Yesterday, we went through a couple of examples of amphibian tail regeneration. Limb regeneration is another example of epimorphic regeneration, which is expertly implemented by some amphibians. Unlike the tail, the limb is patterned in three dimensions: anterior-posterior, ventral-distal and media-proximal to produce the proper number and position of the digits, making the patterning process more complex (McQueen and Towers, 2020). But the potential to – one day – regenerate lost human limbs makes limb regeneration a tantalising subject for researchers and science-fiction writers alike (Cox et al., 2019).
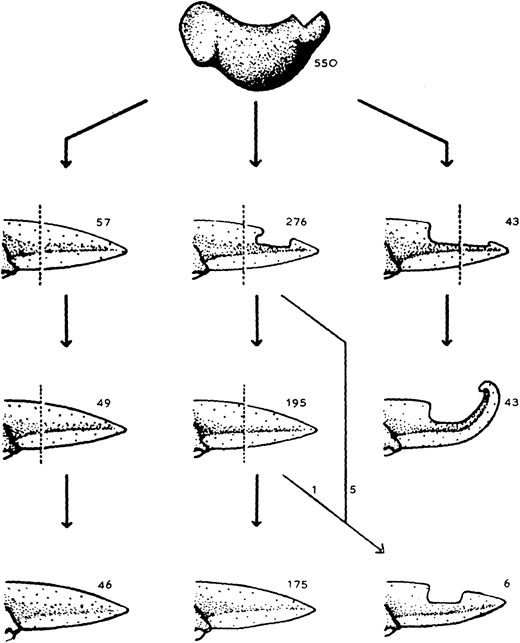
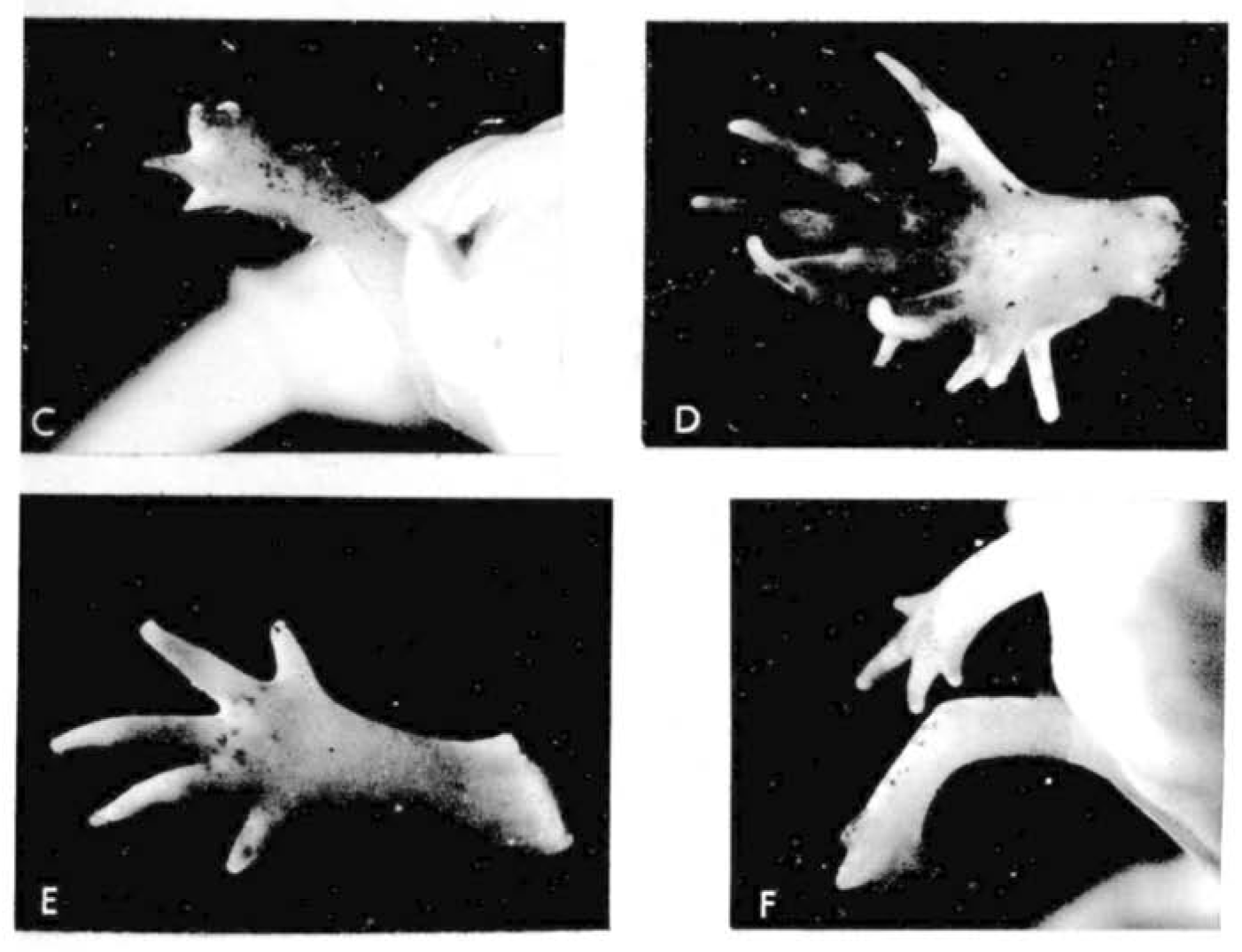
Let us return to the work of our old friend Dr D. R. Newth. In the previous post, we learnt how regeneration could produce a tail more perfect than the original; there’s a similar theme in today’s flashback. A few months after the 1958 publication on tails, Newth posted the second half of his work in JEEM/Development on Triturus helveticus newt limbs (Newth, 1958). There seemed to be an abundance at the time of biologists researching the regenerative abilities of ectopic (supernumerary) limbs by grafting bits of stump tissue, as well as other bits. Such transplants usually produced abnormal limbs with too few or too many digits, which were occasionally resolved during regeneration but there was a lot of conflicting evidence bouncing between journals at the time. One peculiar observation from the past literature is that grafting otic vesicles into the flank induced ectopic limb growth. Newth took a similar approach, engrafting nasal placodes into the body wall to produce an extra foreleg – if anyone has any idea about the mechanism behind placode-dependent limb outgrowth, please do let me know in the comments! This induction approach was quite inefficient (only 19/163 newts grew ectopic limbs) and, like previous reports, these limbs were abnormal with defects ranging from digit-less stumps to polydactyly (Fig. 1). Amputating these defective limbs, however, caused the regeneration of more ‘normal’ looking limbs, with a second amputation of the regenerated limb further improving the phenotype. For example, an ectopic limb that developed with 12 digits, regenerated eight digits following the first amputation and four digits (the normal number) following the amputation of the regenerated limb (Fig. 1). Newth owed these successful regeneration experiments to the amount of limb material, arguing that grafting an inducing centre to form a whole limb was a better approach than grafting stump tissue and provided more material for precise amputation. Again, the position of amputation was deemed significant, but the fascinating mechanism behind this phenomenon remained a mystery.
Fig. 1. Plate 1 from Newth et al., 1958. D shows the original ectopic limb with 10 digits. E shows the regenerated five-fingered limb. F shows the second regenerated limb with, supposedly, four digits.
The limb development and regeneration field has really flourished in the past 65 years and is impossible to adequately summarise here. Evidence, largely from chick and mice, has revealed the many genetic, epigenetic and molecular regulators that control the position of limb budding, outgrowth and digit patterning (McQueen and Towers, 2019). The field has moved in all directions, investigating the roles of the microenvironment, immune system and progenitor populations, as well as their differentiation and lineages, using genome editing, high-resolution imaging and next-generation sequencing technologies. Grafting remains an extremely useful experimental approach (e.g. Vierira et al., 2023) with salamanders, and the axolotl in particular, the main model systems for limb regeneration experiments (Joven et al., 2019). Like in the 1950s, cross-species comparisons are crucial for understanding how tissue regeneration evolved, the factors that control it, and for identifying approaches to induce regeneration in non-regenerative species.
Takeuchi and colleagues published their paper investigating the role of Hox genes during new limb development regeneration last year (Takeuchi et al., 2022; Fig. 2). Their work follows studies in mammals, which have shown that 5′ Hox genes (e.g. Hox13) are crucial for limb development, but whether this requirement is conserved in other tetrapods has not been clear. Takeuchi and team show that Hox13 genes Hoxa13, Hoxc13 and Hoxd13, are expressed both in developing limbs and in blastemas of the newt Pleurodeles waltl. Using CRISPR/Cas9 to knockout Hoxa13, they reveal that Hoxa13 is required for digit formation with the mutants having fewer digits than wild type. Hoxc13 and Hoxd13, however, seem to be dispensable but overexpression of Hoxd13 in Hoxa13 knockout newts partially rescues limb development; Hoxd13 expression in limb buds is strongly dependent on Hoxa13a. Lastly, the authors show that, following the amputation of the forelimbs, Hox13 genes, and Hoxa13 in particular, are also required for digit formation during limb regeneration.
Fig. 2. Movie 1 taken from Takeuchi et al, 2022 showing regeneration of a wild type limb in Pleurodeles waltl.
Both of the articles discussed today concern digit formation in regenerating newt limbs. D.R. Newth showed that regeneration can rescue proper digit development in abnormal limbs and, given that Hox genes have specific positional expression patterns, the work from Takeuchi and colleagues suggests that Hox13 genes might be one of the factors involved in this process. Furthermore, Takeuchi and colleagues demonstrate another parallel, but not identical, link between development and regeneration. Tomorrow, in the final post of this mini-series, we look towards the nervous system.
References
Ben D. Cox, Maximina H. Yun, Kenneth D. Poss; Can laboratory model systems instruct human limb regeneration? Development 15 October 2019; 146 (20): dev181016. doi: https://doi.org/10.1242/dev.181016
Alberto Joven, Ahmed Elewa, András Simon; Model systems for regeneration: salamanders. Development 15 July 2019; 146 (14): dev167700. doi: https://doi.org/10.1242/dev.167700
Caitlin McQueen, Matthew Towers; Establishing the pattern of the vertebrate limb. Development 1 September 2020; 147 (17): dev177956. doi: https://doi.org/10.1242/dev.177956
D. R. Newth; On Regeneration after the Amputation of Abnormal Structures: II. Supernumerary Induced Limbs. Development 1 September 1958; 6 (3): 384–392. doi: https://doi.org/10.1242/dev.6.3.384
Takashi Takeuchi, Haruka Matsubara, Fumina Minamitani, Yukio Satoh, Sayo Tozawa, Tomoki Moriyama, Kohei Maruyama, Ken-ichi T. Suzuki, Shuji Shigenobu, Takeshi Inoue, Koji Tamura, Kiyokazu Agata, Toshinori Hayashi; Newt Hoxa13 has an essential and predominant role in digit formation during development and regeneration. Development 1 March 2022; 149 (5): dev200282. doi: https://doi.org/10.1242/dev.200282
W.A. Vieira, M. Raymond, K. Kelley, M. A. Cherubino, H. Sahin, C. D. McCusker; Integration failure of regenerated limb tissue is associated with incongruencies in positional information in the Mexican axolotl. Front. Cell Dev. Biol. 2023; 11:1152510. doi: https://doi.org/10.3389/fcell.2023.1152510
Hello, I am Tzer Han and I am very happy to introduce myself as a contributor to the ‘New PI Diaries’. I am starting my lab at the Physics Department of UC San Diego, with a broad interest in understanding how symmetry and order emerge in living matter. We are currently focusing on two main directions: (i) using organoids to study tissue mechanics and chart the morphological landscape of multicellular systems; and (ii) using marine invertebrates to study developmental patterning, cilia dynamics and collective behavior. If any of this sounds interesting to you, feel free to check out my website here.
No doubt, becoming a new PI is rewarding but also challenging. Many tasks to be dealt with are unexpected: no one warned you about them and you have exactly one chance to get everything right. One of these tasks, if you are an experimentalist like me, is designing the lab space that suits your experimental needs. Beyond the aspects with which most of us are familiar with, like allocating benches and equipment spaces, there are infrastructures hidden from plain view that matter as much: Where to put the HEPA filter to ensure clean airflow? Should the optical table be on active or passive damping? What architectural precautions to take in case the 75 gallons aquarium tank starts leaking over a long weekend? So, after many rounds of back and forth with the design architects, electrical and mechanical engineers, safety inspectors, I am excited to see the lab design finally taking shape. It will be a few more months before the bidding, contract award, fire marshall approval and actual construction happen. But I will consider this a win for the moment.
What is it like starting your own lab? What exactly needs to be done? In this ‘New PI diaries’ blog series, we will follow the journey of a few new PIs over the course of their first year of starting their research group, giving the Node readers an inside view of the challenges and highlights of being a new PI.
In the coming months, we will be hearing from three upcoming new PIs: Tzer Han Tan, Margot Smit and Elena Camacho. Watch this space!
Tzer Han has just started his lab at University of California, San Diego, looking at how order, symmetry and dynamics emerge in living matter. You can read his first diary entry here.
Margot’s lab at the University of Tübingen, Germany, will be studying plant cell identity and development, in particular, how plants control the timing of fate transitions.
Elena will be starting her group at Centro Andaluz de Biología del Desarrollo in Spain, investigating human embryonic development by leveraging stem cells and mathematics.
Will you be starting your own lab soon? The Node is especially looking for new PIs who will be setting up their lab somewhere outside of Europe and the US. Get in touch if you are interested!
Epimorphic regeneration, the process of replacing lost appendages, is probably the most impressive example of tetrapod vertebrate regeneration. Vertebrate appendages, such as limbs and tails, are composed of many different cell types from different germ layers (including, for example, neurons, skin, muscle, bone and endothelium), which require the complex integration of positional information and patterning to regenerate functional structures (Cox et al., 2019). Amphibians are probably the best-known executors of epimorphic regeneration, with the pink, smiley-faced axolotl intriguing scientists since the 16th century (Joven et al., 2019). Even five centuries later, researchers are continuing to study amphibians to understand epimorphic regeneration. While studies have revealed the importance of blastema and a handful of signalling factors, there’s still a great deal about epimorphic regeneration that remains to be solved. Today, I summarise insights from the following two articles:
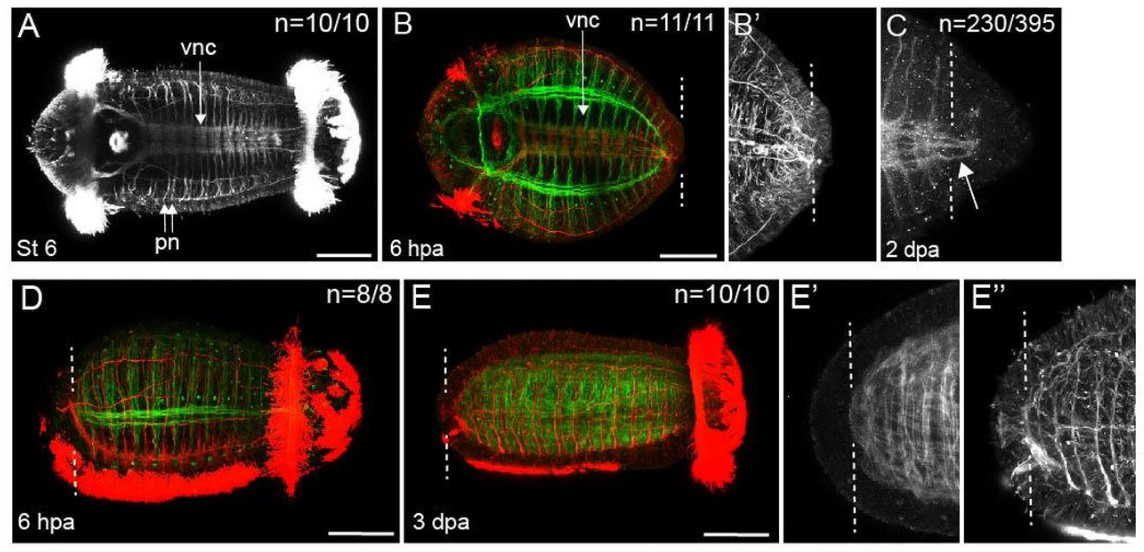
The almost-aptly-amphibian named David Richmond Newth was a Birmingham-born zoologist. After serving in India and Burma during the Second World War, he returned to the UK as a lecturer at Univesity College London. In his 1958 JEEM/Development publication (Newth, 1958), David sought to clarify some previous studies asking whether a regenerated amphibian tail would resemble the original: can animals restore defective appendages and regenerate structures that they didn’t have in the first place? Newth used both axolotl and the frog Rana temporaria as experimental systems, and operated on the embryos to induce the animals to develop an incomplete tail fin. In the frogs, he removed the dorsal half of the tail bud, whereas in axolotl he manipulated posterior neural folds to create ventral fin defects. Next, Newth amputated the tail at different positions along the anterior-posterior axis, cutting anterior to the defect or within the missing tissue. In the case of the Rana tadpoles, David repeated the amputation again (Fig. 1). As long as the amputation occurred anterior to the missing tail tissue, the amphibians could regenerate whole tails but dissection through the missing tissue did not generate complete tail outgrowth from the wound (Fig. 1). By showing that regeneration could form structures that were missing in the original appendage, this work indicates that embryonic development and regeneration might use different mechanisms of growth and patterning. However, the author is careful to point out that, in this case, the mechanism of regeneration in normal animals might not be the same as ‘abnormal’ ones; regeneration of ‘normal’ tissue could still follow developmental mechanisms. Second, this study demonstrates the importance of the position of amputation and indicates that the cells that form the blastema dictate which structures can be formed during regrowth.
Fig. 1. “The regeneration of tails in Rana tadpoles which had earlier lost the dorsal half of their tail-buds. The numbers are of animals surviving to the stages indicated.” Taken from Fig. 3 in Newth (1958).
It seems that the Axolotl is a species that has truly stood the test of time and is just as popular a model today as 500 years ago (Joven et al., 2019). However, technical scientific advances have advanced the toolbox available for asking complex research questions, including the generation of sophisticated axolotl genetic lines and reporters (e.g. Duerr et al., 2022). Although a wide range of frog species have been used in classical studies, present research largely concentrates on Xenopus spp. (Phipps et al., 2020). Whether regeneration employs the same development mechanisms to produce tissues during development remains unclear. Similar developmental signalling pathways (e.g. FGF, Shh) are involved in regenerating appendages; however, single-cell technologies are revealing that the progenitors deriving from the regeneration-specific organising centre – the blastema – have distinct molecular identities from those in the embryo and follow convergent, but different, trajectories (Čapek and Müller, 2019; Tsai et al., 2019). As we heard for wound healing yesterday, many recent investigations have focused on the roles of immune cells in amphibian tail regeneration, showing that myeloid cells and the factors they secrete are required for a regeneration-permissive environment (e.g. Tsai et al., 2019; Aztekin et al., 2020).
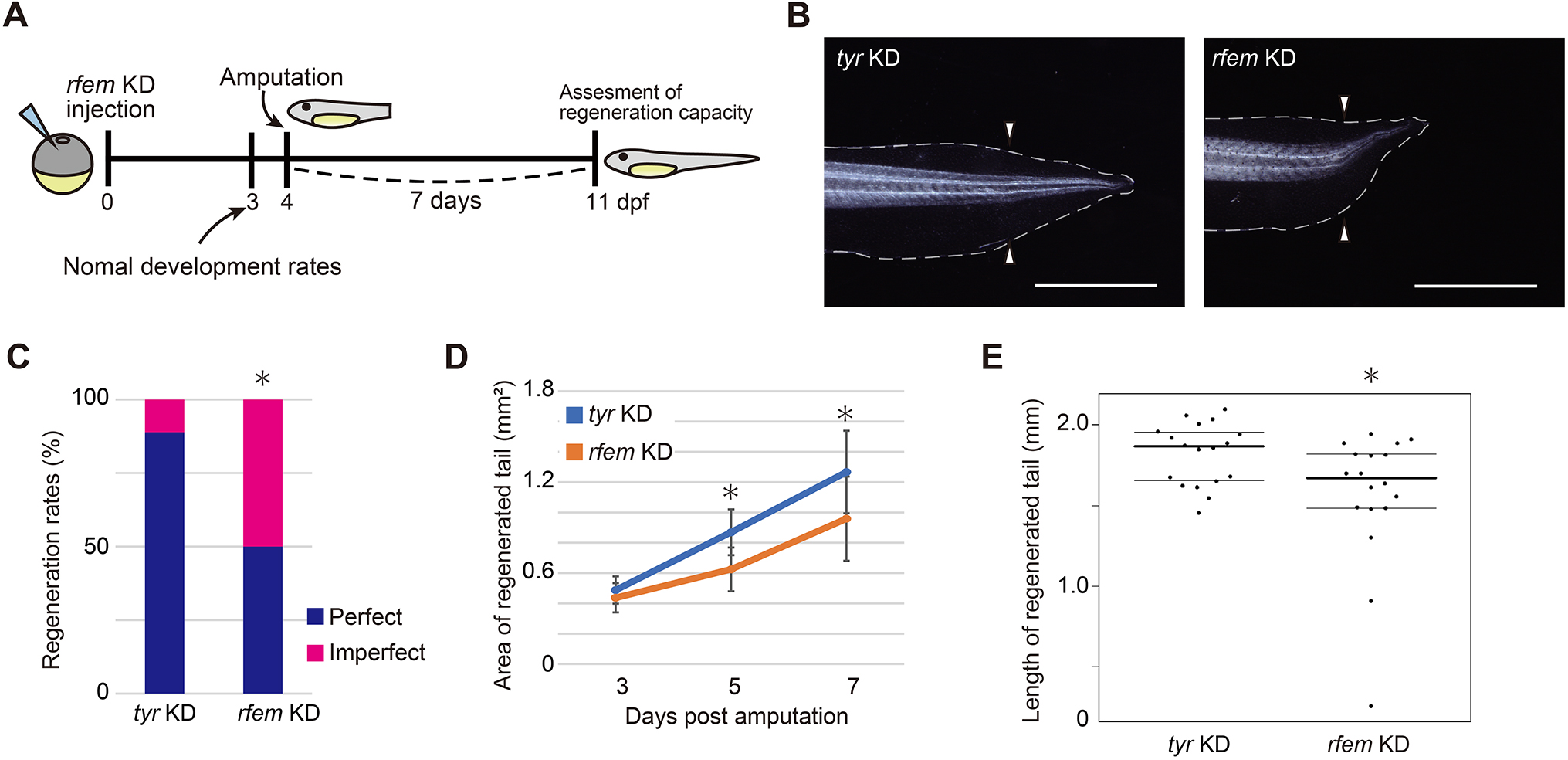
In their article published last month in Development, Deguchi and colleagues continue interrogating the intervention of the immune system in Xenopus laevis tail regeneration (Deguchi et al., 2023). Through a screen to identify cells that support progenitor proliferation, they find that regeneration factors expressed on myeloid (of which there are two paralogues in Xenopus, hereinafter collectively referred to as rfem) increases expression following tail amputation. It’s also required for regeneration because knockdown of rfem generates significantly shorter regenerated tails (Fig. 2). Using RNA sequencing of tail stumps and intact tails, the authors show that rfem-expressing cells cluster with leukocyte markers expressed by macrophages and dendritic cells. Deguchi and colleagues reduce the macrophage population, and thus rfem-expressing cells, by knocking down key monocyte-lineage gene csf1, which also caused tail regeneration abnormalities. Conversely, the addition of rfem-expressing macrophage-like cells in rfem knockdown animals rescues tail regeneration. Although the mechanism of Rfem activity is yet to be elucidated, this study builds upon previous research by identifying a factor through which immune cells might regulate epimorphic regeneration (Aztekin et al., 2020).
Fig. 2. “rfem.L and/or rfem.S are required for normal tail regeneration.” Taken from Fig. 2 in Deguchi et al., 2023 (see publication for full legend).
These two examples from the past and present both ask about the regenerative potential of the amphibian tail. Newth shows that regenerative programs can restore tissues that are effectively formed during development and aren’t possessed by the original structure and indirectly hints about various progenitor populations that might be involved. Deguchi and colleagues take this further to demonstrate that supporting cells, most likely macrophages, have an important role in regulating tail regeneration through the action of rfem-encoded proteins. However, this tale is far from over! Tomorrow, we indulge in a second helping of epimorphic regeneration, focusing on a dexterous but incredibly handy experimental technique: tissue grafts.
References
Can Aztekin, Tom W. Hiscock, Richard Butler, Francisco De Jesús Andino, Jacques Robert, John B. Gurdon, Jerome Jullien; The myeloid lineage is required for the emergence of a regeneration-permissive environment following Xenopus tail amputation. Development 1 February 2020; 147 (3): dev185496. doi: https://doi.org/10.1242/dev.185496
Daniel Čapek, Patrick Müller; Positional information and tissue scaling during development and regeneration. Development 15 December 2019; 146 (24): dev177709. doi: https://doi.org/10.1242/dev.177709
Ben D. Cox, Maximina H. Yun, Kenneth D. Poss; Can laboratory model systems instruct human limb regeneration? Development 15 October 2019; 146 (20): dev181016. doi: https://doi.org/10.1242/dev.181016
Momoko Deguchi, Taro Fukazawa, Takeo Kubo; regeneration factors expressed on myeloid expression in macrophage-like cells is required for tail regeneration in Xenopus laevis tadpoles. Development 1 August 2023; 150 (15): dev200467. doi: https://doi.org/10.1242/dev.200467
Timothy J. Duerr, Eun Kyung Jeon, Kaylee M. Wells, Antonio Villanueva, Ashley W. Seifert, Catherine D. McCusker, James R. Monaghan; A constitutively expressed fluorescent ubiquitination-based cell-cycle indicator (FUCCI) in axolotls for studying tissue regeneration. Development 15 March 2022; 149 (6): dev199637. doi: https://doi.org/10.1242/dev.199637
Alberto Joven, Ahmed Elewa, András Simon; Model systems for regeneration: salamanders. Development 15 July 2019; 146 (14): dev167700. doi: https://doi.org/10.1242/dev.167700
D. R. Newth; On Regeneration after the Amputation of Abnormal Structures: I. Defective Amphibian Tails. Development 1 June 1958; 6 (2): 297–307. doi: https://doi.org/10.1242/dev.6.2.297
Lauren S. Phipps, Lindsey Marshall, Karel Dorey, Enrique Amaya; Model systems for regeneration: Xenopus. Development 15 March 2020; 147 (6): dev180844. doi: https://doi.org/10.1242/dev.180844
Stephanie L. Tsai, Clara Baselga-Garriga, Douglas A. Melton; Blastemal progenitors modulate immune signaling during early limb regeneration. Development 1 January 2019; 146 (1): dev169128. doi: https://doi.org/10.1242/dev.169128
Carlo Donato Caiaffa, Yogeshwari S. Ambekar, Manmohan Singh, Ying Linda Lin, Bogdan Wlodarczyk, Salavat R. Aglyamov, Giuliano Scarcelli, Kirill V. Larin, Richard Finnell
Andrew Liu, Jessica O’Connell, Farley Wall, Richard W. Carthew
Dynamics of Ds and Fat protein distributions across the Drosophila wing pouch from Liu et al. This image is made available under a CC-BY-NC 4.0 International license.
Eduardo D. Gigante, Katarzyna M. Piekarz, Alexandra Gurgis, Leslie Cohen, Florian Razy-Krajka, Sydney Popsuj, Hussan S. Ali, Shruthi Mohana Sundaram, Alberto Stolfi
Cell division and differentiation in C. robusta embryos from Gigante et al. This image is made available under a CC-BY-NC 4.0 International license.
Marcia Mesquita Peixoto, Francisca Soares-da-Silva, Valentin Bonnet, Gustave Ronteix, Rita Faria Santos, Marie-Pierre Mailhe, Xing Feng, João Pedro Pereira, Emanuele Azzoni, Giorgio Anselmi, Marella de Bruijn, Charles N. Baroud, Perpétua Pinto-do-Ó, Ana Cumano
Juan Manuel Gomez, Hendrik Nolte, Elisabeth Vogelsang, Bipasha Dey, Michiko Takeda, Girolamo Giudice, Miriam Faxel, Alina Cepraga, Robert Patrick Zinzen, Marcus Krüger, Evangelia Petsalaki, Yu-Chiun Wang, Maria Leptin
Yogeshwari S. Ambekar, Carlo Donato Caiaffa, Bogdan Wlodarczyk, Manmohan Singh, Alexander W. Schill, John Steele, Salavat R. Aglyamov, Giuliano Scarcelli, Richard H. Finnell, Kirill V. Larin
Akanksha Jain, Gilles Gut, Fátima Sanchís Calleja, Ryoko Okamoto, Simon Streib, Zhisong He, Fides Zenk, Malgorzata Santel, Makiko Seimiya, René Holtackers, Sophie Martina Johanna Jansen, J. Gray Camp, Barbara Treutlein
Cora Demler, John Coates Lawlor, Ronit Yelin, Dhana Llivichuzcha-Loja, Lihi Shaulov, David Kim, Megan Stewart, Frank Lee, Thomas Schultheiss, Natasza Kurpios
Simon Schneider, Andjela Kovacevic, Michelle Mayer, Ann-Kristin Dicke, Lena Arévalo, Sophie A. Koser, Jan N. Hansen, Samuel Young, Christoph Brenker, Sabine Kliesch, Dagmar Wachten, Gregor Kirfel, Timo Strünker, Frank Tüttelmann, Hubert Schorle
Paula Cubillos, Nora Ditzer, Annika Kolodziejczyk, Gustav Schwenk, Janine Hoffmann, Theresa M. Schütze, Razvan P. Derihaci, Cahit Birdir, Johannes E. M. Köllner, Andreas Petzold, Mihail Sarov, Ulrich Martin, Katherine R. Long, Pauline Wimberger, Mareike Albert
A. Lardenois, A. Suglia, CL. Moore, B. Evrard, L. Noël, P. Rivaud, A. Besson, M. Toupin, S. Léonard, L. Lesné, I. Coiffec, S. Nef, V. Lavoué, O. Collin, A. Chédotal, S. Mazaud-Guittot, F. Chalmel, AD. Rolland
Joab Camarena, Sofia E. Luna, Jessica P. Hampton, Kiran R. Majeti, Carsten T. Charlesworth, Eric Soupene, Vivien A. Sheehan, M. Kyle Cromer, Matthew H. Porteus
Michał Brouze, Agnieszka Czarnocka-Cieciura, Olga Gewartowska, Monika Kusio-Kobiałka, Kamil Jachacy, Marcin Szpila, Bartosz Tarkowski, Jakub Gruchota, Paweł Krawczyk, Seweryn Mroczek, Ewa Borsuk, Andrzej Dziembowski
Prakhar Bansal, Erin C Banda, Heather R Glatt-Deeley, Christopher Stoddard, Jeremy W Linsley, Neha Arora, Darcy T Ahern, Yuvabharath Kondaveeti, Michael Nicouleau, Miguel Sabariego-Navarro, Mara Dierssen, Steve Finkbeiner, Stefan F Pinter
Jos Smits, Dulce Lima Cunha, Jieqiong Qu, Nicholas Owen, Lorenz Latta, Nora Szentmary, Berthold Seitz, Lauriane N. Roux, Mariya Moosajee, Daniel Aberdam, Simon J. van Heeringen, Huiqing Zhou
Lucas G. A. Ferreira, Marina M. L. Kizys, Gabriel A. C. Gama, Svenja Pachernegg, Gorjana Robevska, Andrew H. Sinclair, Katie L. Ayers, Magnus R. Dias da Silva
Temperature control and simultaneous monitoring of transcriptional activity from Martini and Garcia. This image is made available under a CC-BY-ND 4.0 International license.
Prakhar Bansal, Erin C Banda, Heather R Glatt-Deeley, Christopher Stoddard, Jeremy W Linsley, Neha Arora, Darcy T Ahern, Yuvabharath Kondaveeti, Michael Nicouleau, Miguel Sabariego-Navarro, Mara Dierssen, Steve Finkbeiner, Stefan F Pinter
Alexa Giovannini, Sabrina Piechota, Maria Marchante, Kathryn S Potts, Graham Rockwell, Bruna Paulsen, Alexander D Noblett, Samantha L Estevez, Alexandra B Figueroa, Caroline Aschenberger, Dawn A Kelk, Marcy Forti, Shelby Marcinyshyn, Ferran Barrachina, Klaus Wiemer, Marta Sanchez, Pedro Belchin, Merrick Pierson Smela, Patrick R.J. Fortuna, Pranam Chatterjee, David H McCulloh, Alan Copperman, Daniel Ordonez-Perez, Joshua U Klein, Christian C Kramme
Wai Kit Chan, Danilo Negro, Victoria M Munro, Helen Marshall, Zrinko Kozić, Megan Brown, Mariana Beltran, Neil C Henderson, David J Price, John O Mason
Mercedes Barrero, Anna V. López-Rubio, Aleksey Lazarenkov, Enrique Blanco, Moritz Bauer, Luis G. Palma, Anna Bigas, Luciano Di Croce, José Luis Sardina, Bernhard Payer
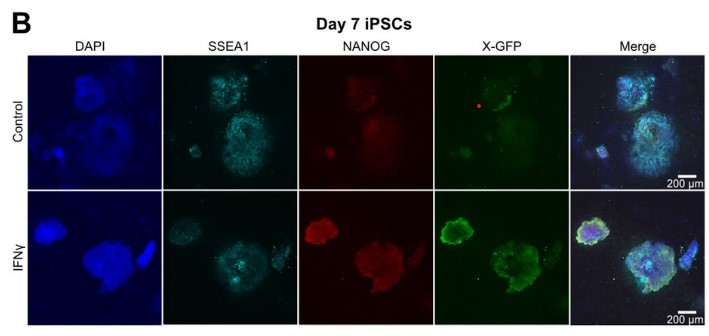
iPSC colonies upon early interferon γ treatment from Barrero et al. This image is made available under a CC-BY-NC 4.0 International license.
Tammy T Nguyen, Zinger Yang Loureiro, Anand Desai, Tiffany DeSouza, Shannon Joyce, Lyne Khair, Amruta Samant, Haley Cirka, Javier Solivan-Rivera, Rachel Ziegler, Michael Brehm, Louis M Messina, Silvia Corvera
Xiaoyu Yang, Ian R. Jones, Poshen B. Chen, Han Yang, Xingjie Ren, Lina Zheng, Bin Li, Yang E. Li, Quan Sun, Jia Wen, Cooper Beaman, Xiekui Cui, Yun Li, Wei Wang, Ming Hu, Bing Ren, Yin Shen
Diana R. Rodríguez-García, Yossmayer del Carmen Rondón Guerrero, Lucía Ferrero, Andrés Hugo Rossi, Esteban A. Miglietta, Ariel A. Aptekmann, Eliana Marzol, Javier Martínez Pacheco, Mariana Carignani, Victoria Berdion Gabarain, Leonel E. Lopez, Gabriela Díaz Dominguez, Cecilia Borassi, José Juan Sánchez-Serrano, Lin Xu, Alejandro D. Nadra, Enrique Rojo, Federico Ariel, José M. Estevez
Craig L. Cowling, Arielle L. Homayouni, Jodi B. Callwood, Maxwell R. McReynolds, Jasper Khor, Haiyan Ke, Melissa A. Draves, Katayoon Dehesh, Justin W. Walley, Lucia C. Strader, Dior R. Kelley
Matēj Drs, Samuel Haluška, Eliška Škrabálková, Pavel Krupař, Andrea Potocká, Lucie Brejšková, Karel Muller, Natalia Serrano, Aline Voxeur, Samantha Vernhettes, Jitka Ortmannová, George Caldarescu, Matyas Fendrych, Martin Potocký, Viktor Žárský, Tamara Pečenková
Sebastjen Schoenaers, Hyun Kyung Lee, Martine Gonneau, Elvina Faucher, Thomas Levasseur, Elodie Akary, Naomi Claeijs, Steven Moussu, Caroline Broyart, Daria Balcerowicz, Hamada AbdElgawad, Andrea Bassi, Daniel Santa Cruz Damineli, Alex Costa, Jose A Feijo, Celine Moreau, Estelle Bonnin, Bernard Cathala, Julia Santiago, Herman Hofte, Kris Vissenberg
Simon Amiard, Léa Feit, Lauriane Simon, Samuel Le Goff, Loriane Loizeau, Léa Wolff, Falk Butter, Clara Bourbousse, Fredy Barneche, Christophe Tatout, Aline. V. Probst
Linkan Dash, Maxwell R. McReynolds, Melissa A. Draves, Rajdeep S. Khangura, Rebekah L. Muench, Jasper Khor, Jodi B. Callwood, Craig L. Cowling, Ludvin Mejia, Michelle G. Lang, Brian P. Dilkes, Justin W. Walley, Dior R. Kelley
Avilash S. Yadav, Lilan Hong, Patrick M. Klees, Annamaria Kiss, Xi He, Iselle M. Barrios, Michelle Heeney, Anabella Maria D. Galang, Richard S. Smith, Arezki Boudaoud, Adrienne H.K. Roeder
David Bolumar, Javier Moncayo-Arlandi, Javier Gonzalez-Fernandez, Ana Ochando, Inmaculada Moreno, Ana Monteagudo-Sanchez, Carlos Marin, Antonio Diez, Paula Fabra, Miguel Ángel Checa, Juan José Espinos, David K. Gardner, Carlos Simon, Felipe Vilella
Isabel Tundidor, Marta Seijo-Vila, Sandra Blasco-Benito, María Rubert-Hernández, Gema Moreno-Bueno, Laura Bindila, Rubén Fernández de la Rosa, Manuel Guzmán, Cristina Sánchez, Eduardo Pérez-Gómez
Khursheed Iqbal, Brandon Nixon, Benjamin Crnkovich, Esteban M. Dominguez, Ayelen Moreno-Irusta, Regan L. Scott, Ha T.H. Vu, Geetu Tuteja, Jay L. Vivian, Michael J. Soares
Jesús A. Andrés-San Román, Carmen Gordillo-Vázquez, Daniel Franco-Barranco, Laura Morato, Cecilia H. Fernández-Espartero, Gabriel Baonza, Antonio Tagua, Pablo Vicente-Munuera, Ana M. Palacios, Maria P. Gavilan, Fernando Martín-Belmonte, Valentina Annese, Pedro Gómez-Gálvez, Ignacio Arganda-Carreras, Luis M. Escudero
Nitesh Kumar Sharma, Ram Ayyala, Dhrithi Deshpande, Yesha M Patel, Viorel Munteanu, Dumitru Ciorba, Andrada Fiscutean, Mohammad Vahed, Aditya Sarkar, Ruiwei Guo, Andrew Moore, Nicholas Darci-Maher, Nicole A Nogoy, Malak S. Abedalthagafi, Serghei Mangul
Wound healing is a crucial process in both regenerating and non-regenerating tissues. In addition to restoring the biological function of a tissue by removing damaged cells and reconstituting a continuous epithelium, the response to a wound can determine whether a pro-regenerative environment is provided (e.g. a blastema) or, alternatively, induce the formation of a fibrotic (and sometimes pathogenic) scar through the deposition of extracellular matrix (Wong and Whited, 2020). In this third episode, I discuss the highlights from the following two articles:
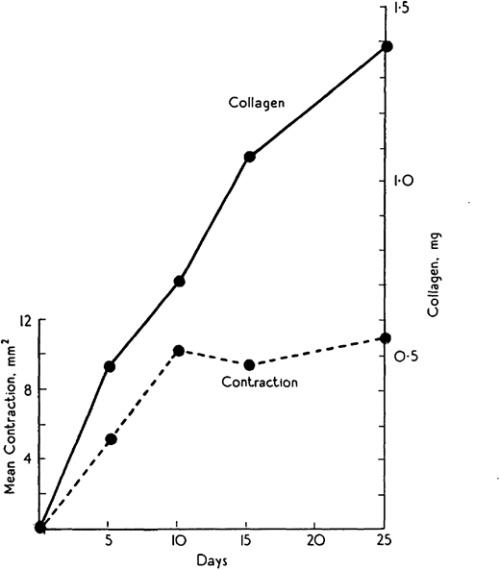
Michael Abercrombie was a prominent English cell biologist who’s credited with early science public engagement by writing biology books for The People with popular publisher, Penguin. Michael’s co-authors for this paper are somewhat more elusive, but together, Abercrombie, Flint and James seemed to have a penchant for tattooing rodents. Their 1956 JEEM/Development publication follows an earlier paper from the same authors, published in the same journal, describing the purse string-like contraction of rat skin following wounding and the increase of extracellular matrix protein, collagen, in scabs during the healing process (Fig. 1; Abercrombie et al., 1954). In their follow-up piece, at a time when one could publish a paper containing no figures or tables whatsoever(!), Abercrombie and colleagues turn their tattoo needles towards male guinea pigs (although I’m unsure why rats were abandoned) to further characterise the role of collagen. Abercrombie, Flint and James set up an experiment in which their guinea pigs were fed a vitamin C-deficient diet, based on the knowledge that vitamin C is needed for collagen synthesis (Wolbach and Bessey, 1942), with a control group provided with a vitamin C supplement. After injuring the skin of the guineas and using tattoos to trace skin deformation as it healed, they showed that, while wound contraction was unaffected, vitamin C-deficient animals had smaller, fragile scabs that contained less collagen as determined by measuring hydroxyproline as a proxy (Abercrombie et al., 1956). These results were able to uncouple the possible roles of collagen in wound closure versus scarring, and led the way for future research to focus on a cell-based – rather than matrix-based – mechanism of epidermal contraction.
Fig. 1. “Graph showing change of collagen content (continuous line) compared with diminution of wounded area (broken line) with time after initial operation.” Taken from (Abercrombie et al., 1954).
Almost 70 years later, collagen is still an important focus in the regeneration community. The wound-healing process of many other organs, in addition to the skin, has been studied. The number of experiments using guinea pigs dropped considerably, but are starting to reemerge, particularly in the field of endometrium regeneration. Instead, the lab staples, mice and Drosophila, are a common mammalian model for, well, just about everything and the relatively young lab species, the zebrafish, has gained prominence, due to its ability to regenerate the heart (Simões and Riley, 2022) and spinal cord (Becker and Becker, 2022). Focus has turned less towards the exact role of collagen and more towards the cells that secrete it and, importantly, whether this secretion can be regulated in a way that promotes regeneration and prevents scarring with important therapeutic implications. Among these recent studies, the process of inflammation and the role of immune cells during wound healing and regeneration has gained attention (Ginhoux and Martin, 2022). However, tattooing, it seems, has been lost from many a materials and methods section.
Macrophages directly contribute collagen to scar formation during zebrafish heart regeneration and mouse heart repair Filipa C. Simões, Thomas J. Cahill, Amy Kenyon, Daria Gavriouchkina, Joaquim M. Vieira, Xin Sun, Daniela Pezzolla, Christophe Ravaud, Eva Masmanian, Michael Weinberger, Sarah Mayes, Madeleine E. Lemieux, Damien N. Barnette, Mala Gunadasa-Rohling, Ruth M. Williams, David R. Greaves, Le A. Trinh, Scott E. Fraser, Sarah L. Dallas, Robin P. Choudhury, Tatjana Sauka-Spengler and Paul R. Riley
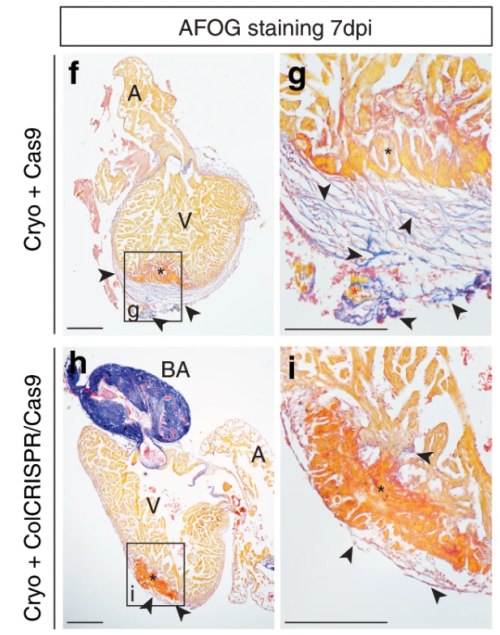
Simões (University of Oxford) and colleagues’ paper, published in Nature Communications in 2020, challenges previous assumptions in the field that immune cells only stimulate fibroblasts to deposit collagen during cardiac wound healing. It’s chock-full of figures and data – in stark contrast to Abercrombie and pals – and they use a vast array of sophisticated genetic techniques, including transgenic transplants and cell-specific knockouts in both neonatal (regenerative) and adult mice, as well as zebrafish, along with imaging and next-generation sequencing (Simões et al., 2020). The authors perform transcriptome analyses of macrophages to show that they have distinct responses to wounds depending on the type (e.g. resection vs. cryoinjury in zebrafish) and the age of the animal (e.g. neonate vs. adult mice). In addition, in the unregenerative adult mouse and zebrafish cryoinjury conditions, macrophages express collagen. Using fluorescently tagged collagen constructs, the authors show that adult macrophages directly contribute collagen to the scar. Supporting this, the transplant of adult macrophages into neonates induces ectopic scarring during cardiac healing. Importantly, the genetic knockdown of collagen genes in zebrafish macrophages is sufficient to significantly reduce scar formation in the heart following cryoinjury, potentially providing a permissive environment for cardiac regeneration (Fig. 2). This work from Simões and colleagues posit macrophages as a direct source of collagen during scarring and provides a future avenue for therapeutic treatment of heart damage.
Fig. 2. “AFOG staining of representative images showing healthy myocardium (yellow), injured myocardium (orange) and collagen (blue). Excess scar tissue (arrowheads) at the periphery of the cryoinjured area (black asterisk) is seen in hearts transplanted with 6 days post fertilization (dpf) GFP+ macrophages injected with Cas9 only (f, with high-magnification inset in g).” Taken from Simões et al., 2020.
Both these papers are interested in understanding collagen during wound healing. Abercrombie, Flint and James, determine that collagen is dispensable for skin wound contraction, but not for scab formation, while Simões and colleagues reveal macrophages as an unlikely source for collagen deposition in the mammalian heart. Tomorrow, we turn our heads (or tails) towards an excellent demonstration of epimorphic regeneration.
References
M. Abercrombie, M. H. Flint, D. W. James; Collagen Formation and Wound Contraction during Repair of Small Excised Wounds in the Skin of Rats. Development 1 September 1954; 2 (3): 264–274. doi: https://doi.org/10.1242/dev.2.3.264
Thomas Becker, Catherina G. Becker; Regenerative neurogenesis: the integration of developmental, physiological and immune signals. Development 15 April 2022; 149 (8): dev199907. doi: https://doi.org/10.1242/dev.199907
Florent Ginhoux, Paul Martin; Insights into the role of immune cells in development and regeneration. Development 15 April 2022; 149 (8): dev200829. doi: https://doi.org/10.1242/dev.200829
Filipa C. Simões, T. J. Cahill, A. Kenyon et al. Macrophages directly contribute collagen to scar formation during zebrafish heart regeneration and mouse heart repair. Nat Commun 2020; 11, 600. https://doi.org/10.1038/s41467-019-14263-2
Filipa C. Simões, Paul R. Riley; Immune cells in cardiac repair and regeneration. Development 15 April 2022; 149 (8): dev199906. doi: https://doi.org/10.1242/dev.199906
Alan Y. Wong, Jessica L. Whited; Parallels between wound healing, epimorphic regeneration and solid tumors. Development 1 January 2020; 147 (1): dev181636. doi: https://doi.org/10.1242/dev.181636
We are delighted to announce that we are offering an opportunity for a 3-month internship to work on the three community sites – the Node, preLights and FocalPlane. This is being offered as a Professional Internships for PhD Students (PIPS) placement to students on the BBSRC DTP program in the UK, or on similar programs where an internship forms part of the PhD training.
If you have a passion for science communication and writing, as well as a love for all things biology, this could be the perfect internship for you! You’ll get an insight into what it’s like to work in the online scicomm environment, and you’ll have the opportunity to come up with ideas for content, talk to potential authors about writing for us, help community site users with their posts, and run the relevant social media accounts. As this internship is based in a publishing company, you’ll also learn about how science publishing works from the inside.
Unlike many other mammalian tissues, adult skeletal muscle has a remarkable aptitude for regeneration. Even after severe and repeated damage, functional skeletal muscle can be regenerated within a month.Such robust skeletal muscle restoration has been attributed to a population of Pax7+ stem/progenitor cells named satellite cells, which proliferate in response to injury, differentiate and fuse to generate mature muscle fibres – the mature, multinucleated, functional cells of the muscle (Collins and Kardon, 2021). In this second instalment, I discuss the highlights from the following two articles:
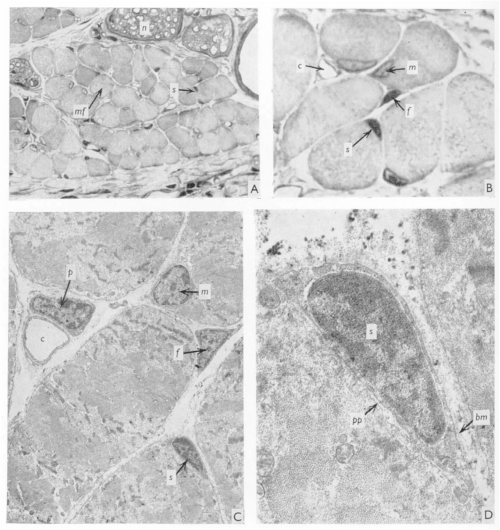
J. C. T. Church spent most – if not all – of his career in East Africa, with some time at Makerere University College Medical School, Uganda, and then at University College in Nairobi, Kenya, where he worked as an orthopaedic surgeon. When not in the hospital, the “enthusiastic teacher” saw it fit to poke holes in the local population of fruit bats (Church and Noronha, 1965). In 1970, JEEM/Development published J.’s latest revelations from the fruit bat Eidolon helvum regarding the ability of skeletal muscle to regenerate following injury. Back then, satellite cells were well known but it remained to be determined whether they were indeed the source of restoring muscle fibre nuclei (myonuclei) during regeneration. Muscle regeneration had also largely been descriptive and, in his publication, Church provided perhaps the first quantitative report of cell populations in the skeletal muscle using electron-microscopy (EM; Fig. 1). Church’s approach was to perform single or repeated muscle ‘crush lesion’ experiments, followed by light and EM imaging, to identify and count the ratio of satellite cells to myonuclei. Although the number of muscle fibres remained stable, Church observed increased satellite cells, fibroblasts and myonuclei during regeneration following both single and double lesions. Overall, this study suggested that satellite cells could provide myonuclei to restore muscle fibres and, importantly, self-renew to sustain the population and regenerative response – a key behaviour of stem/progenitor cells.
Fig. 1. “(A) A light photomicrograph of normal bat web muscle, in transverse section, showing numbers of muscle fibres (mf), nerve fascicles (n), and a satellite cell (S), hardly distinguishable at this magnification, ×450. (B) A light photomicrograph, magnified from Fig. 1A, showing the satellite cell (S), an endomycial fibroblast (f), a myonucleus (m) and a capillary (c). × 1700. (C) An electron photomicrograph of the cells in Fig. 1B, showing the satellite cell (S), fibroblast (f), myonucleus (m) and a pericyte (p) adjacent to the capillary (c). × 4500. (D) An electron photomicrograph, enlarged from Fig. 1C, showing the satellite cell (S) lying under the muscle fibre basement membrane (bm), with its inner plasma membrane lying adjacent to that of the syncytium (pp). × 21500.” Taken from Fig. 1 of Church, 1970.
Let us now leap forward 50 years or so. Fortunately for chiropterophiles, fruit bats are now an uncommon model system for skeletal muscle regeneration research (and regenerative biology in general), with mice and zebrafish a more popular choice instead. It took the invention of inducible genetic labelling and ablation of satellite cells in the 2000s to finally confirm that satellite cells are necessary and sufficient for muscle regeneration (Collins and Kardon, 2021). EM has been complemented by state-of-the-art imaging of genetically manipulated (e.g. transgenic reporter) animals and it’s become increasingly evident that other cell populations, in addition to satellite cells, are needed for regeneration. Whether satellite cells contribute to muscle homeostasis, however, has been far less clear.
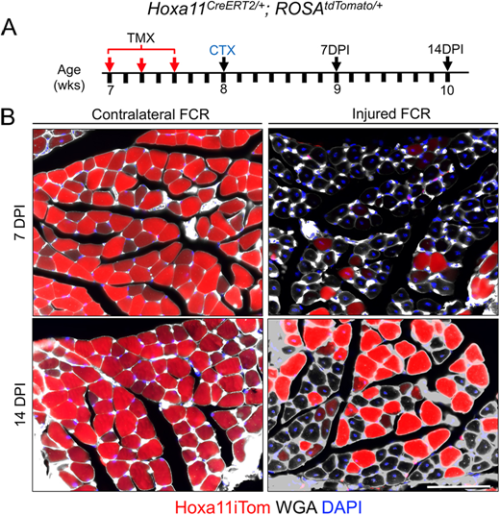
Flynn and colleagues’ study (University of Wisconsin School of Medicine and Public Health), published in Development earlier this year (Flynn et al., 2023), comes at a time of some conflicting evidence in the field when the ablation of satellite cells in adult muscle may or may not lead to muscle degeneration – even in the absence of injury. Evidence that muscle homeostasis is preserved when satellite cells are lost suggests that some other stem/progenitor population can contribute myonuclei for tissue maintenance. Using immunofluorescence and genetic labelling, Flynn and colleagues show that a population of Hoxa11-expressing interstitial stromal cells directly contribute to muscle fibres postnatally. Through genetic lineage-tracing experiments, the team confirm that this Hoxa11-expressing population doesn’t go through a satellite cell-like state (i.e. they are not Pax7+) and, therefore, they are an independent progenitor population. In response to injury, Hoxa11-lineage cells minimally contribute to muscle generation (Fig. 2), contributing to the growing body of evidence of satellite cell-independent muscle formation.
Fig. 2. “Hox11 lineage makes minimal contribution to regenerating myofibers after injury.” Taken from Fig. 9. in Corey et al., 2023.
These articles illustrate a joint interest in understanding the cells that give rise to functional muscle. While J. C. T. Church contributed to the knowledge of satellite cells as a stem/progenitor population in regenerating muscle, the search for muscle-generating cell populations still continues many (many) years later, with Flynn and colleagues identifying a whole new population of Hoxa11-expressing cells that maintain muscle homeostasis. Join me again tomorrow for another trip down memory lane, when we look at early studies of wound healing and the important role of the extracellular matrix.
References
J. C. T. Church, R. F. Noronha. The use of the fruit bat in surgical research. East Afr Med J. July 1965; 42 (7): 348-55.
J. C. T. Church; Cell populations in skeletal muscle after regeneration. Development 1 April 1970; 23 (2): 531–537. doi: https://doi.org/10.1242/dev.23.2.531
Brittany C. Collins, Gabrielle Kardon; It takes all kinds: heterogeneity among satellite cells and fibro-adipogenic progenitors during skeletal muscle regeneration. Development 1 November 2021; 148 (21): dev199861. doi: https://doi.org/10.1242/dev.199861
Corey G. K. Flynn, Paul R. Van Ginkel, Katharine A. Hubert, Qingyuan Guo, Steven M. Hrycaj, Aubrey E. McDermott, Angelo Madruga, Anna P. Miller, Deneen M. Wellik; Hox11-expressing interstitial cells contribute to adult skeletal muscle at homeostasis. Development 15 February 2023; 150 (4): dev201026. doi: https://doi.org/10.1242/dev.201026
Do you feel as if a novel imaging approach would give you new insights into your sample or provide a new way to answer your research questions? Imagine you could take your research question to any institute and work with the experts to unlock the power of imaging technologies and get new data and insights. Well, you can – with Euro-BioImaging.
What is Euro-BioImaging?
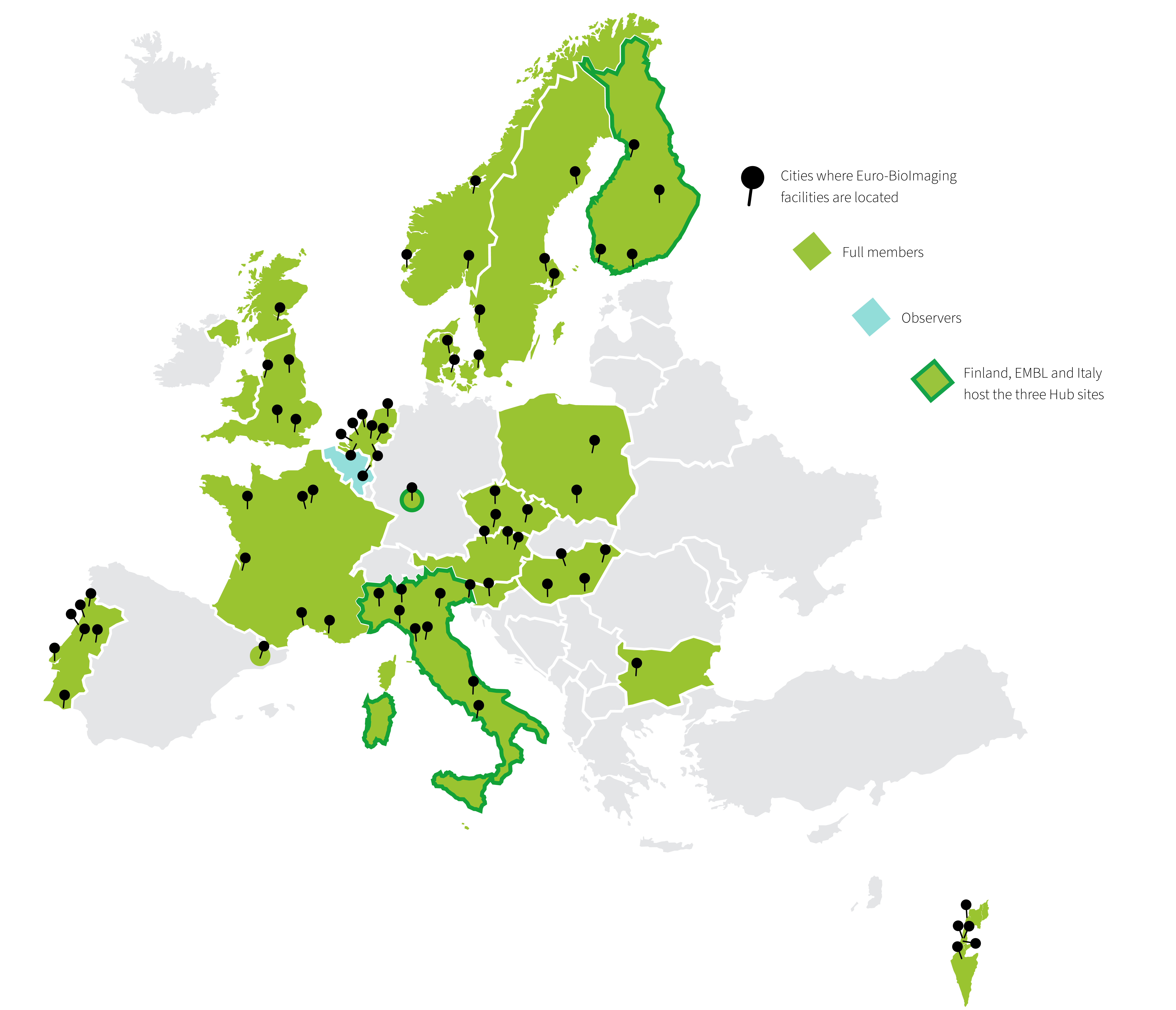
Euro-BioImaging is the European research infrastructure for biological and biomedical imaging that functions as the gateway to over 170 world-class imaging facilities across Europe. Through Euro-BioImaging any researcher, from anywhere around the world, can get access to imaging technologies, image data services and training.
It builds on a set of already existing national and international facilities of excellence in imaging technologies, the Euro-BioImaging Nodes, which provide physical or remote access to imaging technologies, deliver training and support the users at all the stages of their research projects.
Together, over 500 core facility staff work at Euro-BioImaging Nodes and support researchers. We are a fabulous resource for researchers across scale, from atoms to humans, and in diverse disciplines in the life sciences.
Figure 1: Map of Euro-BioImaging member countries, facilities and Hub sites.
How does Euro-BioImaging support researchers?
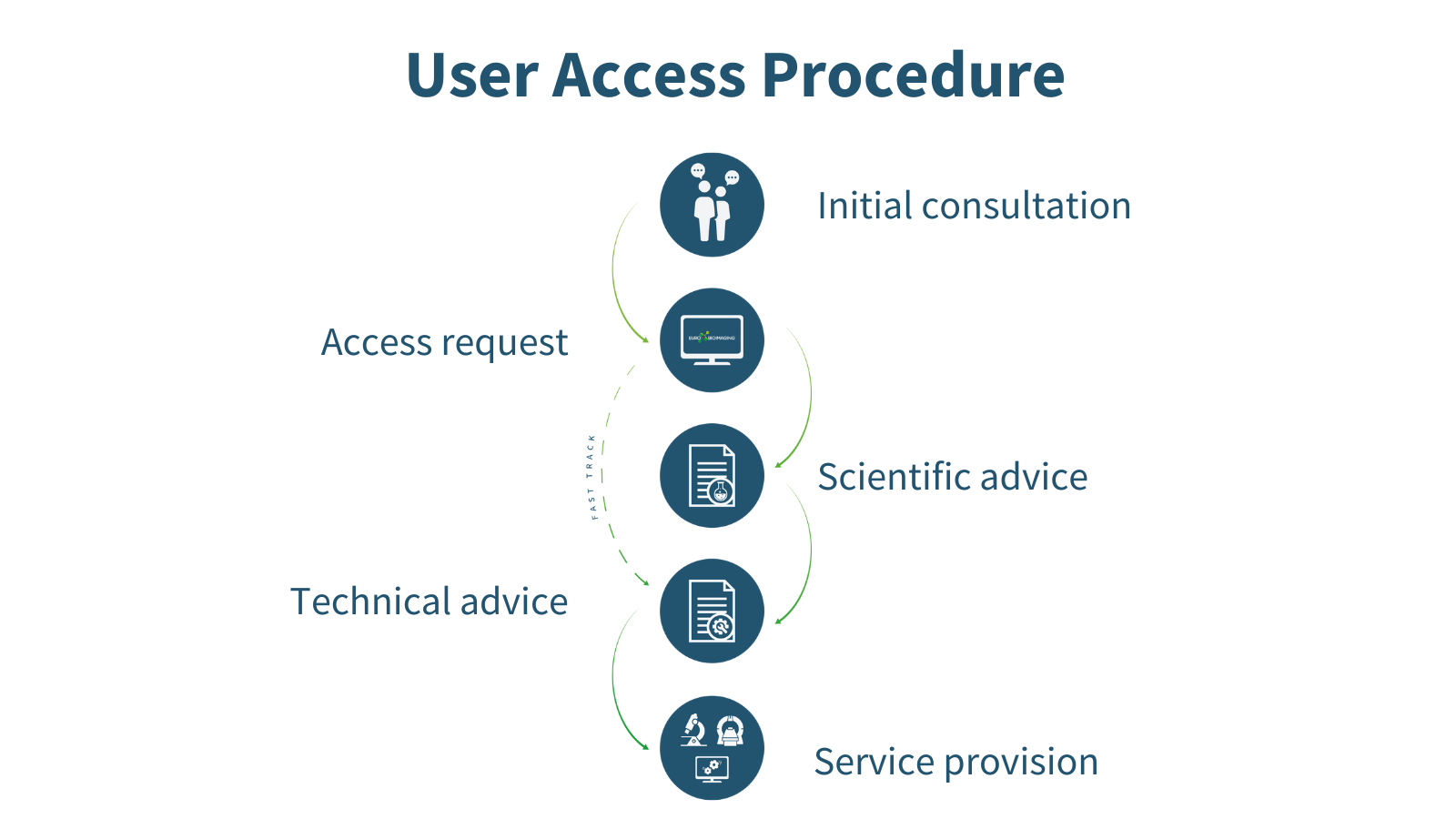
Every researcher, independent of research area, level of expertise, and geographical location, can apply for Euro-BioImaging services whenever they have a project requiring imaging technologies or expertise which they do not have ready access to at their home institute. The expert staff at the Euro-BioImaging Hub can help future users choose the right technology and facility for their research question in the first step of the User Access procedure. Applying for user access is a highly collaborative process in which a researcher has multiple opportunities to hone their experiment and get scientific and technical input from reviewers and technical experts at the imaging facilities before the application is accepted and service provision begins.
Figure 2: The Euro-BioImaging User Access procedure.
An accepted Euro-BioImaging project can be a game-changer. It democratizes access to high-end imaging technologies to push a research question and publication to the next level, and is a starting block towards acquiring new skills, expertise and scientific insight. Euro-BioImaging Users benefit from the expertise of the imaging scientists at the Euro-BioImaging facilities when it comes to sample preparation, experimental set-up and data analysis. Depending on the scope of the project and selected technology, users may also learn skills that they can take back to their home institute.
These skills open doors, especially for early career researchers. Being selected for Euro-BioImaging user access is also a good endorsement of the underlying scientific question or application. Undertaking a project at a Euro-BioImaging facility proves a researcher’s ability to plan and carry out an experiment from start to end.
What imaging technologies are available?
Through the large number of facilities, Euro-BioImaging can offer access to the full range of imaging technologies in the biological and biomedical imaging field. Our technology portfolio covers everything from the nano- to the tissue- and organism scale. We are constantly adding new technologies, making sure that the latest cutting-edge imaging technologies, such as MINFLUX and spatial transcriptomics, are available in open access to all researchers.
Figure 3: The Euro-BioImaging technology portfolio
Harnessing the imaging revolution
The Euro-Bioimaging technology portfolio ranges from light and electron microscopy on the biological imaging side to an expanding range of applications of biomedical imaging, from plant and ex-vivo imaging to animal and human imaging applications.
Electron Microscopy
Our Electron Microscopy portfolio covers cryo-EM techniques for ultrastructural exploration, such as cryo-electron tomography (cryo-ET), as well as the full complement of volume EM techniques, such as FIB-SEM, Array Tomography and Serial Blockface SEM. Many of our facilities also specialise in correlative methods, Correlative X-ray Imaging and EM (CXEM) and correlative light and electron microscopy (CLEM).
Light Microscopy
In light microscopy, our Nodes offer everything from basic confocal microscopy up to single molecule location approaches and intravital imaging. Our light microscopy techniques allow for 3D live cell imaging, tracking, high content screening, and include a variety of functional imaging techniques to explore protein dynamics in living cells. Recently we have added a number of new and highly requested methods, such as MINFLUX, Single Particle tracking and Lattice Lightsheet microscopy to our portfolio.
Model Systems
Euro-BioImaging facilities also offer access and support with a wide range of model systems and how to get the best imaging results out of them, from Drosophila and zebrafish to mouse embryos and organoid systems. Here access to instruments is complemented by technical expertise of facility staff, to support specialized sample handling.
Support Technologies
And of course, we also provide access to adaptive and support technologies, such as laser- based microdissection, Feedback Microscopy, high-speed imaging, microscopes at high biosafety levels, and specialized sample preparation methods, such as Tissue Clearing and Expansion Microscopy.
Euro-BioImaging can also support you if you want to explore the physical and chemical properties of your samples, through access to a range of methods such as MassSpec Imaging, Atomic Force Microscopy, and chemical imaging, such as µ-XRF and μ-PIXE.
Figure 4: Micro-PIXE at the Jožef Stefan Institute, part of our SiMBION Node in Slovenia
How will Euro-BioImaging enhance my research?
So, when you read about a cool, new microscopy method in the literature, you can now allow yourself, not just to imagine, but test the impact that method could have on your research question. If it’s a technology that Euro-BioImaging offers, you can always apply. Because the idea behind Euro-BioImaging is to make the best imaging resources available to all researchers, providing new answers to scientific questions and increasing the impact of research.
What about image data analysis?
Image data analysis is an integral part of any experiment and is therefore usually integrated into the experimental concept at Euro-BioImaging at an early stage. Experts at the Nodes help users extract their data and set up image analysis pipelines, typically preparing for image analysis and data extraction, sometimes even before the actual experiment begins. In addition, Euro-BioImaging offers its users Image Data Analysis (IDA) as a stand-alone service through expert Image Analysts at the Nodes, irrespective of where the image data was acquired.
Users can contact our Nodes when they need:
• Biological and biomedical image data analysis support
• Image registration, segmentation, tracking and more
• Data workflows, bespoke analysis tools and machine learning methods
• Access to high performance computing and specialized software
Before you submit a proposal, feel free to browse our technologies and services, view our user stories to get a feel for what Euro-BioImaging can offer, and reach out to us for help in choosing the right imaging method.
What other opportunities are available?
Training
If you’re not ready to undertake a full experiment with Euro-BioImaging, why not start with a training course? Euro-BioImaging Nodes offer a wide range of training courses – covering the full spectrum of biological and biomedical imaging technologies as well as sample preparation and handling, and image data analysis. Some courses are taught remotely and virtually, increasing their accessibility. Taking a course at a Euro-BioImaging facility is a great way to learn a new skill or improve your technique and to build your network. Here’s an overview of training courses available at Euro-Bioimaging Nodes.
Community building
In addition, Euro-BioImaging organizes regular events focusing on imaging for the benefit of the entire community. You are welcome to join our weekly “Virtual Pub” – a free weekly lecture series, open to all imaging enthusiasts. Topics include new biological and biomedical imaging technologies, image analysis, and other topics of interest.
In addition, we organize a “User Forum” twice a year to highlight the importance of imaging to different research areas. These events feature keynote presentations from prominent scientists as well as presentations from users at our Nodes. We usually record them and make the content available on our YouTube channel.
And finally, we are present at many conferences and community events. So we hope to meet with you and talk face-to-face about the wonderful opportunities Euro-BioImaging can provide. Until we meet in person, you can always reach out to us by email info@eurobiomaging.eu or sign up for our Newsletter to stay informed.
In this latest SciArt profile, we talked to Sonhita Chakraborty about her scientific background in plant biology, how science and art blend into each other, and her first solo art exhibition.
Can you tell us about your background and what you work on now? I’ve worn different hats in the field of plant biology for over 10 years now. After completing my PhD in plant molecular biology, I joined the publishing company Elsevier as a scientific editor. I really enjoy reading and thinking about molecular pathways over a piping hot cup of coffee.
Mitodelic – digital illustration of a mitochondriun. This piece was on the ‘Molecular Cell’ journal cover for their special issue on mitochondria
Were you always going to be a scientist? I didn’t think so. After a trip to Disney World, when I was 10, I would tell everyone who would care to listen that I was going to become an animator for Disney. I think I believed it too. In the last year of my high school I was convinced to pick a more “practical ” career choice and pursue art as a hobby. Out of fear of missing out on opportunities I chose to rendezvous with biology instead of art.
Replication Dream – An artistic and whimsical representation of DNA replication
And what about art – have you always enjoyed it? People often ask me when I first started making art and that’s hard to pinpoint. The happiest memories of my childhood are when I was making art. Some of my early “masterpieces” were “frescos” that I painted as a toddler onto the underside of my parent’s dining table. Stationary stores and art supplies will always delight me.
What or who are your most important artistic influences? I have been spellbound by David Goodsell’s paintings of cells from the get go. On Instagram I’ve come across a cornucopia of very talented and imaginative scientists, artists and science artists who constantly inspire me. I’ve also noticed that feeling grounded and connected with nature can really stoke my creative flame.
Optic Fascination – digital illustration and tribute to the founders of the microscope
How do you make your art? Right now I mostly make art on my iPad but up until 2-3 years ago I used to water colour. I miss the feeling and urgency of pushing wet paint around on a page but digital art let’s me do so much more and explore other artistic aspects of myself.
Big Science – digital collage from publicly available images
Does yourart influence your science at all, or are they separate worlds? That’s a great question! Science and art are definitely not separate in my books and they blend into each other. The amazing science I read about at work gives me a lot of interesting ideas, some of which have become journal cover art. My art doesn’t seem to influence my science (yet). I’ll be starting a postdoc in the fall so I’m not sure what sort of artistic inspirations I will draw from my science (and vice versa) – I can’t wait to find out!
What are you thinking of working on next? I just had my first solo art exhibition at the local library. At work I’m designing a few art covers for different journals. I’m also illustrating for a biology student group at the North Carolina State University. I have an endless list of personal artistic projects I’d like to make time for so I won’t be running out of projects to work on anytime soon.
First exhibition hosted by the Toronto Public Library

 (No Ratings Yet)
(No Ratings Yet)
 (2 votes)
(2 votes)