The Beddington Medal is the BSDB’s major commendation to promising young biologists, awarded for the best PhD thesis in Developmental Biology defended in the year previous to the award. It was announced in 2002 [letter #23/2] and first awarded in 2004.
Rosa Beddington was one of the greatest talents and inspirational leaders in the field of developmental biology. Rosa made an enormous contribution to the field in general and to the BSDB in particular, so it seemed entirely appropriate that the Society should establish a lasting memorial to her. The design of the medal, mice on a stylised DNA helix, is from artwork by Rosa herself.
Nominations for the Beddington Medal
The eligibility period covers PhD dissertations which were defended during the calendar year previous to the award (i.e. until end of December 2020). Furthermore, applicants need to have at least one paper accepted or close to acceptance. Nominations should be in two parts:
From the candidate, up to 2 pages A4 describing the thesis and supplemented with up to 1 extra page of figures from the thesis to illustrate key results, plus a 1 page CV, including statement of prizes/awards already received. These should all be in the form of a single pdf file of no more than 1 MB. A candidate exceeding these limits risks having to resubmit their application. In addition, candidates should supply formal documentation of the date of submission of the thesis.
From the candidate’s PhD thesis supervisor, a letter of support, sent independently, consisting of no more than 2 pages A4, describing why the student was deserving of this award. This letter should explicitly comment on the status of publications arising/expected from the thesis work, and also on any unusual circumstances, including duration of study.
Candidates can be of any nationality, must be BSDB members at the time of nomination, and at least one of their supervisors must be UK-based. Nominees must be able to attend the BSDB Spring Meeting, where the winner is to present a plenary talk.
All nominations received will be considered and voted upon by the Committee and the winner invited to present the Beddington Medal lecture on their thesis work at the following BSDB Spring Meeting.
We are seeking a highly motivated candidate strongly interested in interdisciplinary science, to join our groups (van de Pavert and Guignard labs) and work on a project at the crossroad between Computer Science and Developmental Biology as a Ph.D student.
Quantification and modelling of embryonic lymph node organogenesis at the single cell scale.
The role
The project aims at studying lymph node (LN) formation during mouse embryonic development. LN formation requires hematopoietic lymphoid tissue inducer cells (LTi) to interact with mesenchymal cells at precise locations within the embryo, where they subsequently form aggregates. We have postulated that the peripheral nervous system outgrowth initiates the earliest events in LN formation. Indeed, preliminary data show that LTi aggregates morphology and cell density is affected in mouse embryos lacking neuronal subsets.
To understand the relationship between neuronal outgrowth and lymph node formation, the successful candidate will work between the van de Pavert lab and the Guignard lab to develop new computational methods to reconstruct and quantify LTi aggregates and peripheral nervous system morphology at the single cell scale. The reconstructions will then be used to develop a machine learning framework to systematically quantify phenotypes in perturbed mouse embryos. These quantifications will in turn allow to model the effects of neuronal outgrowth on LN formation.
The successful candidate will work towards:
building a library of whole-mount mouse embryo images
developing computational methods for the analysis of the generated library, including:
reconstruction and mapping of the neuronal network
quantification of LTi aggregate morphologies and positions
developing computational methods to automatically stage mouse embryos
modelling LN formation in relation to neuronal network morphology
Keywords
Quantitative embryogenesis, whole-mount analysis, peripheral nervous system, immune system, image analysis, machine learning, big data analysis
Whom would we like to hire?
The student should be enthusiastic, creative and ambitious, have good communication skills and be eager to learn. A master degree with major or minor in computer science is required. Affection for developmental biology is preferred. Some experience in developmental biology is also preferred but not required. Note: exception can be made for students who have not studied computer science if the student can prove coding skills.
The offer
3-year Ph.D. position in the van de Pavert lab and the Guignard lab
Call open for applications: January 15 – February 26
Interviews of shortlisted candidates by the evaluation committee: April 20 to April 22
The pre-selection process will be based on qualifications and expertise reflected on the candidates CV and motivation letter. It will be merit-based. All candidates will be informed whether they have been pre-selected or not.
We are a group of computer scientists with a strong interested in biology in general and more specifically in embryonic development. We develop novel computational methods and models that allow the analysis of very large 3D movies of animal embryonic development (up to 2TB per movie). We work in close relationship with biologists to tailor our methods so that they help to address fundamental biological questions.
The developmental biology question that mainly but not only animates us is to better understand the mechanisms driving embryogenesis to robustly form a complex organism despite genetic polymorphism and variable environmental conditions.
We are interested to study the first cues required to form lymph nodes at specific locations within the embryo. We are fascinated by the interactions between different cell types, such as hematopoietic cells, mesenchymal cells and neurons which eventually will generate a highly organized LN. To study these interactions, we use a multi-disciplinary approach and combine techniques such as 3D immunofluorescence imaging, flow cytometry and single cell sequencing on embryos from different mouse models.
The Turing Centre for Living Systems (CENTURI) is an interdisciplinary project located in Marseille (France).
CENTURI aims at developing an integrated interdisciplinary community, to decipher the complexity of biological systems through the understanding of how biological function emerges from the organization and dynamics of living systems.
The project federates 15 teaching and research institutes in biology, physics, mathematics, computer science, engineering and focuses on Research, Education and Engineering, 3 missions that hold interdisciplinary as their core principle.
The research and training programmes implemented under the auspices of CENTURI will foster new collaborations, will transform practices, will attract new talents and thereby contribute to making the Luminy campus a leading site for the interdisciplinary study of biological systems.
i3S – Institute for Research and Innovation in Health (Porto, Portugal) is looking to recruit a senior researcher with an established international reputation in Neural Cell Biology and strong expertise in securing, managing and leading collaborative research projects and teams/institutional units.
NCBio offers:
5-year contract as Research Coordinator, research equivalent to Full Professor, with an internationally competitive salary;
Relocation expenses, along with support concerning housing, legal/immigration issues, spouse’s employment, kindergarten/schooling, etc;
250 thousand euros in seed money in the first year of operation for the group’s research activities and to cover initial operational costs;
Total scientific freedom including the selection of a research team (four additional researchers – 2 at the Associate Professor level, 1 Postdoctoral Fellow, 1 Technician) paid directly by the NCBio project throughout its course.
Admission requirements include:
PhD degree obtained at least 10 years before application;
Established international reputation based on research excellence in the field of Neural Cell Biology;
Strong record of publication of influential papers;
Large experience in leading research groups;
Proven track-record in securing and managing significant funding;
Experience in establishing collaborative relationships with relevant industry R&D stakeholders;
Track-record in workshop and conference organising committees and delivering invited talks.
The successful applicant will be expected to commit to the position for the grant’s full duration.
Call opens on February 1st and the preliminary deadline is May 31st.
For any inquiries, please contact us at erachairs@i3s.up.pt
The recruiting process will be conducted following the Portuguese labor code, and selection criteria that prioritise merit and transparency, as well as a non-discrimination and equal access policy.
The mammalian retina contains a variety of functionally distinct cell types that are generated by progenitor cells in a specific chronological order. A new paper in Development probes the role of the POU-homeodomain factors Pou2f1 and Pou2f2 in the timely generation of cone photoreceptors in mice. We caught up with first author and PhD student Awais Javed and his supervisor Michel Cayouette (Director of the Cellular Neurobiology Research Unit at the Montreal Clinical Research Institute, Professor at the Université de Montréal and Adjunct Professor at McGill University) to hear more about their work.
Awais (L) and Michel (R).
Michel, can you give us your scientific biography and the questions your lab is trying to answer?
MC: I obtained my PhD in Neurobiology from Université Laval, in Québec city, Canada, working on viral vector-mediated gene transfer approaches in mouse models of retinal degeneration. Towards the end of my PhD I became very interested in understanding how all the beautiful cell types I was looking at under the microscope were generated, and decided I would study neural development during my postdoc. I contacted several labs and, fortunately for me, the famed developmental neurobiologist Martin Raff, whom I admired greatly, offered me the last postdoc spot available in his lab before he retired. Realizing this unique opportunity, my wife and I decided to move to London where I spent 3 years studying asymmetric cell divisions in retinal progenitors and the relative contribution of intrinsic and extrinsic signals in cell fate specification. When Martin retired, I was in the middle of a project that I wanted to finish before looking for an independent position. I then joined Ben Barres’ lab at Stanford University, USA, where I continued to work for 2 years on the project, which was finally published in 2003. In November 2004, I started my independent career in Montreal.
Ever since the beginning, my lab has been focused on studying how cell diversification is achieved during nervous system development, with the long-term goal of using this knowledge as the basis to develop regenerative therapies. We are particularly interested in understanding how neural progenitors know when it is time to make a specific combination of cell types and, once a temporal window has been defined, how progenitors choose between alternative cell fates available to them at that time. We primarily use the mouse retina as a model system to address these questions, but we have also studied myelination and the inner ear to ask questions related to cell polarity, which we also study in the context of asymmetric cell division in the retina.
And Awais – how did you come to work in Michel’s lab and what drives your research today?
AJ: In the final year of my undergraduate degree at University College London, I was revising the course material for the end of term developmental biology exam, including the temporal competence cascade in neuroblast ventral nerve cord specification in Drosophila. I found it fascinating how the same set of genes could regulate cell fate in different parts of the ventral nerve cord, and I wondered if there was a similar cascade in the vertebrate central nervous system. A quick PubMed search led me to the work of the Cayouette lab, who showed that Ikaros, the fly hunchback homolog, was a temporal competence factor in the mammalian retina. As it so happens, I was planning to move to Canada for personal reasons and the first lab I looked up was Michel’s. I started my PhD trying to find similarities between vertebrate and invertebrate neurodevelopment, but now I feel more driven by the complexity biology has to offer and how different the systems can be.
How has your research been affected by the COVID-19 pandemic?
AJ: I was very fortunate because on the last day before the lockdown; we received the next-gen sequencing results for a number of experiments I had submitted in February. The first 2 months of the shutdown, I learned how to analyse these datasets using R and Python. I had no previous knowledge of using any programming language, so it was a true test of perseverance. When the lab finally opened up, I was just happy to be back at the bench: I didn’t think I would ever say this but I definitely missed genotyping!
I didn’t think I would ever say this but I definitely missed genotyping!
MC: The lab has been completely shutdown with only essential activities allowed for almost 2 months. Like Awais, people in the lab tried to use this difficult period to make some progress. Some wrote a draft of their paper, analysed large datasets or counted cells, while others took online classes and learned how to code. I was very proud of my group – they were amazing and really used this time as best they could, despite all the challenges associated with the situation. But of course, everything was delayed in the lab. Most difficult for us was having to scale down our animal colony, as the animal facility staff was reduced. We are just now getting back to normal after several months. This also meant it took longer than expected to carry out the revisions for our paper, but the Development editors were helpful in guiding us to prioritize experiments and were flexible with the time allowed for us to do these, which was appreciated!
Why is timing critical for the generation of cellular diversity in the retina?
AJ & MC: While it is not always clear why specific neurons must be generated before others, it is generally accepted that cell birth order is critical to ensure proper neural circuit formation. Just as the foundation of a house must be built first because other parts of the house sit on it, certain types of neurons constitute the foundation of a given circuit, as they receive inputs from neurons produced later. This tightly regulated chronology is critical for the generation of highly complex tissues.
Can you give us the key results of the paper in a paragraph?
AJ & MC: In this paper, we investigated the role Pou2f1 and Pou2f2 in the developing mouse retina. We show that both genes are necessary and sufficient for cone photoreceptor cell fate specification during retinal development. We further report that Ikzf1, an early temporal identity gene that we previous identified in the retina, upregulates Pou2f1, which in turn represses the late temporal identity factor Casz1, thereby defining a temporal identity window conducive to cone photoreceptor production. Mechanistically, we show that Pou2f1 activates Pou2f2, which then represses expression of the rod-promoting factor Nrl in postmitotic photoreceptor precursors by binding to a POU-specific site in the promoter, thereby favouring the cone fate. As Pou2f1 and Pou2f2 are orthologues of fly pdm, which is well known for its role in neuroblast temporal patterning, our results, together with previously published studies, suggest that some aspects of this cascade are conserved in vertebrates. This work also establishes a link between temporal identity genes and cell fate determinants in the mammalian central nervous system.
E15 mouse retina immunostained for Rxrg, which labels newly generated cones at the apical side and ganglion cells at the basal side of the retina.
Loss of Pou2f2 reduces, but does not eliminate, cone production: how are cones determined in its absence?
AJ & MC: It is of course possible that other genes compensate for the loss of Pou2f2. Potential candidates include Onecut1 and Onecut2, as a partial loss of cones, similar to what we observed in this study, is observed in double knockout animals. Another possibility is that abolishing the temporal window for cone production induces progenitors to generate cones outside the normal window using alternative pathways. Finally, we have previously suggested that loss of temporal identity in progenitors does not lead to complete absence of any given cell type, but simply to a reduced probability of generating these cell types, which might explain why we do not observe a complete loss of cones in this study.
Do you know what restricts Pou2f1 expression to early RPCs? Is there evidence of mutual inhibition with other temporal factors?
AJ & MC: This is a very interesting question. We show in our paper that Ikzf1 upregulates Pou2f1 expression in early RPCs, but it remains unknown what actually turns off Pou2f1 expression in later progenitors. An obvious candidate is Casz1, which might repress Pou2f1 at later stages, similar to what is observed in Drosophila neuroblasts, but our preliminary experiments looking at this possibility do not appear to support this model. Another possible regulator of Pou2f1 is Foxn4, which was recently discovered as a regulator of Ikzf1 and Casz1 in the mammalian retina. More work is needed to fully elucidate this issue.
When doing the research, did you have any particular result or eureka moment that has stuck with you?
AJ: The main result that comes to mind was actually one of the first observations that led us down the path of this paper. Initially, immunostainings of Pou2f1 showed strong expression in ganglion cells, as well as other cell types, which we later found out to be cone and horizontal cells. When we first tried gain-of-function experiments with Pou2f1 using a retroviral vector, we did not initially consider that cone production would be induced. Out of sheer curiosity, I added S-opsin as a marker for cones and, to my surprise, I found S-opsin+ cells in the overexpression condition. My first thought was that I had mixed up the viruses or the immunostainings, but when it started to repeat, I was convinced it was a real result. It wasn’t a eureka moment per se, but it felt quite good to stumble onto a finding that would later turn out to be the central hypothesis of the paper.
And what about the flipside: any moments of frustration or despair?
AJ: The ChIP-qPCR experiments, without a doubt. It is running joke in the lab at this point because of how tired I was during lab meetings when I was working on those experiments. It was very labour intensive because I used embryonic retinal tissues and I had quite a bit of optimization to do before I could get any results. But it was well worth it, when I finally got the experiment to work. In the end, it was a fantastic lesson learned on perseverance.
What next for you after this paper?
AJ: I am finishing up a few projects in the lab and hoping to graduate in early 2021. I am currently looking for postdoc opportunities and I am very excited to continue investigating cell fate specification in other parts of the central nervous system.
Where will this story take the Cayouette lab?
MC: We are becoming increasingly interested to determine whether temporal factors could be used to promote tissue regeneration. The idea is simple, as early temporal factors are sufficient to reprogram late-stage progenitors into generating early-born cell types, we wonder whether these factors might also be able to reprogram differentiated cells into neural progenitors. We would also like to determine whether progenitors invariably go through all the temporal identity windows defined by the temporal factors we have studied so far in the retina (Ikzf1, Pou2f1, Pou2f2 and Casz1) or whether some can skip a given window. Answers to this question might explain the huge heterogeneity of clonal composition observed in lineage-tracing studies in the retina. This will require the development of tools to follow temporal factor expression in real time in single cells, but I think it would be very cool to address this question, as it has far-reaching implications. Finally, detailed mechanistic understanding of temporal patterning remains poorly understood, even in flies. We are becoming increasingly interested in this question. It is likely that temporal identity is defined by specific epigenetic landscapes, and whether and how temporal factors shape chromatin conformation and nuclear architecture is a problem we will likely focus on in years to come.
Finally, let’s move outside the lab – what do you like to do in your spare time in Montreal?
AJ: I am an avid painter and it has helped me deal with the ups and downs of a PhD. I also founded a drama club at the institute. We wrote and directed a couple of plays to a full audience at one of the institute’s auditoriums, which was an amazing experience. I did not realize that many scientists are fantastic artists!
MC: Since I was a child, I have been playing ice hockey and I reached fairly high competitive level. To this day, I continue to play once or twice a week, often with students who seem to be getting younger and faster each year… I am also a fervent cyclist and love to ride the roads of the countryside and the trails of the provincial park around our house. Finally, I love great food and wine, and since Montreal has a large choice of fantastic restaurants, it is a good place to be!
Yesterday we held the fourth webinar in our series, this time chaired by Development Editor Swathi Arur. Here you’ll find recordings of the talks and their live Q&A sessions moderated by Swathi.
The Genetics Unzipped podcast is back for 2021 with a new series of stories from the world of genes, genomes and DNA, from the history of genetics to the latest cutting-edge research.
In our first episode we take a look back at the discovery of messenger RNA, or mRNA, in the 1960s. There are some big names involved – Francis Crick, Sydney Brenner, Francois Jacob and more – but who actually discovered this vital molecular messenger? And why did nobody win a Nobel Prize for it?
Then we come right up to the present day to look at mRNA vaccines for COVID-19, which have been developed at breakneck speed to tackle the pandemic. We find out how mRNA vaccines work, how they were developed so fast for COVID-19, and how this new technology might change the face of immunization and public health in the future.
If you enjoy the show, please do rate and review on Apple podcasts and help to spread the word on social media. And you can always send feedback and suggestions for future episodes and guests to podcast@geneticsunzipped.com Follow us on Twitter – @geneticsunzip
The laboratory of Pamela Geyer invites mature, highly motivated candidates to apply for a position as postdoctoral fellow in the Department of Biochemistry at the University of Iowa. These fellows will join a research team interested in understanding the impact of nuclear architecture in stem cell survival and aging. Prominent changes in nuclear morphology occur during physiological aging and in disease. Our recent data demonstrate that the asymmetrically dividing germline stem cells (GSCs) in the Drosophila ovary employ a non-canonical mode of mitosis that sensitizes these stem cells to defects in proteins comprising the nuclear lamina, a protein network that build contacts with the genome to regulate transcription, replication and DNA repair. Current studies are focused on exploring the mechanisms involved in execution of this non-canonical mitosis and implications for stem cell health and longevity. In addition to driving science at the bench, the fellow will be introduced to the network of collaborative scientists with shared interests in defining biological processes critical for development and disease. The Geyer laboratory is located in a state-of-the-art facility that features a collaborative atmosphere to foster innovation, and is part of the Carver College of Medicine, a highly ranked biomedical research institute. The University of holds a long tradition of diversity, equity, and inclusion, a transition that began upon its founding with being the first public university to admit men and women on an equal basis (1855). Iowa City is a medium-sized, culturally rich city with vibrant communities in the arts, sciences, and literature. Interested applicants should contact Pamela Geyer (pamela-geyer[at]uiowa.edu), including a letter of interest, a CV and contact information of three references.
Cellular differentiation describes the process by which embryonic cells become different from one another, acquiring distinct identities and specialised functions. These cells are responding to signals that modify the dynamics of the interacting key genes controlling the cell’s state. The arrival of more and better single cell data opens up new opportunities for better understanding of this cellular decision-making. However, to realise these opportunities, new mathematical and statistical tools are needed to characterise the cellular dynamics and to organise, analyse and visualise such data. This workshop aims at reviewing the current work in this area.
Announcement of post-doc scholarship at the Division of Pediatrics, Lund University, Sweden
The importance of HIF-2afor neural crest cell migration and its connection to tumor initiation
It is well known that hypoxia, i.e. low oxygen levels, predicts poor outcome and metastatic burden in a variety of solid tumors. Transcriptionally active Hypoxia Inducible Factors (HIFs) and their downstream target genes orchestrate the cellular adaptation to increased oxygen demands in both normal and tumor cells (Hammarlund, Flashman, Mohlin & Licausi, Science 2020). We have recently shown that the cancer-associated protein HIF-2a is important for trunk neural crest stem cell development (Niklasson et al., Dev Dyn 2020). In this project, we are interested in investigating how HIF-2a affects early and late neural crest cell migration, population of organs and how this connects to childhood tumor initiation.
The purpose of this scholarship is for the candidate to deepen their skills in the field of molecular physiology. The candidate will acquire theoretical knowledge within the areas of developmental and tumor biology as well as learn a set of methodologies including microdissections, cell culture, cloning, virus work and analyzing omics data. The candidate will learn to handle several different research organisms, including chick embryos and in vivo mouse models.
Preliminary start date: 1st of March 2021 (to be discussed)
– To be eligible for a post-doc scholarshop at Lund University the recipient must hold a PhD degree within a relevant field. The PhD degree must not be from Lund University. The PhD degree must not be older than three years. The applicant must not have been employed at Lund University in the past two years.
– Obtained PhD within cancer research, developmental biology or equivalent.
– Published papers in peer-reviewed international journals
– Fluent in written and spoken English
The following qualifications are advantageous but not necessary:
Experience in:
Analyzing omics data (e.g., RNA sequencing and mass spectrometry)
In vivo models (e.g., chick embryo, mice, rats, zebrafish)
Wet lab techniques
Imaging
The candidate will be part of a team, but at the same time needs to be independent enough to lead their own projects. The candidate should be fluent in English and have experience from oral and written scientific communication.
We put great emphasis on personal suitability.
Written application, including reference number, is to be sent via e-mail to the supervisor and must include the following:
– CV
– Personal letter stating the reasons why the study suits the applicant (maximum one page)
This is the first in a new series on the Node profiling scientists who do art (or artists who do science). We’ll learn about why they do it, their artistic influences, their techniques and tips. We’ll be looking for new people to feature in this series throughout the year – whatever kind of art you do, from sculpture to embroidery to music to drawing, if you want to share it with the community just email thenode@biologists.com (nominations are also welcome!).
Our first profile is of Suyash Naik, a graduate student from the IST Austria, whose art we first saw on Twitter.
Where do you work and what are you currently working on?
Suyash
I study at the Institute of Science and Technology Austria, with the supervision of Dr Carl-Phillip Heisenberg and Dr Edouard Hannezo. I’m currently in the third year of my graduate studies working to understand the mechanics of epithelium movement during zebrafish embryonic development. I am especially interested in understanding the intermediate filament cytoskeleton’s role as the epithelium spreads during gastrulation. For this I am learning to apply various interesting biophysical techniques that our labs have developed, such as laser cutting and imaging.
Has science always been an important part of your life?
I’d say curiosity and learning new things have been important aspects of my life. I grew up with an interest in natural history and dinosaur fossils, and also liked watching and reading popular science fiction. Naturally I was always interested in learning more about science and understanding the world we live in. This interest was fuelled further by great teachers and incredible opportunities that allowed me to do the things I do day to day now. For instance, in school I took part in a group that used to plan and do simple experiments revolving around DNA. This led me to pursue an education at IISER where I had the opportunity to explore and work in different labs.
And what about art – were you always interested in drawing? What or who are your artistic influences?
I have been interested in sketching and photography but I have never had the opportunity to pursue any formal education in them. But recently I made more of an effort to consistently create and also be more forthcoming with the things I’m doing and have done in the past, as a way to have an outlet for creativity and to track my mental health during these stressful times. For me what is most interesting and important is the change of perspective I gain when viewing something as an artistic inspiration rather than a scientific curiosity. I have always enjoyed visual art like paintings and animations. The lockdowns this year also greatly increased my appreciation for street art. Watching great pieces from some of my favourite artists like Nekro and Illunis has been a real joy during lockdown walks along the Danube canal this year. But most importantly of all I draw inspiration from my creative friends who make some truly amazing artwork!
“For me what is most interesting and important is the change of perspective I gain when viewing something as an artistic inspiration rather than a scientific curiosity.”
How do you approach your art?
I mostly use digital mediums currently as I have some familiarity with them, but I am still hoping to learn different types of techniques to create sciart. I am looking forward to learning more about art techniques in the coming years and creating/experimenting different styles. I usually enjoy colorful and futuristic looking artwork and that is something I especially want to learn how to create.
Tell me about your zebrafish artwork?
For the zebrafish artwork, I had thought of the design long ago to sort of combine the ideas that I wanted to base my PhD around and things that were ongoing in my lab at the moment.
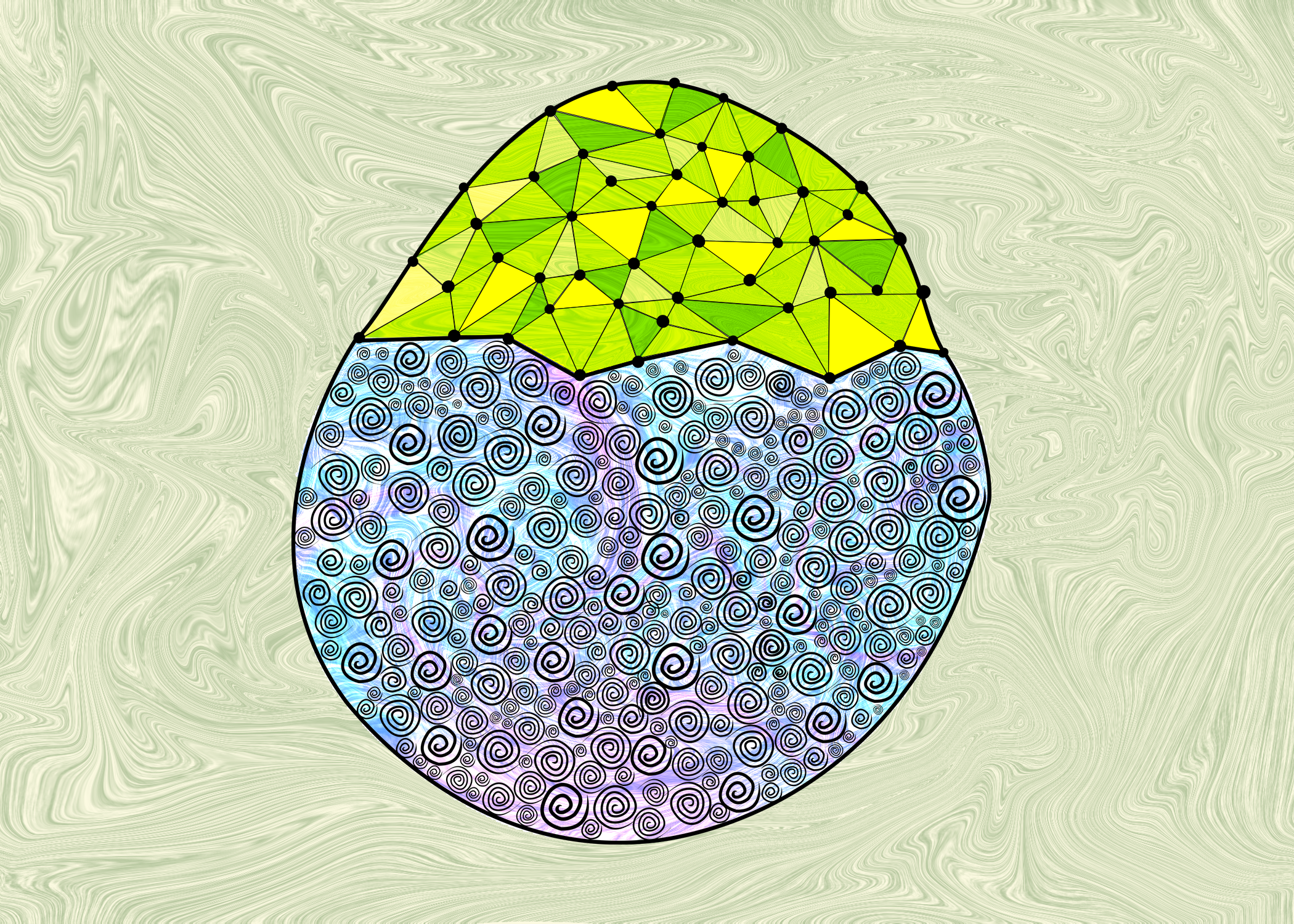
Zebrafish embryo
The embryonic part of the embryo is an artistic representation of a model epithelium. Recently in my lab a few great publications (such as this article by Schwayer et.al, this article by Boocock et. al., this and this review by Shamipour et.al.) explore how flows of different cytoskeletal proteins play an important role during development. This made me think of some acrylic pour that a friend of mine had made, and I wanted to incorporate that as a component of the artwork. That leads to the flowy backdrop of color that exists in the picture. The yolk over which the epithelium moves is a collection of nutrition for the developing embryo in the form of collected yolk granules. These were represented in spirals depicting these granules.
And what about the Hydra pictures – I think both have reference to historical pieces or myths?
The Hydra pictures were some that I had created working on a project during my undergraduate studies.
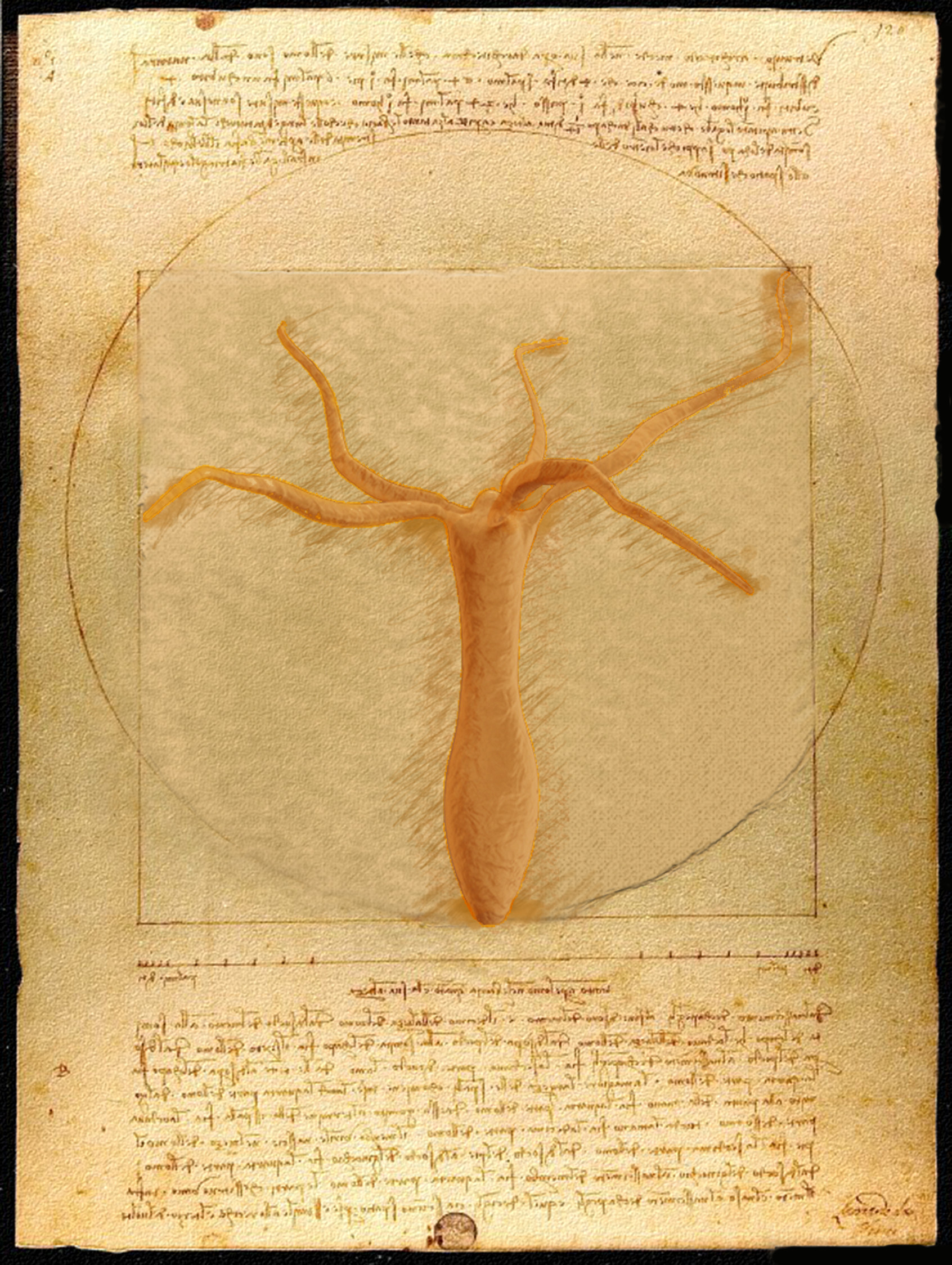
Hydra vitruvitous
I was fascinated by the role of mechanics in the somersault motion of the organism. Of course it was a running joke in my group that the organism had gotten its name from its incredible ability to regenerate and hence was mythical. This along with the fact that I was enjoying working alongside Leonardo da Vinci as an assassin in virtual gamespace (Assassin’s creed series) made me want to depict Hydra as a part of something you would find in his work. I felt drawn to the Vitruvian man as it was very a mechanical description of the human body.
Hydrassil
When the work was published we also wanted to create something to celebrate the occasion and for trying to get selected as a cover article. For that I made another similarly mythological/gaming themed piece as a succession for the vitruvian Hydra. Hence I decided to go again along similar themes and went with a celtic Norse mythological reference. I hence depicted the major steps in the somersault movement at the branches and roots of the world tree, Yggdrasil. The sun rises in the background shining light onto the pioneering steps in a ‘primitive’ organism performing a motion on a solid surface.
What are you thinking of working on next?
I am looking forward to getting back to some exciting experiments I have planned for next year, maybe those would drive some new interesting ideas. I do not have a concrete plan of where I am going next with my art studies but what I do know is that I want to create and learn as much as I can!

 (No Ratings Yet)
(No Ratings Yet)

 (1 votes)
(1 votes)

 (3 votes)
(3 votes)