity9. Observe below the dramatic shape changes of the ES cell nucleus within one minute (click to play):
These shape changes are generated by the cytoskeleton. However, some might say the cell is devoted to a pre-ordained genetic program independent of the outside world modulo chemical signaling. Then why does the nucleus coordinate its shape with cytoskeletal dynamics? One potential explanation is mechanotransduction, which posits that forces from the outside world are transduced into changes in cell function. An idea within the mechanotransduction framework is that cytoskeleton-driven nuclear shape changes and subsequent rearrangements of chromatin precipitate changes in gene expression. It is now high time – in the days of CHiP and HiC highlighting chromosomal molecular interactions10,11 – for physicists to do for the nucleus what they have done for the cytoskeleton: provide a material foundation.
Against this backdrop, we set out to understand how ES cell nuclei respond to forces.
Specifically, we wished to know how nuclei of ES cells exiting pluripotency respond to forces. We investigated mouse ES cells maintained in a naïve pluripotent state using inhibitors described elsewhere12. By removing the inhibitors from the medium, ES cells begin to differentiate, and after approximately one day in transition, they exit pluripotency and prime for differentiation. We were especially interested in comparing these transition ES cells – the ones that are exiting the pluripotent state – to naïve pluripotent ES cells. Details of how we defined naïve and transition ES cells can be found in our recent work published in Nature Materials10.
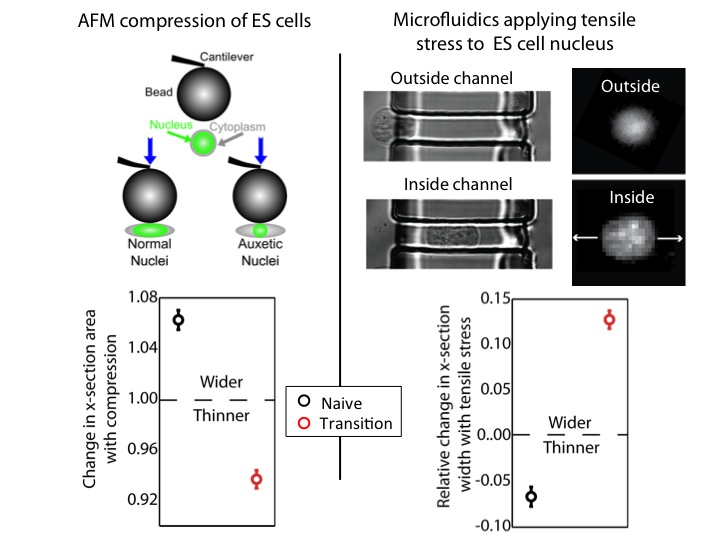
We used an atomic force microscope (AFM) to compress naïve and transition ES cells. We found that the nuclei of transition ES cells, in contrast to naïve ES cells, shrank in cross-section upon compression. This counters expectations. If one presses, for instance, on a ball, couch cushion, or plastics, they would be surprised if these items shrank in cross-section when they press. Nevertheless, that’s what happens to the transition ES cell nucleus.
For further exploration, we tested what happens when tensile stresses are applied via the cytoskeleton. We developed a microfluidic system comprising channels smaller in cross-section than ES cells, but larger than average nuclei. The cytoplasm elongates, and applies tensile forces to the nucleus. Ordinarily, with applied tensile force a material, such as a rubber band, becomes thinner in cross-section and approximately conserves volume. This is what occurs with naïve nuclei, and nuclei of other cell types we investigated (including lineage primed cells). However, transition nuclei – against anticipation – expand in cross-section with applied tensile stress; indeed, the nuclear volume increases by up to 50%.
At left, we show that by compressing transition, but not naive, ES cells with AFM, we see that the nucleus shrinks. At right, we show that by using our microfluidic system to stretch cells and apply tensile forces to the nucleus, the nucleus gets wider in cross-section in transition ES cells, but not naive ES cells.
At left, we show that by compressing transition, but not naive, ES cells with AFM, we see that the nucleus shrinks. At right, we show that by using our microfluidic system to stretch cells and apply tensile forces to the nucleus, the nucleus gets wider in cross-section in transition ES cells, but not naive ES cells.
A material that expands in cross-section when stretched and shrinks in cross-section when compressed is called auxetic13. It is an unusual material property (one must imagine a rubber band getting fatter when stretched) not generally observed in biological materials. It is used for materials like bulletproof vests and soundproofing, due to its ability to immediately absorb impact energy throughout the material instead of just at the impact point. It is also a useful property for materials used for absorption – like sponges – due to the large volume gain and consequent opening of space resulting from a stretch (consider a sponge mop that is pulled, expanding potential fluid pockets in the material).
We investigated possible mechanisms for this unusual nuclear property. We looked for differences in nuclear envelope proteins such as Lamin B1 and Nesprins, but found no meaningful difference between transition and naïve ES cells. However, electron microscopy images showed that transition nuclei were more granular and less dense than naïve nuclei. Therefore, we induced chromatin decondensation using Trichostatin A (TSA)14. With TSA, we provoked an auxetic nucleus in naïve ES cells. This led us to conclude that auxetic nuclei are due, at least in part, to increased global chromatin decondensation in transition ES cells. This is a relatively surprising result from a biological perspective, given the conventional wisdom that naïve ES cells are in state of minimum global chromatin decondensation. However, given that auxetic materials tend to be low density, it was entirely predictable from the standpoint of materials science.
Finally, we addressed the potential function of an auxetic nucleus. We suspected that the function might lie in the dramatic absorptive properties implied by auxeticity. To explore this potential function, we first loaded both transition and naïve ES cells with fluorescein, which in cells becomes more highly concentrated in the cytoplasm than the nucleus. We used our microfluidic technique to apply tensile stresses to the nucleus, and observed fluorescein flowing from the cytoplasm into the nucleus in the transition ES cells, but not in the naïve ES cells. This result indicates that auxeticity opens up porous space in transition nuclei. Coming back to the possible function of an auxetic nucleus – and auxetic sponges – we now hypothesise that auxeticity may prove to be a mode of mechanotransduction. Tensile stresses would lower the free energy barrier associated with nuclear translocation, delivering differentiation specific signaling factors from the cytoplasm to the nucleus in the differentiating ES cell. This is an especially compelling idea given the transition of epiblast from a compressed epithelial phenotype to a spread mesenchymal phenotype after implantation, when they are primed for differentiation.
The idea of a nuclear sponge facilitating an increased differentiative capacity is an enticing prospect for the function of auxeticity, but there are other possibilities. We noticed with our AFM studies that transition ES cells (not naïve ES cells) stiffened with increased compression, as one would expect with an auxetic material. Possibly, this stress-driven stiffening (and conversely softening with tensile stress) could have a significant effect on dynamic sorting of nascent tissue layers within the developing embryo.
Ultimately, there is much work to be done to understand the how and why of auxetic nuclei in ES cells. It is, however, exemplary of new discoveries in biology made by applying physics to biological systems. We hope this discovery will be coupled to emerging awareness of molecular and mechanical signaling in the nucleus to generate a richer comprehension of pluripotency, differentiation, and embryogenesis.
1. Deng, L., Trepat, X., Butler, J., Millet, E., Morgan, K., Weitz, D., & Fredberg, J. (2006). Fast and slow dynamics of the cytoskeleton Nature Materials, 5 (8), 636-640 DOI: 10.1038/nmat1685
2. Gardel, M., Schneider, I., Aratyn-Schaus,, Y., & Waterman, C. (2010). Mechanical Integration of Actin and Adhesion Dynamics in Cell Migration Annual Review of Cell and Developmental Biology, 26 (1), 315-333 DOI: 10.1146/annurev.cellbio.011209.122036
3. Paluch, E., & Heisenberg, C. (2009). Biology and Physics of Cell Shape Changes in Development Current Biology, 19 (17) DOI: 10.1016/j.cub.2009.07.029
4. Lecuit, T., Lenne, P., & Munro, E. (2011). Force Generation, Transmission, and Integration during Cell and Tissue Morphogenesis Annual Review of Cell and Developmental Biology, 27 (1), 157-184 DOI: 10.1146/annurev-cellbio-100109-104027
5. Lombardi, M., Jaalouk, D., Shanahan, C., Burke, B., Roux, K., & Lammerding, J. (2011). The Interaction between Nesprins and Sun Proteins at the Nuclear Envelope Is Critical for Force Transmission between the Nucleus and Cytoskeleton Journal of Biological Chemistry, 286 (30), 26743-26753 DOI: 10.1074/jbc.M111.233700
6. Swift, J., Ivanovska, I., Buxboim, A., Harada, T., Dingal, P., Pinter, J., Pajerowski, J., Spinler, K., Shin, J., Tewari, M., Rehfeldt, F., Speicher, D., & Discher, D. (2013). Nuclear Lamin-A Scales with Tissue Stiffness and Enhances Matrix-Directed Differentiation Science, 341 (6149), 1240104-1240104 DOI: 10.1126/science.1240104
7. Kind, J., Pagie, L., Ortabozkoyun, H., Boyle, S., de Vries, S., Janssen, H., Amendola, M., Nolen, L., Bickmore, W., & van Steensel, B. (2013). Single-Cell Dynamics of Genome-Nuclear Lamina Interactions Cell, 153 (1), 178-192 DOI: 10.1016/j.cell.2013.02.028
8. Pagliara, S., Franze, K., McClain, C., Wylde, G., Fisher, C., Franklin, R., Kabla, A., Keyser, U., & Chalut, K. (2014). Auxetic nuclei in embryonic stem cells exiting pluripotency Nature Materials, 13 (6), 638-644 DOI: 10.1038/nmat3943
9. Pajerowski, J., Dahl, K., Zhong, F., Sammak, P., & Discher, D. (2007). Physical plasticity of the nucleus in stem cell differentiation Proceedings of the National Academy of Sciences, 104 (40), 15619-15624 DOI: 10.1073/pnas.0702576104
10. Bickmore, W., & van Steensel, B. (2013). Genome Architecture: Domain Organization of Interphase Chromosomes Cell, 152 (6), 1270-1284 DOI: 10.1016/j.cell.2013.02.001
11. Nagano, T., Lubling, Y., Stevens, T., Schoenfelder, S., Yaffe, E., Dean, W., Laue, E., Tanay, A., & Fraser, P. (2013). Single-cell Hi-C reveals cell-to-cell variability in chromosome structure Nature, 502 (7469), 59-64 DOI: 10.1038/nature12593
12. Ying, Q., Wray, J., Nichols, J., Batlle-Morera, L., Doble, B., Woodgett, J., Cohen, P., & Smith, A. (2008). The ground state of embryonic stem cell self-renewal Nature, 453 (7194), 519-523 DOI: 10.1038/nature06968
13. Evans, K.E., & Alderson, A. (2000). Auxetic Materials: Functional Materials and Structures from Lateral Thinking! Advanced materials, 12 (9), 617-628
14. Chalut, K., Höpfler, M., Lautenschläger, F., Boyde, L., Chan, C., Ekpenyong, A., Martinez-Arias, A., & Guck, J. (2012). Chromatin Decondensation and Nuclear Softening Accompany Nanog Downregulation in Embryonic Stem Cells Biophysical Journal, 103 (10), 2060-2070 DOI: 10.1016/j.bpj.2012.10.015
This story starts with me shaking and nervous and stumbling through my mid-term thesis committee meeting presentation. Not only was I presenting the results from the first half of my PhD but on top of that, I was presenting this work to Francois Guillemot, basically a legend in the field of developmental neurobiology. Luckily for me, my nervousness was completely unfounded and I quickly learned that Francois is not only incredibly knowledge but also very friendly and not at all intimidating. I presented the data I obtained that demonstrates an essential role for microRNA-9 (miR-9) in regulating the quiescent state of neural stem cells (NSCs) in the adult zebrafish pallium. After providing me with a lot of helpful insight and advice he generously offered to have me come to his lab to look at miR-9 in the NSCs of the adult mouse brain. Additionally, his lab had recently optimized a protocol to induce quiescence in vitro in primary adult hippocampal derived NSCs (AH NSCs) which would be a great model in which to test the expression of miR-9 in quiescent versus activated NSCs.
I immediately accepted his offer without hesitation but I soon realized that traveling to and staying in London might end up being quite expensive. Luckily for me, the money provided by the Development Travelling Fellowship enabled me to spend 3 weeks in the Guillemot lab at the NIMR in Mill Hill.
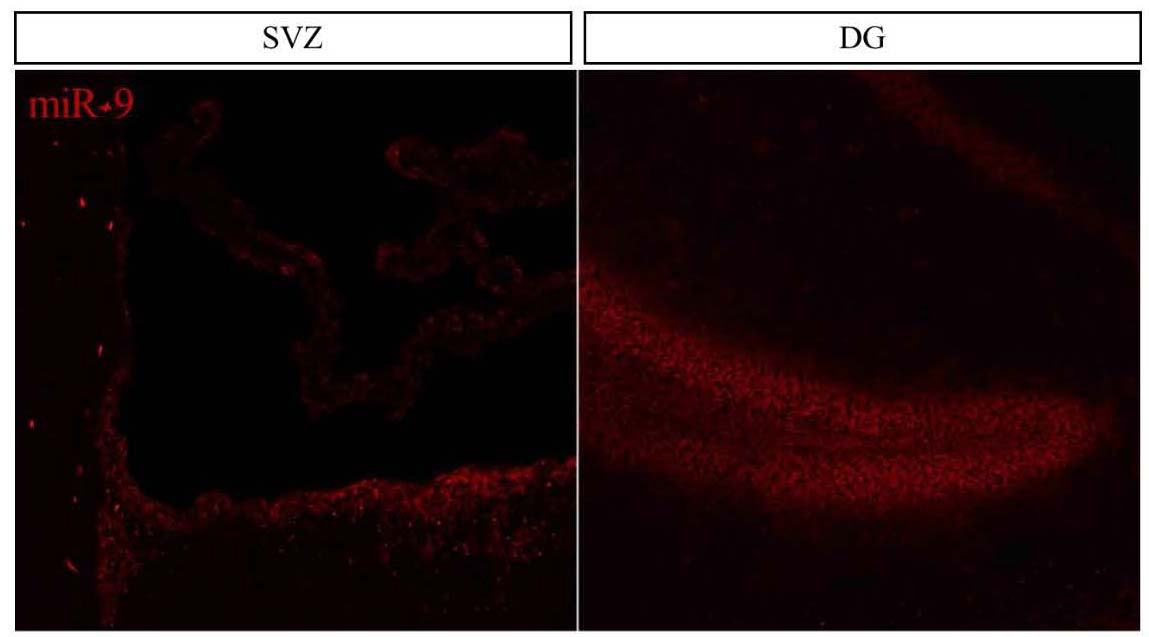
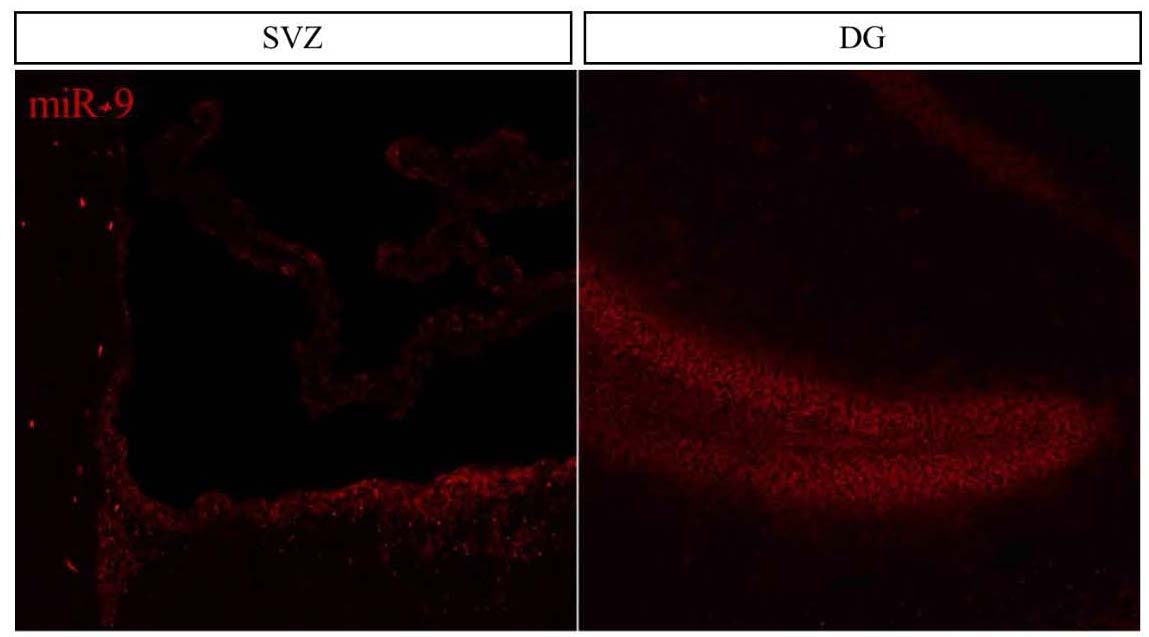
The time I spent at the NIMR in Mill Hill proved to be extremely fruitful and scientifically enriching. Upon my arrival in the Guillemot lab I was greeted by an incredibly friendly group of people and I knew that I would have a great time in the lab. Additionally, all of the mouse samples and cells were ready for me so no time was wasted and I was immediately able to start my search for miR-9 in the mouse brain. Everyone in the lab was incredibly organized and helpful which enabled me to complete a tremendous amount of work and in the end I obtained exciting results showing that miR-9 is highly expressed in the dentate gyrus (DG) and subventricular zone (SVZ), the two germinal zones of the adult mouse brain (see figure below), and also that miR-9 is much more abundant in quiescent NSCs compared with proliferating NSCs in vitro. Additionally, the NIMR is an incredibly stimulating scientific environment and I had the good fortune to attend a number of very interesting lab and departmental meetings and seminars. All of this on top of scientific discussions at the pub!
Overall, thanks to the generous Development Travelling Fellowship, I had an incredibly productive and scientifically stimulating 3 weeks in Mill Hill. I was able to obtain results that will be of great benefit to my PhD project, I learned a number of useful techniques and most importantly, I met a bunch of interesting young scientists that I am looking forward to seeing again soon!
A 5 year research assistant position to study regulation of macrophage migration in Drosophila is available in the Evans lab (Infection and Immunity/The Bateson Centre, University of Sheffield). The closing date for applications is 29th June.
Philip Ingham is a geneticist and developmental biologist, based at the Imperial College, London – Nanyang Technological University, Lee Kong Chian School of Medicine in Singapore. From identifying the Drosophila trithorax mutant to the discovery of the vertebrate hedgehog genes, from perfecting in situ techniques in the fly to helping make zebrafish a successful model system, Phil has made significant contributions to the developmental biology field over three decades. In recognition for his career, Phil was awarded the Waddington Medal at the 2014 BSDB Spring meeting, where we had the opportunity to interview him. Phil is also the current president of the International Society of Developmental Biologists (ISDB).
How are you finding this BSDB spring meeting?
Very much as I remember it from the last time I came, 9 years ago. It is great that it is attended by many students and postdocs, with lots of questions after the talks. The programme is quite varied, the quality is very high and there is good international participation.
One of the reasons why you are attending this meeting is to receive the Waddington Medal. What does it mean to you to receive this prize?
The Waddington Medal is the best thing that I could imagine receiving! It is always nice to get recognition for what you do, but to receive an accolade from this society means everything to me. I have been associated with the BSDB since 1987, and I have been attending their meetings since I was a student. In fact, one of the earliest talks I ever gave was at the BSDB meeting in Sussex in 1982. So getting this award is really thrilling.
There is a rumour that before being a scientist you were a priest. This isn’t the case, but is it true that you didn’t always intend to be a geneticist?
I never intended to be a geneticist. When I went to university I don’t think I would have been able to tell you what genetics was! I had no knowledge of, and no interest in, biological sciences. I went up to Cambridge with the intention of reading social psychology, but as you couldn’t study that in your first year, I did religious studies and theology, as they were the closest related subjects. An important component of the course was looking at the effects of scientific thought on religious belief, especially Darwinism, and that is when I started thinking and reading about evolution. Then I wandered into a Biology of Cells lecture one day and saw a movie of mitosis, which I found spell-binding. That is when I decided I wanted to do biological sciences. Because of my background, the only subject I could study was genetics. You didn’t really need to know any biology to study genetics!
You were very fortunate to be invited to the exclusive EMBO Drosophila workshop in Crete when you were still a PhD student. How did this happen and what opportunities did it give you?
It was a fantastic opportunity. It happened because I was naïve enough to apply to it, as I didn’t realise what the rules and restrictions were. The fact that I had just submitted my first paper on the discovery of trithorax (Ingham and Whittle, 1980) to Molecular and General Genetics (now Molecular Genetics and Genomics) and Walter Gehring, who was also chairing the workshop selection committee, was handling it, probably had some impact. It was a chastening experience actually, as I had to give a short talk without preparation or slides. Most importantly, I got to meet several people who I still know to this day and with whom I have interacted very closely, such as Janni Nüsslein-Volhard, Eric Wieschaus and David Ish-Horowicz. David in particular was very kind and friendly towards me, and this played a part in my later decision to apply to his lab. Going to the Crete meetings is a fantastic way to get a clear overview of the cutting edge research that is going on, and all the leading people were there.
You mentioned that it was at this meeting that you first met Janni Nüsslein-Volhard, who later persuaded you to change from Drosophila to zebrafish. Why did you decide to focus on vertebrate development and was this a difficult transition?
Janni is a very powerful influence! She became interested in my work when I moved to David’s lab and started working on segmentation. In 1986 I was working at the LMB (MRC Laboratory of Molecular Biology, Cambridge, UK) and collaborating with Gary Struhl on the molecular analysis of one of Janni’s favourite mutants, even skipped. But when I visited her in Tübingen with the intention of showing her my data, she was much less excited than I expected, because she had already decided that fish was the next big thing.
She persuaded me that Drosophila was pretty much solved and that it was time to move on to a genetically tractable vertebrate model organism. I really am a geneticist, so the idea of having a new organism, starting afresh with a blank sheet of paper as I had done with flies nearly 10 years earlier, was very appealing. In addition, I knew I had the option of moving back to the newly established (but now defunct) Developmental Biology Unit of the Imperial Cancer Research Fund in Oxford, which was very vertebrate orientated, and I could see how working on fish would fit well there.
In retrospect, I think it probably wasn’t a good idea for me to start working on fish then, because it diverted my pretty limited resources away from flies. Compared with flies, fish can be incredibly frustrating: you can see the potential for doing experiments due to their beautiful embryology, but it just takes forever to do them. Although the fish community is generally very generous at exchanging material, we just don’t have the huge resources that exist in flies, and the genetics of fish is relatively primitive even now. So you can spend a lot of time creating mutants, transgenics and antibodies, and often, by the time you are ready to do the experiment, it is no longer relevant. That said, I think we are at the dawn of a new era when zebrafish will come of age as a powerful system for disease modelling and screening for novel therapeutics.
Your collaboration with Cliff Tabin and Andy McMahon was very successful, identifying the vertebrate hedgehog genes. Why do you think it was such a productive collaboration?
One simple answer is that Andy and Cliff are incredibly driven people. Whenever they set out to do something it is going to work, so being involved in a collaboration with them is almost a guarantee of success! However, once we found the hedgehog homologues and did the initial characterization, the collaboration fell apart almost as quickly as it started. Cliff has spoken at some length about this collaboration (Tabin, 2009), and in Kathy Weston’s book Blue Skies and Bench Space (Weston, 2013) he says that it is a shame that we didn’t collaborate more: a sentiment which I share. There were some tensions between our labs, but the three of us got on extremely well together. Through that collaboration they probably became two of my best scientific friends.
I believe there was some disagreement in naming one of the homologues Sonic hedgehog…
I didn’t know about the computer game, so I didn’t grab the significance: I just thought it was a silly name. People talk about silly names in Drosophila, but fly gene names are always pretty descriptive, whereas Sonic was not descriptive at all. So much so that I actually resisted even mentioning the name Sonic in our paper. In retrospect, this was probably a mistake, because our paper doesn’t get noticed as much since the name isn’t in the title. And it is one of the favourite gene names out there: Lewis Wolpert once commented ‘Phil, the one gene that my students remember is Sonic hedgehog’!
After spending most of your scientific career in the UK you moved to an A*STAR institute in Singapore. How does scientific research in Asia compare with that in Europe?
I don’t think it is as different as people imagine, but I can’t answer that question with any great authority since, although I have visited several Asian countries, I have only actually worked in one. Considering Singapore, the first thing to say is that the A*STAR institutes are very similar to Western institutes. The IMCB (Institute for Molecular and Cellular Biology) [the institute to which Phil first moved in Singapore] in particular is modelled on the LMB, so much so that when I arrived I really felt that I had gone back home! There is a certain level of topdown management of research strategy, but I don’t think it is that different from the way strategies are determined in Europe or the US – there are the same demands on politicians to justify the spending that they put into research. Maybe in Singapore there is a more extreme version of it, strategies are implemented more strictly, and things are expected to happen more quickly. For instance, the transition in A*STAR from the honeymoon period when you could do anything to having to do industrially relevant research was very sudden. It happened over a matter of months, while in Europe and the US it might have happened over a number of years. But in terms of the day-to-day practice of science, it is not so different.
You recently became the president of the ISDB. What are you aiming to achieve during your presidency?
The principal activity of the ISDB is organising the ISDB Congress every 4 years; we’ll be holding the next one in 2017 in Singapore. However, my predecessor Claudio Stern has really increased the profile and the activities of the ISDB. He has been very energetic in going around the world encouraging national communities to organise themselves into societies, and coming up with various funding schemes, such as the ISDB–Mechanisms of Development lecture. Claudio is quite a hard act to follow. I am going to try to follow his lead and continue to help communities to establish societies where they don’t exist – not least in Singapore, where we still only have a developmental biology ‘club’!
During your career you were not afraid to change lab, institute and even model organism to pursue your scientific interests. What is your advice for young scientists when making their career choices?
The best advice in science is always to do what you find really interesting. Maybe circumstances are very different now, and you do have to be a bit more directed and strategic. However, I can honestly say that I never thought about my career at all – I just did what I found interesting! Maybe I have just been incredibly lucky, but it is an approach that has paid off. I remember when I joined the ICRF (Imperial Cancer Research Fund) that I was told in no uncertain terms by other postdocs suspicious of a fly geneticist that there was no way I would ever get a position there – but then I spent 10 years of my career at the ICRF, ultimately as a Principal Scientist. I don’t think it is a good idea to try to follow trends and second guess what is going to put you in a ‘marketable’ position. Science requires so much hard work and dedication that if you are not passionate about what you are doing, you are not going to succeed.
What would people be surprised to find out about you?
I come from a strong agricultural background and am the only member of my family to have graduated from university. My grandfather was a dairyman and cattle dealer, and my happiest days as a teenager were spent working on my cousin’s arable farm outside Liverpool, picking potatoes and making hay! I also have several farming cousins in Massachusetts who I try to visit whenever I am in Boston. Also, I played guitar in a rock band when I was younger. My youngest daughter is a very good guitarist, so I still jam with her occasionally.
Department/Location: Wellcome Trust – Medical Research Council Cambridge Stem Cell Institute
Salary: £28,132-£36,661
Reference: PS03564
Closing date: 22 July 2014
Limited funding: The funds for this post are available until 30 June 2017 in the first instance.
The Stem Cell Institute is a world-leading centre of excellence in stem cell biology and regenerative medicine, supported by a strategic funding partnership between the Wellcome Trust and the Medical Research Council (www.stemcells.cam.ac.uk).
The Institute comprises over 300 researchers spanning fundamental science through to clinical applications. Our vision is of a deep understanding of stem cell biology for the prevention and treatment of human disease.
We are seeking a dynamic, innovative and self-motivated individual with the skills and experience to enhance the performance and profile of the Stem Cell Institute. The Coordinator will assist the Director and the Institute Steering Committee to optimise communication and connectivity within the Institute and to achieve greater identity and visibility.
The Coordination Officer will put in place a strategy to enhance the profile of the Institute with the research community, sponsors, policy makers and the media. They will work closely with the Institute’s Public Engagement Officer to ensure that there is effective and professional communication with both our internal and external stakeholders.
The role holder will be responsible for organising and facilitating communication, networking and information exchange within the Stem Cell Institute, which is currently spread across several sites in Cambridge. Therefore the role holder must demonstrate a proven track record in relationship building, report writing, event organisation and data management. The Coordination Officer will also assist in preparing the Institute’s annual reports and securing renewed funding for the Institute, and will be secretary to the Institute Steering Committee.
You will have outstanding organisational and administrative experience and be comfortable working to tight deadlines with minimal supervision. You should have demonstrable experience in web-based/social media communication and you should have excellent written and verbal communication and negotiation skills. You will be IT literate and able to work both on your own and as part of a team.
The post will report to the Institute Director. You will have a degree (or equivalent). Experience of working in the HE or research sector in a senior administrative role would be an advantage.
Once an offer of employment has been accepted, the successful candidate will be required to undergo a health assessment.
To apply online for this vacancy and to view further information about the role, please visit: http://www.jobs.cam.ac.uk/job/4160. This will take you to the role on the University’s Job Opportunities pages. There you will need to click on the ‘Apply online’ button and register an account with the University’s Web Recruitment System (if you have not already) and log in before completing the online application form.
Please upload your current CV and cover letter with your application by Tuesday 22nd July 2014.
Interviews will be held at the beginning of August 2014. If you have not been invited for interview by 31st July 2014, you have not been successful on this occasion.
Please quote reference PS03564 on your application and in any correspondence about this vacancy.
The University values diversity and is committed to equality of opportunity.
The University has a responsibility to ensure that all employees are eligible to live and work in the UK.
Here are the highlights from the current issue of Development:
Insights into familial dysautonomia
Familial dysautonomia (FD) is a germline autosomal recessive disease that is characterized by impaired peripheral sensory and sympathetic neuron development. The disease is known to be caused by mutations in the gene encoding Elp1 (also known as IKBKAP), but how Elp1 functions in neurons is unclear. Now, Warren Tourtellotte and colleagues investigate the role of Elp1 in mice (p. 2452). The researchers first generate conditional knockout mice in which Elp1 is ablated from the neural crest progenitors that give rise to sympathetic and sensory neurons. They demonstrate that the loss of Elp1 in these progenitors has no effect on their migration, proliferation, cell fate specification or survival. By contrast, target tissue innervation was perturbed following Elp1 ablation in neural crest progenitors, leading to increased apoptosis of post-migratory sympathetic and sensory neurons. Furthermore, they report that the ablation of Elp1 in post-migratory sympathetic neurons disrupts tissue innervation, and this is associated with attenuated axon branching. In line with this, the authors demonstrate that Elp1-depleted sympathetic and sensory neurons exhibit impaired neurite outgrowth and altered tubulin dynamics, suggesting a role for Elp1 in cytoskeletal regulation. These and future studies of this new mouse model for FD offer promising insights into the role of Elp1 in neural development and disease.
New atlas-builder software and the eNeuro atlas
The recent advent of tools for manipulating and monitoring gene expression calls for efficient ways to document, access and analyse these gene expression patterns. Although a number of databases and gene expression atlases have been compiled in recent years, many of them are limited with regards to their content and utility. Here, Chris Doe and colleagues develop new software that overcomes these limitations (p. 2524). This new ‘atlas-builder’ software can be used to create an atlas of gene expression in any tissue in any organism with stereotyped cell positions. Importantly, they report, the atlases generated by this software are three-dimensional, allow for the registration of an infinite number of markers, are searchable and are open-ended; additional markers can be added by users. To validate the software and to help demonstrate its advantages, the authors generated an ‘eNeuro’ atlas of the Drosophilaembryonic CNS. The authors initially populated the atlas with eight transcription factors that mark the major CNS cell types. The atlas was subsequently expanded to include data from 75 Gal4 lines expressed in sparse patterns, thereby allowing the identification of molecularly distinct subsets of interneurons and revealing unexpected diversity among motor neurons. The ‘atlas-builder’ software and the eNeuro atlas, both of which have been made publicly available, promise to be valuable resources for the developmental biology community.
Sp(1)ecifying haematopoietic cells
Haematopoiesis – the formation of blood cells – is regulated by a number of ubiquitous and tissue-specific transcription factors, but the extent of interplay between these factors is unclear. Sp1 is a transcription factor that is ubiquitously expressed and regulates the expression of thousands of genes, and it has been shown that Sp1-deficient mouse embryos die during early development. Now, on p. 2391, Sjaak Philipsen, Constanze Bonifer and colleagues reveal a crucial role for Sp1 during the early stages of haematopoiesis. Using mouse embryonic stem cells (ESCs) that express a DNA binding-deficient variant of Sp1, the researchers first show that Sp1 activity is required for the differentiation of ESCs to hematopoietic lineages; the cells can progress through most steps of blood cell development but are unable to complete terminal differentiation. Furthermore, they demonstrate that gene expression in Sp1-deficient ESCs becomes progressively deregulated as they differentiate. In particular, they report, some Cdx and Hox family genes that are direct targets of Sp1 are downregulated at an early stage of differentiation, and this is followed by the progressive deregulation of other genes that are implicated in haematopoiesis, suggesting that the effects of Sp1 deficiency are cumulative. Together, these findings identify a crucial role for Sp1 during haematopoiesis and provide detailed insight into the hierarchy of the transcriptional network that drives blood cell formation.
Getting to the root of nodule development
Root nodulation in plants is a form of de novoorganogenesis and involves the dedifferentiation of root cortical cells in response to rhizobia-derived factors. However, due to the complexity of this event, our understanding of the factors and mechanisms that initiate nodule formation is limited. Now, Takuya Suzaki and co-workers (p. 2441) reveal a role for endoreduplication during the onset of nodule development in Lotus japonicus. The researchers identify novel nodulation-deficient mutants that harbour mutations in VAG1, which encodes a protein that is orthologous to a component of the Arabidopsistopoisomerase VI complex that has been implicated as a regulator of endoreduplication. In line with this, the authors report that the number of endoreduplicated cells in vag1 mutant nodules is reduced compared with that of controls. Importantly, the analysis of nuclear size suggests that VAG1-mediated endoreduplication is crucial for the initiation of nodule formation. Finally, the researchers demonstrate that infection threads, the specialized structures used by rhizobia to invade host cortical cells, elongate towards endoreduplicated cells, and this directional elongation is perturbed in vag1 mutants. These findings highlight an essential role for endoreduplication during root nodule development and suggest that VAG1-mediated endoreduplication is required for the efficient guidance of symbiotic bacteria to host cells.
PLUS…
An interview with Phil Ingham
Philip Ingham is a geneticist and developmental biologist, based at the Imperial College, London – Nanyang Technological University, Lee Kong Chian School of Medicine in Singapore. Phil has made significant contributions to the developmental biology field over three decades and, in recognition of these achievements, was awarded the Waddington Medal at the 2014 BSDB Spring meeting, where we had the opportunity to interview him. See the Spotlight article on p. 2363
How to make a hippocampal dentate gyrus granule neuron
Granule neurons in the hippocampal dentate gyrus (DG) are known to be continuously generated throughout adult life, and the ongoing integration of newborn neurons into the existing hippocampal neural circuitry provides enhanced neuroplasticity, which plays a crucial role in learning and memory. In their Primer article, Gage and colleagues summarize the developmental principles that regulate the process of DG neurogenesis and discuss recent advances in harnessing these developmental cues to generate DG granule neurons from human pluripotent stem cells. See the Primer on p. 2366
Chromatin features and the epigenetic regulation of pluripotency states in ESCs
In pluripotent stem cells, the interplay between signaling cues, epigenetic regulators and transcription factors orchestrates developmental potency. Here, Maria-Elena Torres-Padilla and Ian Chambers review what is known about transcriptional heterogeneity in pluripotent stem cells, focusing on the underlying causes of heterogeneity and how transcriptional heterogeneity can be to the benefit of the whole stem cell population. See the Review on p. 2376
Many congratulations to Nathan Kenny (University of Oxford), Kathryn McClelland (Institute for Molecular Bioscience, University of Queensland), and Sophie Miller (University of Cambridge), who took this image at last year’s Woods Hole Embryology course. The image shows a squid (Loligo pealeii) embryo stained with anti-acetylated tubulin (red), anti-sorotonin (green), and DAPI (blue,nuclei).
The runners-up to this competition were the annelid larvae by Poulomi Ray (Clemson University), the live squid embryo by Brijesh Kumar (Indian Institute of Technology, Kanpur) and the squid eye by Ezgi Kunttas (Carnegie Mellon University).
The winning squid embryo image will feature in the cover of a coming issue of Development. This was only the first round though, so expect another set of beautiful images from the Woods Hole embryology course here on the Node very soon!
June 1st, 2014: Exactly one year after my departure flight from Bologna to Boston to attend the 2013 MBL Embryology course held at the Marine Biological Laboratory in Woods Hole, MA (https://thenode.biologists.com/six-weeks-in-woods-hole/events/), I was again in the Bologna airport, only that this time I was landing from the States.
I was returning to my home institution, the University of Modena and Reggio Emilia, Italy where I am a last year graduate student, from the Stowers Institute for Medical Research in Kansas City (MO, USA), where I spent the last 3 months working in the laborartory of Prof. Alejandro Sánchez Alvarado.
Thanks to the 2013 MBL Embryology course, where I met Prof. Sánchez Alvarado (Director of the course together with Prof. Richard Behringer), and thanks to the Development Travelling Fellowship, I could collaborate with a great research group in one of the best places anywhere in the world that a biologist can work at: the Stowers Insitute (http://www.stowers.org/stowers-report/fall-2013/best-places-to-work).
At the time of the 2013 MBL Embryology course, I was working mainly on the immune functions of the freshwater gastropod, Pomacea canaliculata, a mollusk which is considered a dangerous pest already invading North America and South-East Asia from the South America. Pomacea is also the intermediate host of the nematode Angiostrongylus cantonensis, which causes eosinophilic meningitis in humans. This snail is aggressively invading new territories because of its adaptability to different kinds of environmental conditions and resistance to multiple stress conditions. As such, the characterization of the immune system of this snail is important for controlling its diffusion by allowing to uncover features that may be vulnerable to intervetions for eradication.
However, after the 2013 MBL Embryology course in Woods Hole, I became highly interested in regeneration and the incredible mechanisms able to activate either stem or differentiated cells in order to increase the proliferative rate and build either amputated parts or whole organisms altogether. More than two hundred years ago, Lazzaro Spallanzani’s experiments (in part performed in Modena, by the way) showed that terrestrial snails are able to regenerate their heads after decapitation. In Woods Hole, I was really impressed by the regenerative capacity of planarians, which can be included with mollusks and other groups into the lophotrocozoan taxon. Because of this, when I returned to Modena in 2013, I tried and cut the sensory organs of P. canaliculata in order to verify if it is able to regenerate.
I was very excited when I observed that after 2, 3 and 4 weeks, respectively, the cephalic and oral tentacles, as well as the eyes were completely regenerated. This observation formed the basis for my period in Kansas City as a visiting student.
Before I left for Kansas City, I collected the samples of the regenerating cephalic tentacles, oral tentacles and eyes of Pomacea canaliculata at different time points. Once at the Stowers Institute, the RNA purification and sequencing of the samples and the bioinformatic analysis of the data allowed for the construction of a transcriptome database. A high-resolution time-course was produced for each organ and the analysis of the gene expression profiles uncovered a significant number of genes differentially expressed during the regeneration processes surveyed. The availability of the transciptome of these complex organs will allow for detailed molecular analyses of the pathways involved in regeneration, providing a solid foundation for my future studies.
These three months at the Stowers Institute, which directly stemmed from the 2013 MBL Embryology course, gave me the chance to write a personal three-month project, learn new techniques, work with a planarian model, perform experiments in the field of regeneration, and collect data that will be surely useful in my future studies. Last but not least, at the Stowers Institute I met a lot of great scientists, and attended many interesting lectures. Equally enjoyable were my interactions with post-docs from all over the world from whom I benefited extensively through their continuous and kind assistance in my lab activities.
I am confident that this experience will reveal of fundamental importance for me and my scientific career.
Cells move in (still) mysterious ways to achieve morphogenesis.
Prominently, cells of an early vertebrate embryo (blastula, a mass of undifferentiated cells) move extensively during gastrulation to generate the three basic layers of the organism: ectoderm at the surface, endoderm presaging the digestive tube, and the mesoderm in between. At the end of the process, the body plan is clearly visible, its main axes established.
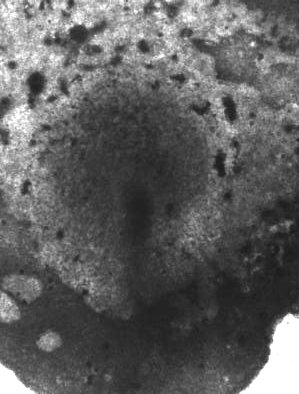
This is perhaps most dramatically illustrated in the extremely large embryos of most amniotes (such as in birds and most mammalians, including the human): they consist of thousands or tens of thousands of cells, arranged in an epithelial disc (epiblast). Gastrulation is achieved through a seemingly fixed structure at the midline, the primitive streak (Figure 1).
Figure 1: Chick embryo at gastrulation. The primitive streak is the darker stricture along the midline.
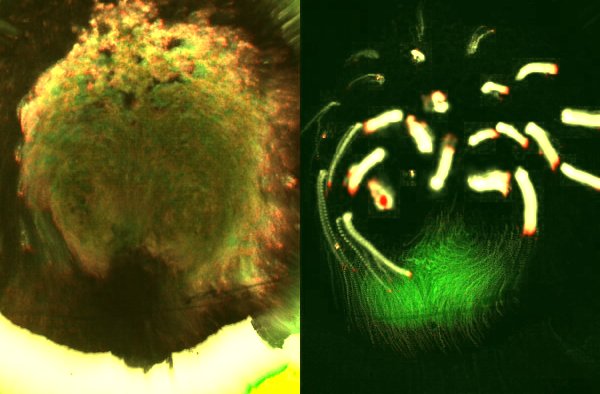
Here, cells engage in epithelial-to-mesenchymal transitions (EMTs, similar to carcinoma progression [1]) to leave the epiblast and contribute to the inner layers. In the chick embryo – a good representative of most amniotes -, virtually all cells in the embryonic disc have long been observed to be engaged in large displacements, directed mainly toward the primitive streak ([2, 3], Figure 2).
Figure 2. Massive movements across the embryonic epiblast. Left, using transmitted light to observe development prior to streak formation. Right, fluorescent marking of the domain where the primitive streak will form (green) and tracks of other groups of cells (yellow; red indicates the direction of movement).
But how is the primitive streak maintained as a fixed structure in the middle of a field of constantly moving cells? How does it form in the first place? To add to the puzzle, massive cell movements in the epiblast actually start well before gastrulation in the form of counter-rotating whorls, the meeting point of which always correspond to where the primitive streak later forms. Is there a causal relationship?
We have now shown [4] that the primitive streak arises by a chain reaction of EMT events. This is primed by rare EMT events, which occur in the epiblast well before gastrulation. EMTs become highly cooperative through a community effect requiring TGF-b signalling. The gene coding for Nodal, a member of this superfamily, is transcribed early in a small domain on the edge of the blastula, but the activity of the protein is antagonised by signals emanating from an amniote-specific extra-embryonic layer, the hypoblast, which initially lines the epiblast. It is its displacement (and the disinhibition of Nodal activity) that triggers the chain reaction of EMTs and, therefore, the initiation and later the maintenance of the primitive streak.
The Nodal expression domain corresponds to a region in which we previously found cell intercalation to occur, in the plane of the epiblast (green in figure 2, right panel) – this is again regulated by the hypoblast, but positively and via different signals (FGFs, [5]). The intercalation region is initially arranged along the edge of the epiblast and intercalation displaces its cells along one of its radii, before gastrulation. So, one key invention of amniotes is a mechanism to regulate Nodal activity and displace it along the future anterior-posterior axis of the embryo, before it can trigger gastrulation. Interestingly, both intercalation and delaying Nodal-dependent chain reaction of EMTs are regulated by the hypoblast, the characteristic evolutionary acquisition of amniotes.
Two key, local cell behaviours therefore drive primitive streak formation. What about the movements in the rest of the epiblast? We turned to computer simulations and we show that these two components can indeed entrain cells in the entire epithelial sheet to move in the correct pattern, and that the model can also represent the experimental conditions of abrogating or ectopically creating one or two of these components. Local actions can trigger global movements.
A few directions now open, which we now pursue in my new lab in Cambridge. There is now a framework for interpreting experiments at molecular or cellular level and linking them quantitatively to large-scale morphogenesis, and refine both experiments and models in step with each other. Such models hold the key not only for understanding large, regulative embryos (including the human), but also for adequately discerning the common developmental patterns and the true acquisitions / losses during evolution, and the constraints that brought them about. Gastrulation also sets the stage for the development of the nervous system and the elongation of the main body axis; we now hope that we have better tools to continue exploring these key issues.
References
[1] Nieto, M. (2011). The Ins and Outs of the Epithelial to Mesenchymal Transition in Health and Disease Annual Review of Cell and Developmental Biology, 27 (1), 347-376 DOI: 10.1146/annurev-cellbio-092910-154036
[2] Gräper, L. (1929). Die Primitiventwicklung des Hühnchens nach stereokinematographischen Untersuchungen, kontrolliert durch vitale Farbmarkierung und verglichen mit der Entwicklung anderer Wirbeltiere Wilhelm Roux’ Archiv für Entwicklungsmechanik der Organismen, 116 (1), 382-429 DOI: 10.1007/BF02145235
[3] Wetzel, R. (1929). Untersuchungen am Hühnchen. Die Entwicklung des Keims während der ersten beiden Bruttage Wilhelm Roux’ Archiv für Entwicklungsmechanik der Organismen, 119 (1), 188-321 DOI: 10.1007/BF02111186
[4] Voiculescu, O., Bodenstein, L., Lau, I., & Stern, C. (2014). Local cell interactions and self-amplifying individual cell ingression drive amniote gastrulation eLife, 3 DOI: 10.7554/eLife.01817
[5] Voiculescu, O., Bertocchini, F., Wolpert, L., Keller, R., & Stern, C. (2007). The amniote primitive streak is defined by epithelial cell intercalation before gastrulation Nature, 449 (7165), 1049-1052 DOI: 10.1038/nature06211
 (1 votes)
(1 votes)
 (No Ratings Yet)
(No Ratings Yet)
 Familial dysautonomia (FD) is a germline autosomal recessive disease that is characterized by impaired peripheral sensory and sympathetic neuron development. The disease is known to be caused by mutations in the gene encoding Elp1 (also known as IKBKAP), but how Elp1 functions in neurons is unclear. Now, Warren Tourtellotte and colleagues investigate the role of Elp1 in mice (p.
Familial dysautonomia (FD) is a germline autosomal recessive disease that is characterized by impaired peripheral sensory and sympathetic neuron development. The disease is known to be caused by mutations in the gene encoding Elp1 (also known as IKBKAP), but how Elp1 functions in neurons is unclear. Now, Warren Tourtellotte and colleagues investigate the role of Elp1 in mice (p.  The recent advent of tools for manipulating and monitoring gene expression calls for efficient ways to document, access and analyse these gene expression patterns. Although a number of databases and gene expression atlases have been compiled in recent years, many of them are limited with regards to their content and utility. Here, Chris Doe and colleagues develop new software that overcomes these limitations (p.
The recent advent of tools for manipulating and monitoring gene expression calls for efficient ways to document, access and analyse these gene expression patterns. Although a number of databases and gene expression atlases have been compiled in recent years, many of them are limited with regards to their content and utility. Here, Chris Doe and colleagues develop new software that overcomes these limitations (p.  Haematopoiesis – the formation of blood cells – is regulated by a number of ubiquitous and tissue-specific transcription factors, but the extent of interplay between these factors is unclear. Sp1 is a transcription factor that is ubiquitously expressed and regulates the expression of thousands of genes, and it has been shown that Sp1-deficient mouse embryos die during early development. Now, on p.
Haematopoiesis – the formation of blood cells – is regulated by a number of ubiquitous and tissue-specific transcription factors, but the extent of interplay between these factors is unclear. Sp1 is a transcription factor that is ubiquitously expressed and regulates the expression of thousands of genes, and it has been shown that Sp1-deficient mouse embryos die during early development. Now, on p.  Root nodulation in plants is a form of de novoorganogenesis and involves the dedifferentiation of root cortical cells in response to rhizobia-derived factors. However, due to the complexity of this event, our understanding of the factors and mechanisms that initiate nodule formation is limited. Now, Takuya Suzaki and co-workers (p.
Root nodulation in plants is a form of de novoorganogenesis and involves the dedifferentiation of root cortical cells in response to rhizobia-derived factors. However, due to the complexity of this event, our understanding of the factors and mechanisms that initiate nodule formation is limited. Now, Takuya Suzaki and co-workers (p. 
 Granule neurons in the hippocampal dentate gyrus (DG) are known to be continuously generated throughout adult life, and the ongoing integration of newborn neurons into the existing hippocampal neural circuitry provides enhanced neuroplasticity, which plays a crucial role in learning and memory. In their Primer article, Gage and colleagues summarize the developmental principles that regulate the process of DG neurogenesis and discuss recent advances in harnessing these developmental cues to generate DG granule neurons from human pluripotent stem cells. See the Primer on p.
Granule neurons in the hippocampal dentate gyrus (DG) are known to be continuously generated throughout adult life, and the ongoing integration of newborn neurons into the existing hippocampal neural circuitry provides enhanced neuroplasticity, which plays a crucial role in learning and memory. In their Primer article, Gage and colleagues summarize the developmental principles that regulate the process of DG neurogenesis and discuss recent advances in harnessing these developmental cues to generate DG granule neurons from human pluripotent stem cells. See the Primer on p.  In pluripotent stem cells, the interplay between signaling cues, epigenetic regulators and transcription factors orchestrates developmental potency. Here, Maria-Elena Torres-Padilla and Ian Chambers review what is known about transcriptional heterogeneity in pluripotent stem cells, focusing on the underlying causes of heterogeneity and how transcriptional heterogeneity can be to the benefit of the whole stem cell population. See the Review on p.
In pluripotent stem cells, the interplay between signaling cues, epigenetic regulators and transcription factors orchestrates developmental potency. Here, Maria-Elena Torres-Padilla and Ian Chambers review what is known about transcriptional heterogeneity in pluripotent stem cells, focusing on the underlying causes of heterogeneity and how transcriptional heterogeneity can be to the benefit of the whole stem cell population. See the Review on p. 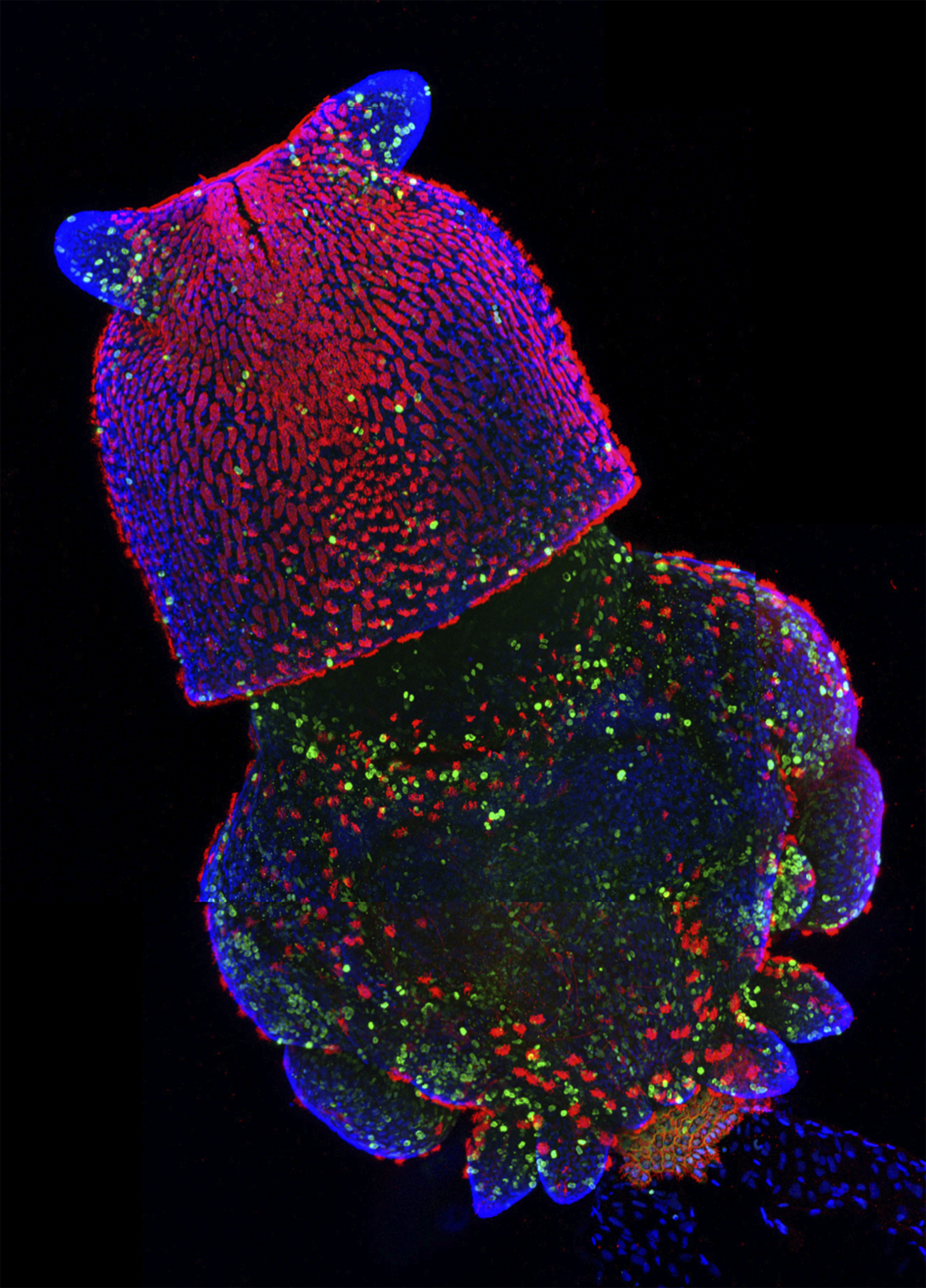
{kind=link}