“Unconventional” posts from #DevMeeting23: Origins of multicellularity (cont’d) & Organoids
Posted by Brent Foster, on 19 September 2023

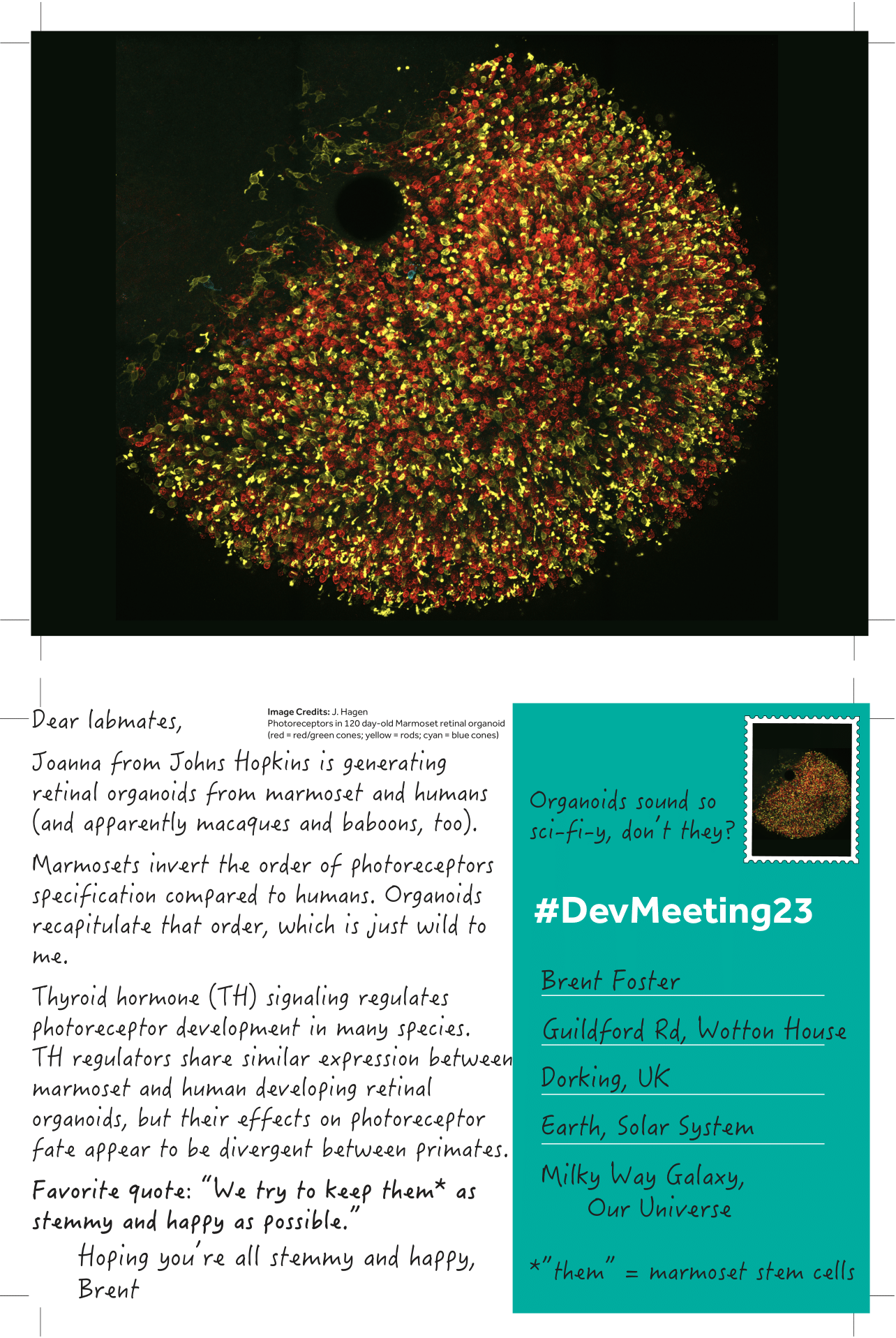
 (1 votes)
(1 votes)Posted by Brent Foster, on 19 September 2023
(1 votes)Posted by Alice Carstairs, on 19 September 2023
Help the NC3Rs fund the best new science that could replace, refine or reduce the use of animals in research.
Our panels are instrumental in supporting the development of new 3Rs models, promoting uptake and championing early career researchers and we are now inviting applications from talented senior researchers to become Panel members from January 2024. For this recruitment round, a wide range of expertise is sought after and applications are particularly welcome from individuals with industrial or interdisciplinary experience. Applications from women, those with a disability, and members of minority ethnic groups are especially encouraged.
For further information, including an application form, visit: https://nc3rs.org.uk/funding-panel-vacancies
The deadline for applications is 4pm (GMT) on Monday 20 November.
If you have any queries, or you would like further information, please email recruit@nc3rs.org.uk.

 (No Ratings Yet)
(No Ratings Yet)Posted by Brent Foster, on 18 September 2023
Introducing “unconventional posts,” a miniseries celebrating the “unconventional” experimental systems presented at #DevMeeting23. Each day I’ll upload handmade postcards spotlighting the breadth of unconventional systems shared in talks or posters and post it on The Node. Here are the first couple to get us started:

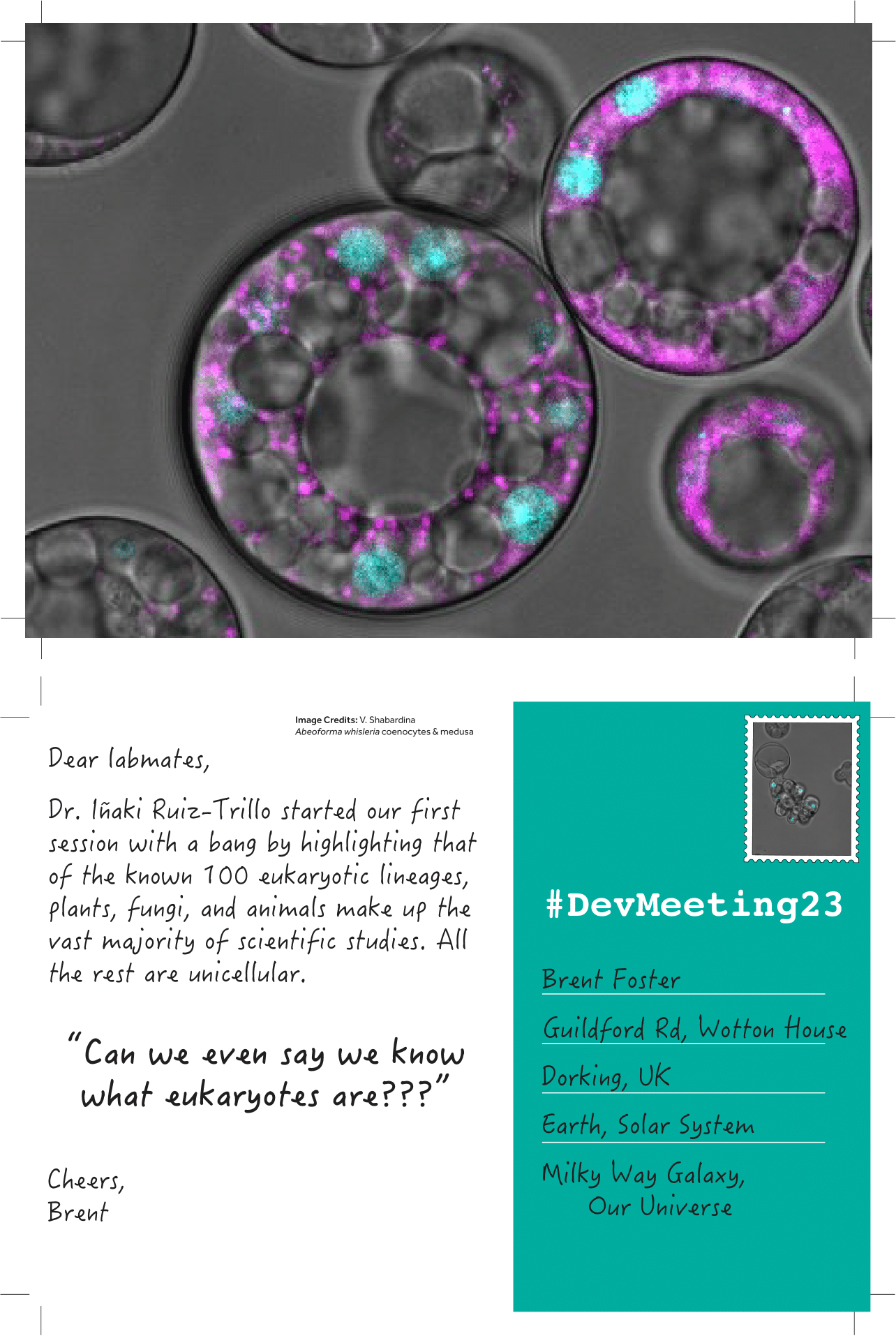
To read the other #DevMeeting23 postcards, visit this page: DevMeeting23 postcards Archives – the Node (biologists.com)
(6 votes)Posted by Dina Mikimoto, on 18 September 2023
When I was a child, I loved the movie “The Fifth Element”—the people working together with living beings from other planets, the space travel while you sleep, the queer clothes, the microwave that makes a fried chicken from a pressed tablet, and the restoration of a living being from a remaining piece. Years later, I am basically working on the last one: creating tissues from cells, building models of organs, and trying to use them to model healthy and diseased conditions. The field I am working in, tissue engineering and regenerative medicine, sounds as futuristic as the film is. And I often feel that people see this field of research in that way— as a science that emerged out of nowhere, in a kind of Frankesteinish and mad scientist way. Talking with people about this area of research and my work, I have heard different reactions, starting from fascination and ending with a question I was asked by one of my friends: “Is it even legal?”. So, I started wondering where this outlandish image was coming from. Is it the lack of awareness of its origin and its goals, or maybe the ethical controversies widely covered by the media, or is it the desire to have simple answers contradicting attempts to share complicated scientific progress?
The first attempts to replace and restore damaged organs
When I started working in this field, I didn’t look too much into history; I just loved the challenge of mixing different subjects and trying to unveil the biological processes by modeling organs outside of the human body. However, talking with people and seeing a lack of trust and disbelief that I was doing something useful or feasible inspired me to investigate the history of how tissue engineering and regenerative medicine came to be. I was struck by how logical its emergence was. The gist of this field is the desire to recreate such biological processes as development and disease outside of the human body and to replace or restore failed organs with artificially created ones. The interest in replacing some lost parts of the human body is not a modern idea, though. One of the oldest mentions that I found is the ancient text on medical and surgical practices from India written somewhere between one millennium BCE to the first centuries CE, which is called “Sushruta Samhita”. The text describes an autograft skin transplantation, which we would now probably call rhinoplasty. The procedure allowed restoration of the nose, which was tightly connected with the image of dignity in India and could be lost due to punishment or in wars1.
Another area with a long history of replacement attempts is dental implantology. Historical evidence suggests that as early as 7th century BC, Etruscans used golden wire and artificial teeth to replace loosened incisor teeth2. The first documented teeth transplantations, though, happened in Europe in the Middle Ages3. And titanium implants were discovered only in the 20th century.
The rapidly changing world of the 20th century
The 20th century brought the beginning of fast scientific progress and associated with it changes in the way we see the world. The work of Swiss surgeon Theodor Kocher (late 19th to 20th century), who perfected thyroid gland surgery and discovered its function, shaped how we see organ transplantation. His findings led to surgeons’ and physiologists’ interest in organ replacement, extensive experiments in animals, and attempts to use animal organs to replace failed organs in humans. Then Alex Carrel figured out how to reconnect the transplanted organ to the respective blood vessels in a host body (early 20th century). Gradually, the surgery skills reached the level when it became obvious that even when the surgery itself is done perfectly, organ transplantation does not lead to the intended results of restoring a fully functioning body. Through many theories, researchers then concluded that the main problem is probably connected with the immune system4. Starting around the 1950s, people gradually learned how to suppress the immune rejection of transplanted organs5 and how to transplant organs, first from living (kidneys) and then from deceased donors6.
The progress medicine and physiology saw in the 20th century is astonishing. Apart from the obvious positive influence of improved medical care and longer life expectancy, it brought a major philosophical shift. The development of organ transplantation marks the end of an era when the prevalent belief was that the human body is a whole thing and diseases are connected with disturbances of liquid flows inside it4. The human body becomes an assembly of organs. Questions like “Do organs hold a part of a host soul?” or “Will transplantation affect the soul of a patient?” captivate the minds of people. We still can see the remnants of these beliefs in art. Just last year, I watched a movie where the main heroine receives a heart transplant and falls in love with a stranger, only to discover that it was the soul of the heart donor living inside her. Everything changes: the way people see themselves, the way people define death, and the way people see each other. It brings the necessity to develop ethical and legal rules for organ donation and transplantation, for experiments on human beings and animals. A lot of questions only become visible with time, like the differences in legal regulations of organ transplantation in different countries resulting in medical tourism.
In parallel with organ transplantation, techniques for growing cells outside the body were developed. First, the physiological solutions, like Ringer’s solution, which allowed keeping organs alive outside the animal body for several hours or days (late 19th century). Then the cells are successfully cultured in vitro by Margaret Reed Lewis (early 20th century), the standard culture medium recipe is developed, and the standardized cell lines appear7. Science is constantly changing people’s views on what is possible. Advances in cell culture bring the question of the possibility of recreating organs outside of the human body. And here we are, seeing the beginning of tissue engineering and regenerative medicine at the end of the 20th century.
The role of communication in scientific progress
While reading about the history of the tissue engineering and regenerative medicine field, I noticed that there wasn’t a lot of information about how these discoveries were accepted by public. Just imagine being born and raised with the idea that your body is the vessel of a soul and then being told that, actually, it is just like a clock – a mechanism with many replaceable parts. I remember a similar experience from my childhood. In primary school, I was told that you should always subtract a smaller number from a bigger one; you can’t do it vice versa. And then, in middle school, I learned that you actually can, and then you will get a negative number. Even though that was the way everyone learned mathematics, I felt betrayed. Why was it necessary to conceal the truth about the possibility of subtracting whatever you want from whatever you want? And what else is concealed until the time to know comes?
I guess part of the answer lies in how science was communicated throughout history. For a long time, science was mainly practiced by the privileged social classes, and communication mostly happened in a kind of top-down direction, where scientists or their benefactors showcased or shared only the things they wanted to share. Only in the 20th and 21st centuries science communication started to be seen as a way to allow people to make informed decisions and even influence the research direction when it concerns the everyday life of society 8. Part of the answer lies in the way science is taught as a subject with straightforward results, skipping a part where scientists were not sure or had conflicting ideas. As a result, when this process is shared, and some facts scientists believe to be true turn out to be wrong, instead of natural progression, it looks like not trustworthy. But also when reading stories like “The Immortal Life of Henrietta Lacks”, documented by Rebecca Skloot, or a recent article about using the CRISPR technique for editing human embryos by Dana Goodyear9, the thing that bothers me is often too superficial communication between scientists themselves. Too often connections are just networking when you briefly listen to 10 minutes of results that took a couple of years to get and then congratulate each other with a newly published article without looking too much into the details. Too rarely it is a meaningful connection when you are interested in another person’s work and ready for in-depth discussions.
I feel that lately, the academic structure is pushing for faster, bigger, better. It needs simple narratives and fast results. The never-ending competition brings the over-focus on your own work; researchers are constantly searching for their own niche, something to be at least a little bit different from others. It sometimes feels that instead of solving the problem, it is important to solve it differently from others. As a result, we have so much literature in our own domain that we hardly have time to follow other fields of science; we are so focused on our own research that hardly have time to listen and understand others. We are constantly networking and collaborating, but it feels that we are as isolated from each other as we can possibly be. Lately, I started asking myself questions. How often do you read papers that are not from your field? How often do you go for an adventure and read on completely unrelated topics? When was the last time you attended an interdisciplinary conference? And I mean truly interdisciplinary, not around the topic you work on, but rather an event where different people talk about the universe, climate change, medicine, and literature in one place. We all want to be heard, but how often are we the ones listening?
I first came up with the question of why people think of tissue engineering and regenerative medicine as a strange science because I felt misunderstood, but the longer I think about it, the more I think that it is a reflection of how the academic system is currently working. And maybe this is one of the reasons I decided to try science communication – to become the one who listens.
Further reading:
1. Saraf S. Sushruta: Rhinoplasty in 600 B.C. The Internet Journal of Plastic Surgery 2006; 3.https://ispub.com/IJPS/3/2/7839 (accessed 8 Sep2023).
2. Donaldson JA. The use of gold in dentistry: An historical overview. Part I. Gold Bull 1980; 13: 117–124.
3. Pasqualini U, Pasqualini ME. THE HISTORY OF IMPLANTOLOGY. In: Treatise of Implant Dentistry: The Italian Tribute to Modern Implantology. Ariesdue, 2009https://www.ncbi.nlm.nih.gov/books/NBK409631/ (accessed 18 Aug2023).
4. Schlich T. The origins of organ transplantation. The Lancet 2011; 378: 1372–1373.
5. Allison AC. Immunosuppressive drugs: the first 50 years and a glance forward. Immunopharmacology 2000; 47: 63–83.
6. Nordham KD, Ninokawa S. The history of organ transplantation. Proc (Bayl Univ Med Cent); 35: 124–128.
7. Yao T, Asayama Y. Animal‐cell culture media: History, characteristics, and current issues. Reprod Med Biol 2017; 16: 99–117.
8. Nielsen KH. Histories of Science Communication. Histories 2022; 2: 334–340.
9. Goodyear D. The Transformative, Alarming Power of Gene Editing. The New Yorker 2023.https://www.newyorker.com/magazine/2023/09/11/the-transformative-alarming-power-of-gene-editing (accessed 7 Sep2023).
(No Ratings Yet)Posted by the Node, on 18 September 2023
Development’s journal meeting, Unconventional and Emerging Experimental Organisms in Cell and Developmental Biology, took place between 17-20 September at the beautiful Wotton House in Surrey, UK. We’re delighted to be able to live-stream one session of this meeting to the broader community.
The event featured two fantastic talks, followed by a panel discussion session.
| 16:00 (BST) | Ehab Abouheif (McGill University) ‘Ants, eco-evo-devo, and the evolution of biological complexity’ |
| 16:30 | Kim Cooper (UC San Diego) ‘Evolution of extreme skeletal proportion in bipedal jerboas’ |
| 17:00 | Why we need to study unconventional models in the light of climate and biodiversity challenges Chair: Cassandra Extavour Panel: Ehab Abouheif, Kim Cooper, Michael Raissig, Iñaki Ruiz-Trillo |
| 18:00 | Close |
(2 votes)Posted by the Node, on 11 September 2023
Where is the lab?
We are based in east London, at Queen Mary University of London, in the United Kingdom.
Lab website: Martin-Duran Lab (martinduranlab.com)
Research summary
We want to understand how development is controlled and evolves to generate new phenotypes. To answer this, we study segmented worms (annelids) with spiral cleavage, a unique and ancient mode of development found in many marine invertebrates. We are interested in discovering how different strategies to specify the early progenitors of all cell types and organs of an animal evolved in spiral cleavage and how changes in the timing and maternal contribution defining those cells might influence phenotypic evolution during subsequent development, such as in the adult morphology and the life cycles. We approach these questions by combining multiple methodologies, from genomics, epigenomics and transcriptomics to more classical embryology, and using non-model organisms, some of which we are establishing as tractable research species for the first time.
Can you give us a lab roll call?
Currently, there are two postdoctoral (Yan Liang and Allan Carrillo-Baltodano) and four postgraduate researchers (Kero Guynes, Francisco Manuel Martín-Zamora, Rory Donnellan and Billie Davis) in my lab.
Yan studies the dynamics of gene expression and regulation during spiral cleavage and their correlation with the evolution of different life cycles and cell fate specification strategies.
Allan works on the evolution of maternal determinants and early cleavage in the specification of the body axes.
Kero investigates the role of DNA methylation during spiral cleavage and in annelids.
Fran has spearheaded the study of gene regulation and histone post-translational modifications during annelid embryogenesis and spiral cleavage.
Rory is investigating the regeneration of anterior structures in our favourite annelid, Owenia fusiformis.
Billie is exploring the evolution and role of 3D genome architecture in annelids and spiral cleavage.

Favourite technique, and why?
Chema: My favourite technique is in situ hybridisation because I love to see where genes are expressed. But I’m sure each member of the lab has their own!
Apart from your own research, what are you most excited about in developmental and stem cell biology?
Chema: I love evolutionary and comparative developmental biology generally. But I am particularly interested in the embryology of weird and understudied creatures, what they can tell us about animal development, and the evolution of the fascinating diversity of animal forms.
How do you approach managing your group and all the different tasks required in your job?
Chema: I like to support and train the members of my lab to become independent and have a sense of ownership of their projects. This is essential to stay motivated and willing to discover new things, even during the challenging moments in every research project. Efficient time management and ensuring time for hobbies, friends, and family is the key!
What is the best thing about where you work?
Chema: Queen Mary and the Department of Biology is a diverse and collegial environment. People work on many different organisms and questions, from plants and microbes to bats and fossils. That is very stimulating! It helps you learn something new daily and think about your interests and problems from new perspectives.
Allan: The community in the biology department at QMUL is very strong. Most postdocs initially come without a sense of what to do in a big, diverse city like London. However, the PhD students, in particular, make everyone welcome, and soon enough, you cannot skip a pub night on Fridays. The informal settings allow for a natural discussion of things that can help in our research and careers.
What’s there to do outside of the lab?
Chema: We are in East London! It is one of London’s most vibrant, diverse, young, and exciting parts.
Allan: London has everything you can imagine, but the music scene is excellent. From small bohemian venues to listen to folk music to mega stadiums to see your favourite rock band. There is something for everyone.
Browse through other ‘Lab meeting’ posts featuring developmental and stem cell biology labs around the world.
(1 votes)Posted by David Menassa, on 11 September 2023
In a recent review published in Acta Neuropathologica, Matuleviciute, Akinluyi & co-authors focus on the evidence obtained from humans to examine how microglia, the brain’s macrophages, participate in brain development. The review focuses on neurodevelopmental disorders, a set of complex disorders with significant heterogeneity in aetiology and presentation, such as autism spectrum conditions (ASCs) and schizophrenia (SZ). The authors make the case for this focus by specifying that many of the observed symptoms in ASCs and SZ are cognitive and characteristically human such as language difficulties, executive function breakdown and positive auditory symptoms. Furthermore, they also highlight key differences between human and non-human microglia. The review specifies that we are far from identifying mechanisms because human studies are descriptive and though, microglia are part of the pathological signature of these disorders, it remains challenging to concur whether they are reactive to an initial insult or causative of altered neurodevelopment. The authors highlight cutting-edge methodologies that are based on human tissues to help elucidate mechanisms of the interactions between microglia and the topography. This includes high-resolution transcriptomic studies and human tissue-based models. Finally, the co-authors critically examine the success of treatments in neurodevelopmental disorders that have targeted microglia[1]. Check it out!
1. Matuleviciute, R., et al., Microglial contribution to the pathology of neurodevelopmental disorders in humans. Acta Neuropathologica, 2023.
(No Ratings Yet)Posted by Alex Eve, on 8 September 2023
This post is part of the regeneration retrospective series.
First, I’d be remiss not to acknowledge that most of my posts have been on vertebrate systems when many invertebrates are also studied for their regenerative capacity. One such example is Hydra (Vogg et al., 2019) but Hydra papers don’t seem to appear in JEEM/Development until the 1960s. While exploring the early issues of JEEM/Development, I found fish manuscripts were similarly rare, which is surprising because – today – zebrafish are a prominent model for development and regeneration (Marques et al., 2019). Indeed, there was also a notable absence of any studies of nervous system regeneration, maybe because such articles were submitted to more specialist neurobiology and neuroscience journals. In this final post of the series, we fill this missing link by discussing the highlights from the following articles:
https://doi.org/10.1242/dev.5.3.256
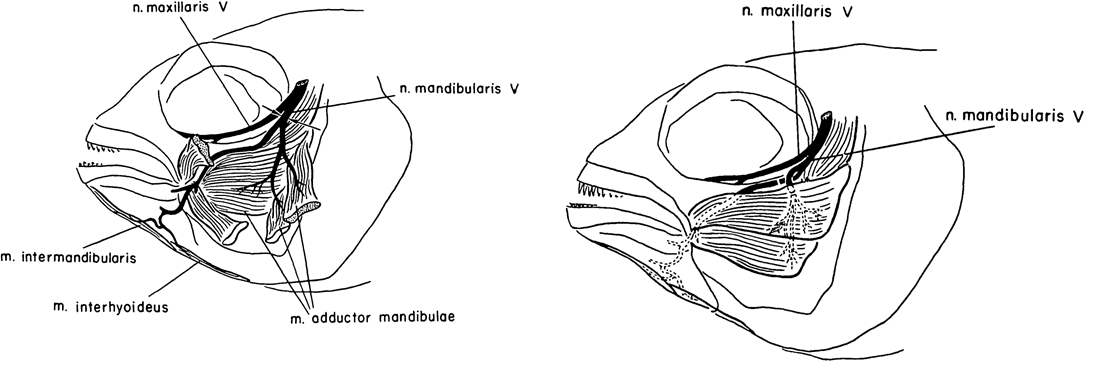
Caltech researcher Roger Wolcott Sperry is most famous for his work on the ‘split brain hypothesis’ and the corpus callosum, for which he won the Nobel Prize for Medicine/Physiology in 1981 (Hubel, 1994; De León Reyes et al., 2020). Alongside his work in cats, Sperry published a few articles on cichlid fish. One such article was published in JEEM/Development in 1957 (Arora and Sperry, 1967). His colleague, R. W. Arora, seems to have left little trace but I think it’s likely he brought the fish to the whole operation. Their study continued previous work on the basis that humans failed to regenerate motor neurons because of aberrant guidance and muscle innervation following wounding or amputation, which doesn’t occur in other vertebrates. To learn more about peripheral nervous system (Murtazina and Adameyko, 2023) regeneration, Arora and Sperry focused on the mandible of Astronotus ocellatus, severing the left mandibular nerve of ten fish and causing partial paralysis had completely recovered by 16 days post-operation (Fig. 1, left). Next, they cut the individual branches of the mandibular nerve and crossed them so that the nerves lay next to muscles they wouldn’t normally innervate (Fig. 1, right). Overall, as with the complete nerve cuts, mandible function was recovered a couple of weeks later with the nerves innervating the adjacent muscle rather than their original target (I have simplified the results here, do see the manuscript for full details). Arora and Sperry even used an electric induction coil to prove innovation and show that stimulating the regenerated nerves would cause the contraction of the new muscle target. This work showed the plasticity of the peripheral nervous system to adapt to the rewiring of the motor neurons with new muscles. Furthermore, this study showed that the guidance cues that direct motor neuron migration and innovation during development might not be present during regeneration, signalling the importance of the microenvironment for pathfinding and differentiating the requirements for neuronal targeting vs. neuronal innervation.
.
Teleost fish, including cichlids, are prominent in today’s research, although I think it’s fair to say that zebrafish are a dominant system. The explosion of zebrafish studies can largely be attributed to the Boston and Tübingen screens, culminating in the publication of a special issue of Development a little over 25 years ago (Nüsslein-Volhard 2012; Mullins et al., 2019). Since, the generation of hundreds (more?) of genetic lines, including fluorescent genetic reporters that capitalise on the zebrafish’s amenability to microscopy and imaging. As with cardiac regeneration, which we heard about earlier this week, the zebrafish (unlike most mammals, including humans) can regenerate its spinal cord (Becker and Becker, 2022).
https://doi.org/10.1038/s41467-019-14263-2
In addition to the peripheral nervous system, zebrafish have a remarkable capacity for regenerating the central nervous system, including the spinal cord, following injury – a phenomenon that doesn’t occur in mammals due to glial cell-dependent scarring. Thus, understanding how to prevent scaring and/or induce regeneration has huge therapeutic potential for spinal cord injuries. In their recent Development paper, Zhou and colleagues use a suite of sophisticated genetic tools to ask how glial cells respond to injury in zebrafish (Zhou et al., 2023). The authors first generate ctgfa-Tracer zebrafish for lineage tracing ctgfa-expressing cells, such as bridging glia, which emerge following spinal cord injury. Using this line, the authors show that bridging glia, ventral ependymal progenitors and regenerating glial cells are derived from ctfga-expressing cells but they minimally contribute to neurons and oligodendrocytes. The authors next turn to ctfga regulation, revealing the gene-regulatory sequences that drive ctfga expression during regeneration. Finally, the authors specifically ablate ctgfa-expressing cells, and show that axon outgrowth and swimming behaviour are affected following spinal injury. Together, Zhou and colleagues determine that ctfga-expressing cells have a pro-regenerative role in spinal cord regeneration.
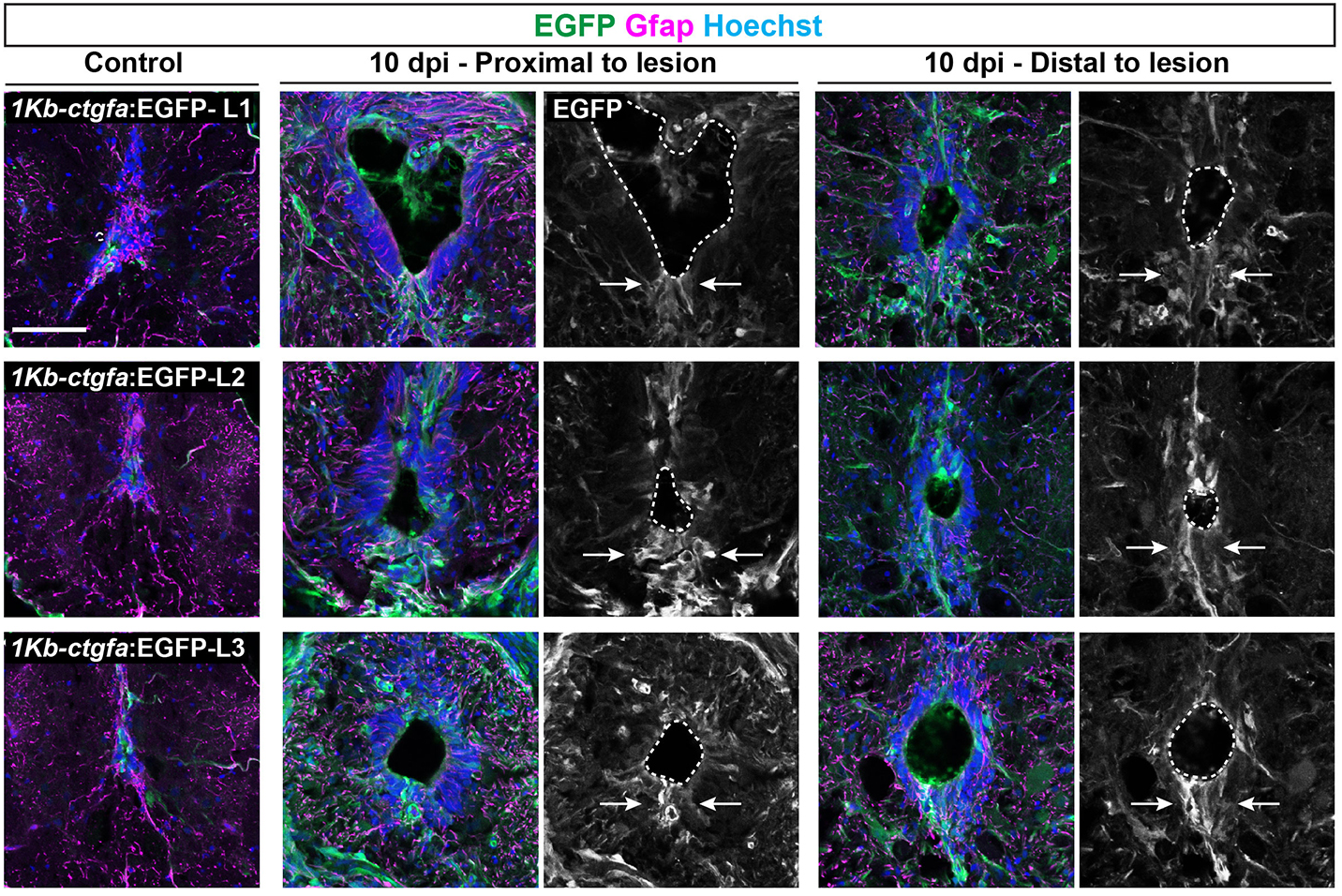
Both these papers address the ability of teleost fish to regenerate the nervous system and highlight the importance of the local microenvironment, as well as using animal behaviour (e.g. eating and swimming) as a read-out of phenotype. Arora and Sperry showed the regenerative plasticity of the peripheral nervous system, whereas Zhou and colleagues highlighted a particular cell population that supports and contributes to central nervous system regeneration.
I hope this little series has been entertaining – I’ve certainly enjoyed learning more about these early papers and the researchers who wrote them. Would you be interested in reading more ‘now and then’ or ‘past and present’ posts on other topics? Share your thoughts in the comments.
H. L. Arora, R. W. Sperry; Myotypic Respecification of Regenerated Nerve-fibres in Cichlid Fishes. Development 1 September 1957; 5 (3): 256–263. doi: https://doi.org/10.1242/dev.5.3.256
Thomas Becker, Catherina G. Becker; Regenerative neurogenesis: the integration of developmental, physiological and immune signals. Development 15 April 2022; 149 (8): dev199907. doi: https://doi.org/10.1242/dev.199907
D. Hubel; Roger W. Sperry (1913–1994). Nature 369, 1994; 186. https://doi.org/10.1038/369186a0
Noelia S. De León Reyes, Lorena Bragg-Gonzalo, Marta Nieto; Development and plasticity of the corpus callosum. Development 15 September 2020; 147 (18): dev189738. doi: https://doi.org/10.1242/dev.189738
Ines J. Marques, Eleonora Lupi, Nadia Mercader; Model systems for regeneration: zebrafish. Development 15 September 2019; 146 (18): dev167692. doi: https://doi.org/10.1242/dev.167692
Mary C. Mullins, Joaquín Navajas Acedo, Rashmi Priya, Lilianna Solnica-Krezel, Stephen W. Wilson; The zebrafish issue: 25 years on. Development 15 December 2021; 148 (24): dev200343. doi: https://doi.org/10.1242/dev.200343
Aliia Murtazina, Igor Adameyko; The peripheral nervous system. Development 1 May 2023; 150 (9): dev201164. doi: https://doi.org/10.1242/dev.201164
Christiane Nüsslein-Volhard; The zebrafish issue of Development. Development 15 November 2012; 139 (22): 4099–4103. doi: https://doi.org/10.1242/dev.085217
Stefano Sandrone. Roger W. Sperry (1913-1994). J Neurol. 2022 Sep;269(9):5194-5195. doi: 10.1007/s00415-022-11232-6. Epub 2022 Jul 22. PMID: 35867150; PMCID: PMC9363358.
Matthias C. Vogg, Brigitte Galliot, Charisios D. Tsiairis; Model systems for regeneration: Hydra. Development 1 November 2019; 146 (21): dev177212. doi: https://doi.org/10.1242/dev.177212
Lili Zhou, Anthony R. McAdow, Hunter Yamada, Brooke Burris, Dana Klatt Shaw, Kelsey Oonk, Kenneth D. Poss, Mayssa H. Mokalled; Progenitor-derived glia are required for spinal cord regeneration in zebrafish. Development 15 May 2023; 150 (10): dev201162. doi: https://doi.org/10.1242/dev.201162
(No Ratings Yet)Posted by the Node, on 8 September 2023

Our first webinar in October will be chaired by Development Editor Debby Silver (Duke University) and features three early-career researchers studying neurodevelopment and regeneration. The webinar will be held using Zoom with a Q&A session after each talk.
Baptiste Libé-Philippot (VIB-KU Leuven Center for Brain & Disease Research)
‘Study of the role of hominid-specific genes in the development and function of human cortical neurons’
Idoia Quintana Urzainqui (EMBL, Heidelberg)
‘Studying the shark embryo to understand the origin and evolution of telencephalic cell types’
Leo Otsuki (Institute of Molecular Pathology)
‘Editing positional memory alters signalling centre output during axolotl limb regeneration’
To see the other webinars scheduled in our series, and to catch up on previous talks, please visit: thenode.biologists.com/devpres
(2 votes)Posted by Kat Arney, on 7 September 2023

Wallace is endlessly fascinating. The magnitude of his scientific contributions, where he came from, what he did to make those achievements… That in itself makes him quite remarkable, right?
Professor Jim Costa, Author of Radical By Nature: the Revolutionary Life of Alfred Russel Wallace
In the latest episode of the Genetics Unzipped podcast, we’re exploring the epic life of Alfred Russel Wallace; adventurer, naturalist and co-discoverer of natural selection. Biologist and author, Jim Costa, recounts the life of this often forgotten founding father of evolution.
Genetics Unzipped is the podcast from The Genetics Society. Full transcript, links and references available online at GeneticsUnzipped.com.
Subscribe from Apple podcasts, Spotify, or wherever you get your podcasts.
Head over to GeneticsUnzipped.com to catch up on our extensive back catalogue.
If you enjoy the show, please do rate and review on Apple podcasts and help to spread the word on social media. And you can always send feedback and suggestions for future episodes and guests to podcast@geneticsunzipped.com Follow us on Twitter – @geneticsunzip
(No Ratings Yet)