The Friedrich Miescher Institute for Biomedical Research (FMI) invites applications for a tenure-track group leader position (assistant professor level). We encourage applications from candidates who have an innovative research program on questions related to mechanisms of development, regeneration and disease. Applications from individuals using quantitative and interdisciplinary approaches are particularly welcome.
The position includes competitive start-up and core funding, as well as access to outstanding in-house core facilities for genomics, proteomics, structure biology, cell sorting, microscopy and image analysis, C. elegans, Hydra and mouse genetics, and computational biology. The Institute also provides a career mentoring program and assistance with research grant applications. Our vibrant, English-speaking scientific environment offers opportunities for multidisciplinary collaboration with FMI research teams in epigenetics, quantitative biology, tissue biology and neurobiology. For further information visit www.fmi.ch.
The FMI is a leading biomedical research institute affiliated with the University of Basel and the Novartis Institutes for Biomedical Research. It hosts around 190 postdoctoral fellows and graduate students recruited worldwide, and has a widely diverse international staff representing over 40 nationalities. We strongly believe that the best science comes out of a work culture that respects and appreciates individual differences and supports everyone to achieve their full potential. For the work-life balance of our employees, we offer flexible working hours and a teleworking program. We provide a supportive and family-friendly environment in which to develop ambitious and original research.
We offer financial support for relocation and settling in. The FMI is located in Basel, the third largest city in Switzerland, in the heart of Europe. Basel is home to a diverse international population and offers a rich cultural life and several multilingual schools.
The FMI is committed to raising the proportion of women in science; we thus strongly encourage female researchers to apply. The Institute is responsive to the needs of dual career couples.
Applications, including a CV, a concise description of research interests and future plans, and contact details for three referees should be submitted at:
A wave of innovations is advancing data-driven computational analysis and machine learning – time for developmental biologists to hop on the surf board! This post, inspired by our recent data-driven work on lateral line morphogenesis, provides a brief primer on key concepts and terms.
written by Jonas Hartmann & Darren Gilmour
From machine translation to self-driving cars and from deep fakes to high-speed trading, data-driven technologies are taking the world by storm, fueled by a torrent of funding from the tech industry. This has led to the rise of data science, a new and loosely defined interdisciplinary field that combines aspects of computer science, statistics and machine learning to extract information or generate predictions from large and otherwise inscrutable datasets.
Meanwhile in biology, it has never been easier to generate large and inscrutable datasets. New omics techniques and new microscopes paired with increased automation provide the means to digitize every aspect of a biological system, from single molecules to whole embryos. Crunching this data deluge and converting it into scientific understanding is simultaneously one of the biggest opportunities and one of the biggest challenges in modern biology – and data science is poised to become a powerful tool in meeting this challenge.
But not all data are created equal. Certain formats, such as counts or sequences, lend themselves more readily to the application of data science techniques. Others, including multi-dimensional images, require several layered steps of processing and analysis to tease apart the rich information encoded within them. Furthermore, much of the recent progress in machine learning is founded on big data, specifically on datasets with thousands or tens of thousands of samples, a scale that is hard if not impossible to achieve for many biological techniques. In short, datasets can be thought to fall onto a spectrum from big data (low information/sample ratio) to rich data (high information/sample ratio) – and when it comes to data-driven inference, big data has taken center stage among data scientists over the past years.
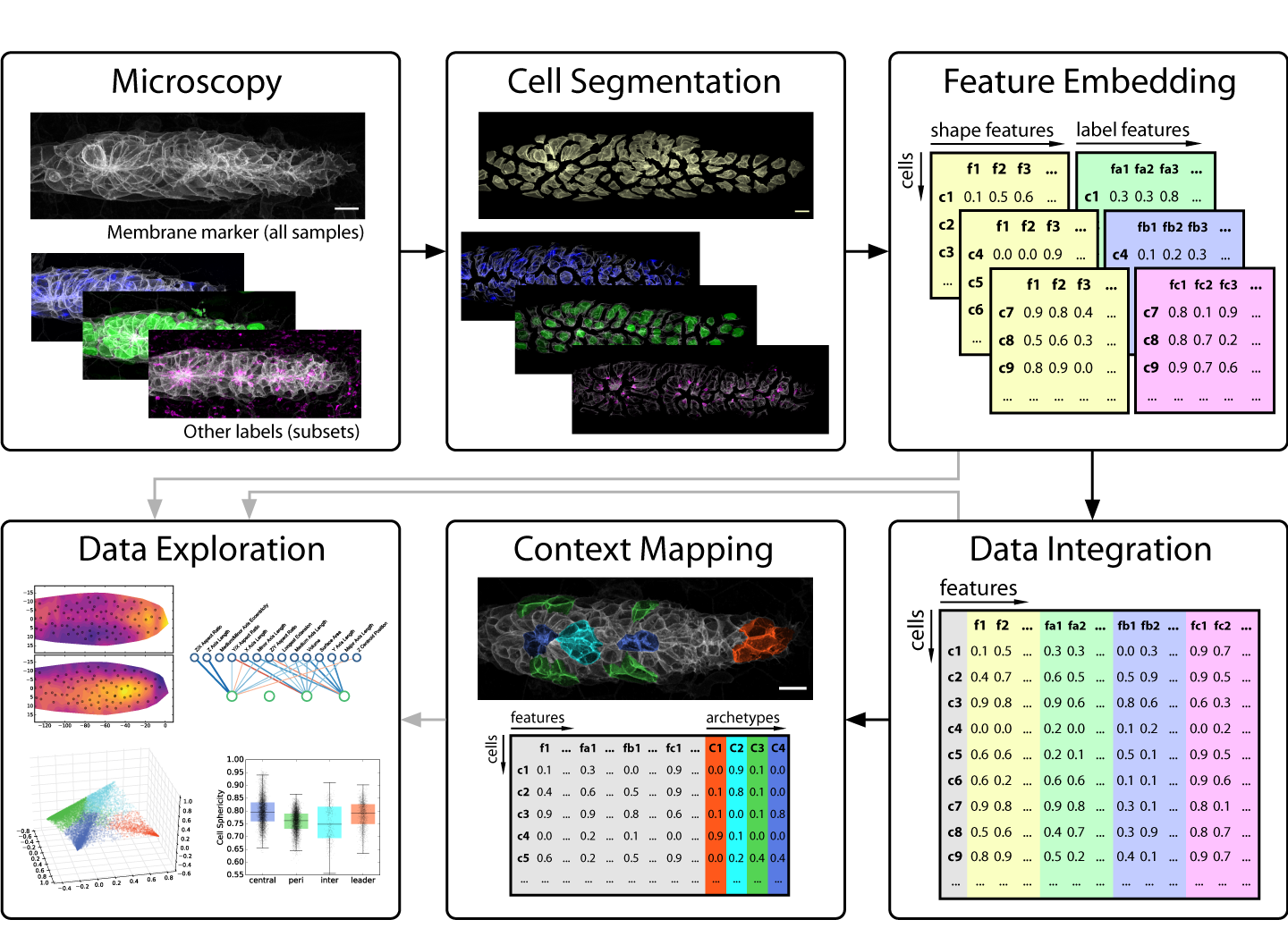
This brings us to developmental biology and more specifically to one of its key data sources: in vivo microscopy. Since high throughput remains a rare luxury in most imaging settings, developmental microscopists very much operate in the realm of rich data. This presents a major challenge for anyone looking to employ data science to disentangle the complexity of embryogenesis. Nevertheless, exciting progress is under way! For instance, many microscopists will be aware of the solutions machine learning is delivering for image analysis problems such as segmentation and tracking [1,2]. But as we learned in our work on the morphogenesis of the zebrafish lateral line primordium [3], segmentation is only the first step on the journey from image to insight (figure 1) – so our focus in this post is on what comes after a decent segmentation has been achieved.
For those not familiar with the lateral line primordium: it is a developmental tissue of a hundred or so cells that migrate as a cohesive group along the flank of the developing embryo, assembling and occasionally depositing rosette-shaped clusters of cells, which go on to form the lateral line sensory organs [4]. The need for tight coordination between collective cell migration and rosette morphogenesis make the lateral line primordium an ideal system to study the interplay between multiple developmental processes, especially since the transparency of zebrafish embryos greatly facilitates high-quality live imaging. With this context in mind, let’s jump into the data science.
Figure 1: An overview of our strategy for analyzing cellular architecture in the lateral line primordium. Note that single-cell segmentation is only the first step of our analysis. See the paper [3] for further details.
From Images to Numbers
Images, especially those produced by 3-5D microscopy of live embryos, are a prototypical example of rich data, as the obtainable number of samples is limited but each sample contains a wealth of information across multiple scales. By giving us both the (relative) abundance and the location of labeled components, images indirectly encode object counts, object shapes, contact interfaces between objects, textures, gradients, colocalization, and much more – and all of this potentially as dynamical measurements over time! Unfortunately, this information is not directly accessible to most modern data science methods, so exploring it in a data-driven fashion is difficult.
The key problem to solve is thus to go from images that encode all this rich information at once to numbers that encode the same information in a more explicit, disentangled fashion. In data science, such descriptive sets of numbers typically take the form of a feature space, which is a 2D array with a row for each sample and a column for each measure, also called a feature. The “space” in “feature space” reflects the notion that each sample’s feature values can be understood as a vector in an -dimensional space, where is the number of features. For microscopy data, features can be extracted to describe an entire image, regions of interest (e.g. different areas of a tissue) or fully segmented objects of interest (e.g. cells, nuclei, vesicles, filaments, etc.).
Strategies for feature extraction generally fall into one of two groups: feature engineering is the manual implementation of various measurements that could be of interest (e.g. volume, surface area, aspect ratios, etc.), whereas feature embedding is the use of an automated computational approach to generate features. The latter is also referred to as latent feature extraction, as implementations often look for variations across the entire dataset and then allocate features that capture this variation, thus uncovering “hidden” (latent) features that underlie the variability in the data. The advantage of feature embedding is that it is unbiased (or, more accurately; less biased), so it can capture interesting phenomena independent of the researcher’s prior assumptions. The disadvantage is that latent features may not correspond to obvious biological properties, so they can be harder to interpret.
In our recent paper, we use point clouds and clustering on those point clouds to bring 3D images of segmented cells into a format that allows us to use a simple Principal Component Analysis (PCA) to derive latent features. We use this approach primarily to analyze cell morphology based on the distribution of a membrane marker in 3D space, but it also works for other markers.
Over the past few years, feature engineering has become less common in data science, as deep neural networks can work directly with raw data; they essentially learn their own internal feature embedding strategy. This can also be used explicitly for latent feature extraction via neural network architectures called autoencoders (see this blog post for a short intro). The downside of this approach is that it usually requires big data to work reliably, so classical feature engineering and feature embedding strategies remain important tools for data-driven analysis of rich data.
A key limitation of the feature space data structure is that it doesn’t readily encode different organizational scales. For instance, it is not straightforward to meaningfully combine data on the tissue scale (a tissue x features array) with data at the cellular scale (cells x features) or below (e.g. protrusions x features) in a single analysis. Since exploring data across multiple scales is central to many biological questions, this is an area where technological innovation on the data science side is required to better accommodate biological applications.
From Numbers to Ideas
So what’s the point of all this work to convert raw image data into a tidy feature space? In short, it’s the wealth of computational tools that are available to operate on and visualize feature spaces to discover interesting patterns that lead to new hypotheses.
A common first step is dimensionality reduction, which essentially amounts to combining features that encode the same information and removing features that mostly contain noise (although this is a very pragmatic way of thinking about it). This is most commonly achieved with PCA. The point of dimensionality reduction is twofold: First, the remaining features are likely of high importance; to understand them is to understand the majority of variation in the entire dataset. Second, many of the tools discussed below struggle with very high-dimensional feature spaces, a problem known as the curse of dimensionality. This reflects our earlier discussion on big data versus rich data; most methods need many more samples than features to work well.
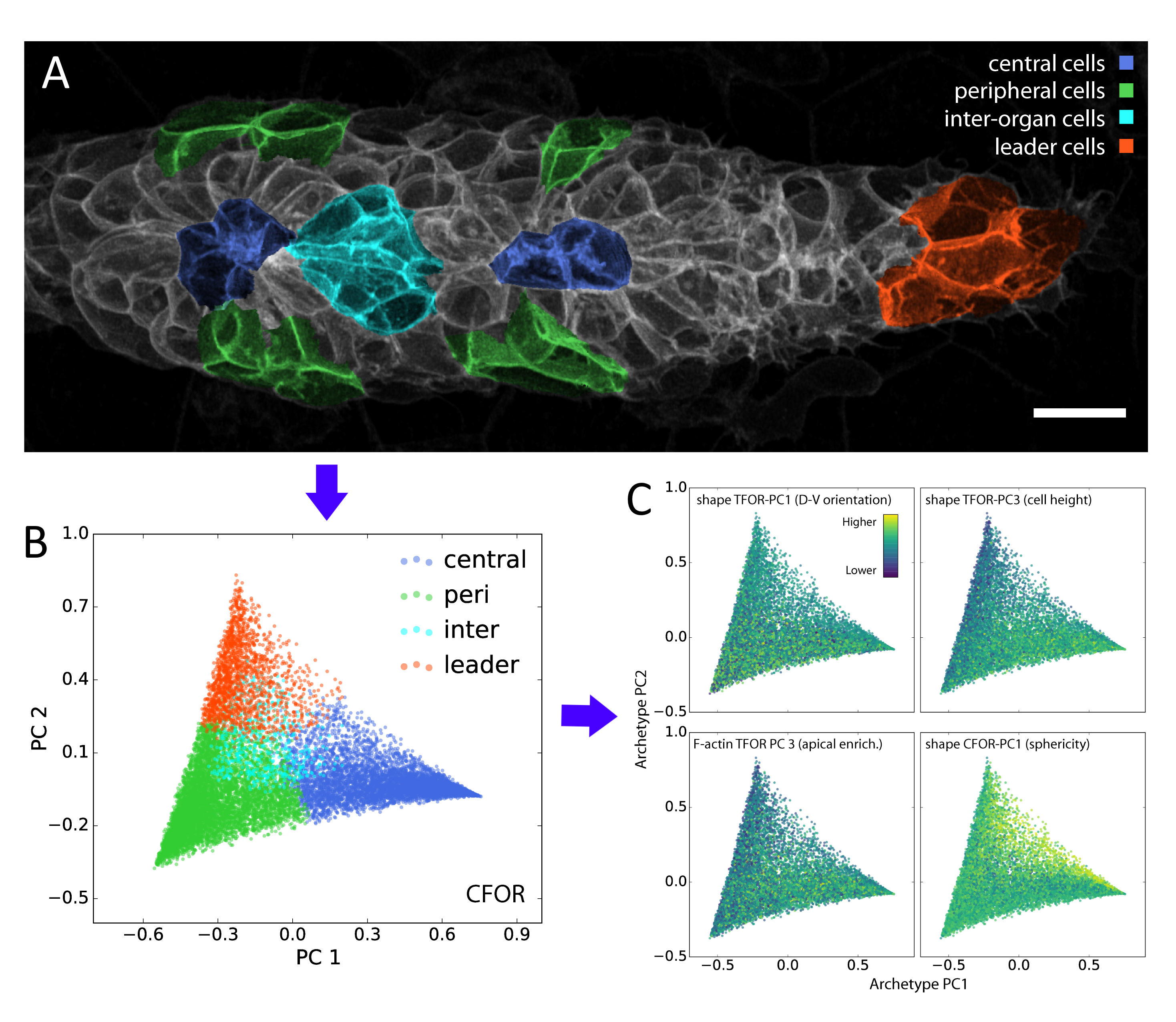
In our study, we performed dimensionality reduction and discovered that one of the most important remaining features relates to cell sphericity. We then mapped this key feature back onto the spatial organization of the lateral line primordium – averaged across all our samples – and found that it shows an unexpected central stripe pattern along the tissue. Given the intrinsic connection between sphericity and surface tension, this led us to the hypothesis that cell surface mechanics might regulate the relative location of cells within the migrating primordium.
Dimensionality reduction and related tools such as clustering (i.e. the identification of distinct subgroups among samples) are so-called unsupervised methods, as they work with a single input dataset. By contrast, supervised methods use both an input feature space and a target feature (also called the ground truth) and aim to learn the relationship between them. These algorithms are trained using a training set, for which both the input features and the target are known, for instance based on manual annotation of a subset of the data. Once trained, they can predict the target feature for new samples, based only on their input features.
A classifier is a supervised algorithm that can be trained to predict which of two or more groups a cell belongs to, given its features. We used this approach to classify cells into specific morphological archetypes, chosen by us to reflect our prior understanding of the tissue’s biology. This helped us make sense of the data from a biological perspective (more on this below).
A regressor predicts a continuous measure rather than a binary or categorical group label. We employed regressors to learn the relationship between shape features and features extracted from other channels, such as the distribution of vesicles or mRNA expression levels (measured by smFISH). Interestingly, regressors can be used as a means of integrating multiple different experiments (e.g. different smFISH stainings) based on reference features common to all experiments (e.g. cell morphology). By training a regressor to predict the experiment-specific results from the reference features, those specific results can be overlaid onto any dataset for which the same references are available. This enables the creation of an integrated dataset, often referred to as an atlas. Such atlases can be built across feature spaces but also directly across images, using neural networks as image-to-image regressors. An impressive example of which is given by the Allen Integrated Cell [5].
Although these data-driven techniques are powerful, unsolved challenges remain. One such challenge is the translation between how humans understand data and how computers understand data. For instance, it is difficult to include expert knowledge into a deep learning algorithm. Conversely, it can be hard to interpret the results produced by an algorithm in a biologically meaningful way. We used our classification of cells into different morphological archetypes to visualize and analyze our entire dataset from a more interpretable perspective, an approach we call context-guided visualization (figure 2).
Figure 2: Manual annotations (A) of contextual knowledge, in this case whether a cell is a leader cell or a peripheral, central or inter-organ follower cell, can be projected onto the entire dataset (B) through a machine learning classifier. This is the basis for context-guided visualization (C), which allows the data to be viewed and analyzed through the lens of prior biological knowledge.
The Future of Data-Driven Developmental Biology
It’s important to stress that data science techniques, although interesting and useful in a multitude of ways, cannot and should not replace “traditional” science. To complete the full cycle of science, the hypotheses extracted from the data need to be tested with specifically designed experiments, which is something we are currently pursuing for the observations we made in our data-driven analysis of the lateral line primordium.
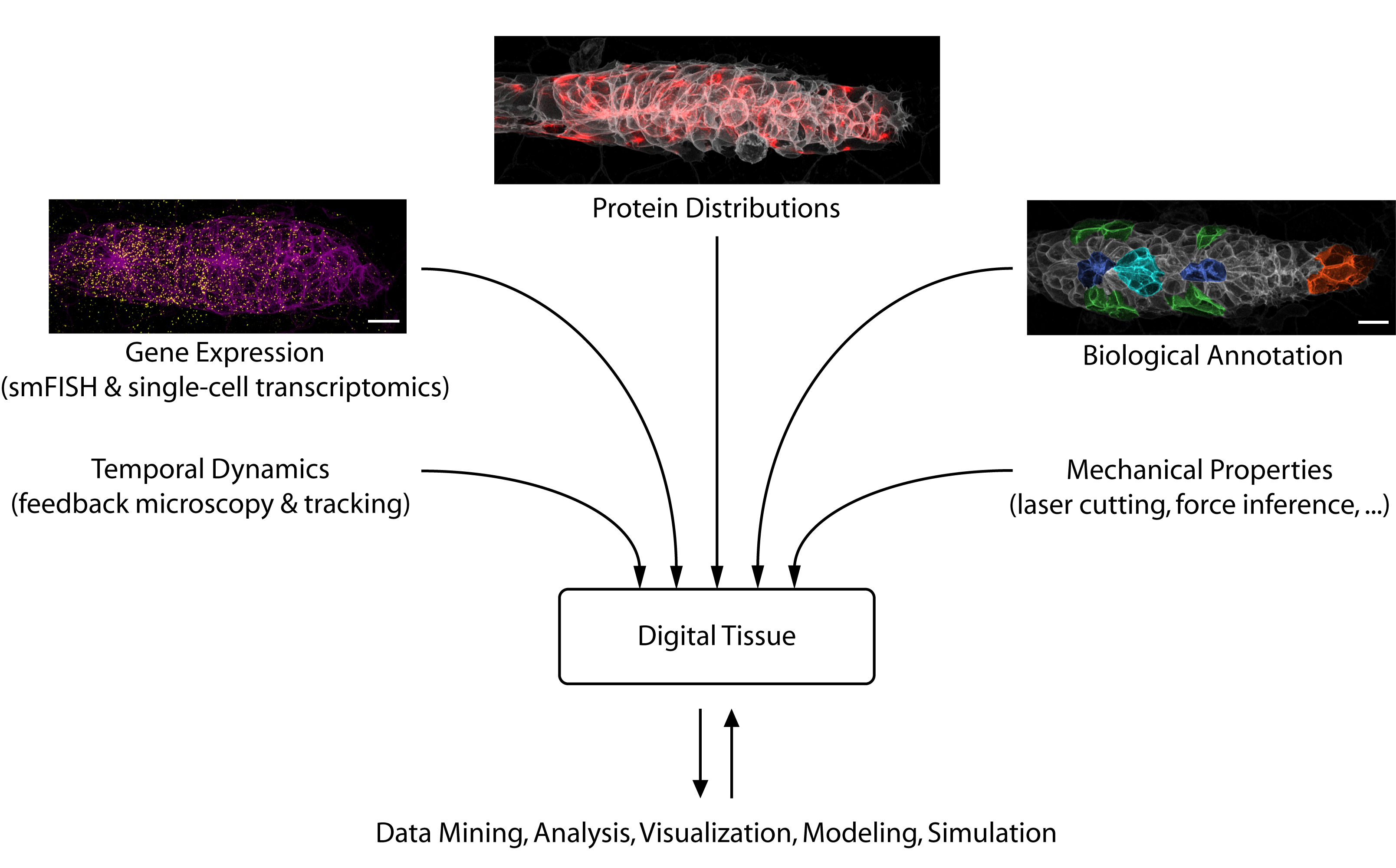
Looking toward the future, we see considerable potential for data science to accelerate developmental biology. Besides solving challenging image analysis problems (such as single-cell segmentation), data-driven methods can serve as “hypothesis generators” that allow us to comb through the complexity and messiness of cell and tissue biology. Eventually, large databases could be constructed that interlink different types of information into “digital tissues” (figure 3) or even “digital embryos”, which can be mined for interesting patterns and relationships. Early examples of such large-scale atlases are already starting to come online [6].
But data-driven developmental biology has a long way to go. As we have pointed out, biology is uniquely complicated in its dynamic and multi-scale nature, which plays to some of the weaknesses of current data-driven tools. Thus, there is a need for technical work to build biology-specific data science platforms and to adapt methods from other data-driven fields to the peculiarities of biology. Perhaps even more importantly, biologists need to receive the training required to understand and employ these techniques in their own work. Fortunately, data literacy training is on the rise in schools and universities, computer programming is easier to learn than ever and the relevant computational tools are being packaged more and more accessibly. Slowly but surely, data science will thus find its way into the developmental biology toolbox.
Figure 3: A digital tissue (or digital embryo) is a multi-modal dataset that enables integrated analyses of the diverse factors that play together to mediate organogenesis.
Although there are guides to train potential peer reviewers for reviewing research articles, the format and purposes of reviews aren’t the same, so they must be approached from a different perspective and assessed by different criteria. You might even be an established group leader by the first time you’ve been invited to evaluate a review simply because there are just fewer of them than research articles. So, what does an editor expect from your report? Broadly speaking, you can think about it in terms of A, B, Cs.
Accurate
The most important role of a peer reviewer is to make sure that the review articles are scientifically accurate. By this, I mean that statements should be factually correct, with appropriate references that support the conclusions. Accuracy also extends to the structure of the article. For example, does the title and abstract actually reflect what is covered in the review? Figures, although schematic and simplified, should also present the latest interpretation of the data accurately.
Balanced
Next, think about whether the review is balanced. As a whole, think about whether the articles covers everything you’d expect to see on the topic; make sure to point out if a large body of work is missing and check that the reference list isn’t biased! Even for opinion pieces, it is important that review-type articles are balanced and provide both sides of an argument. It is helpful for the authors to point out ongoing debate or controversy in the field, especially for non-specialists who might not be aware of the types of discussions that are happening. Where the authors propose new ideas and hypotheses, it is important that they distinguish these points from established facts. In addition, remember that studies have both strengths and weaknesses, and that techniques or approaches have their advantages and limitations – are both sides acknowledged?
Clear
Review-type articles are read by specialist and non-specialists alike, so it’s important that they are clear and accessible to a broad audience. For journals that developmentally edit or copy edit manuscripts, many of these points will be addressed by the in-house editor – although it is still useful for them to know what you think needs work, even if you don’t provide a point-by-point list of necessary changes.
It’s crucial that language and phrasing is clear and unambiguous to avoid confusion or misinterpretation. As an expert in your field, it might be easy to gloss over an acronym that you’re familiar with or to know that a gene or cell exists by two (or more) names. Take care to check that authors always introduce and define new terms, and use them consistently. Think about whether the structure of the article prepares you for what to expect and whether you can navigate the article easily. Most reviews should guide the reader through the article and provide all the necessary background to make links in thought between data and conclusions. Finally, are the figures well designed, well presented and intuitive? Would additional figures, boxes or tables help to clarify text and illustrate important key points?
Specifics
On occasion, an editor might raise specific queries with you about the manuscript. For example, if the article is very long, they might ask if you can identify specific parts that could be cut. Alternatively, they might notice a lack of recent references and might ask if the article is timely, as well as a whole range of other questions.
Some lastthoughts
Remember to be polite, courteous and constructive in writing your report.
Try to be specific – refer to line or page numbers if you have concerns with a particular statement.
Most articles will be proofread before publication, so there often isn’t the need to list every typo you find.
Do highlight if you think spelling, grammar or punctuation should be addressed, but avoid suggesting that it should be read by a ‘native English speaker’. This suggests that authors with English as a second language write less well then native English speakers – this is simply not the case!
The Department of Genetics and Development invites applications for one tenure-track faculty position at the Assistant Professor level. We are seeking outstanding applicants who use genetic approaches to study developmental or physiological processes. Our department spans a broad range of interests including developmental biology, physiology, DNA repair and recombination, cancer and human genetics. Applicants must hold a Ph.D. and or an M.D. degree and will be expected to develop an internationally recognized extramural funded research program. Interested applicants should submit a curriculum vitae and a statement of research interests, and arrange for three referees to submit letters of recommendation. Columbia University is an Equal Opportunity Employer and the school has allocated resources to target the recruitment of underrepresented minorities and a particular attention will be given to URM applicants.
A postdoctoral position is immediately available for highly motivated recent PhD or MD/PhD graduates in the Singh Lab (https://medicine.vumc.org/person/bhuminder-singh-phd) at the Division of Gastroenterology, Hepatology, and Nutrition at Vanderbilt University Medical Center to participate in basic studies controlling epithelial polarity as well as more translational efforts to develop innovative therapeutic approaches for colorectal cancer (CRC).
Available projects include:
• Regulation of EGFR signaling by polarized trafficking of its ligand epiregulin (EREG) and mechanisms underlying transformation upon apical mistrafficking of EREG;
• Role of receptor tyrosine kinases MET and RON in resistance to EGFR-targeted therapeutics in CRC.
The projects will involve 3D cultures of genetically or pharmacologically manipulated cell lines and CRC organoids and in vivo cancer xenografts. The effects of specific stimuli and contribution of individual molecules will be monitored by live or fixed (confocal) microscopy, cellular signaling, phenotypic assays, multiplex fluorescence imaging, and comparative RNA-seq (single-cell or bulk) or proteomic analysis. Please see recent publications at: https://scholar.google.com/citations?user=Z9Wjl6AAAAAJ&hl=en
Candidates are sought with experience in signal transduction, biochemistry, cell biology, or cancer biology. Individuals are expected to have excellent oral and written communication skills, as well as a track record of productivity.
Please send a CV, a cover letter, and names of three references to Dr. Bhuminder Singh:bhuminder.singh@vumc.org. The position is available immediately.
The research group of Dr Max Yun, ‘Regeneration of Complex Structures in Adult Vertebrates’ at DFG/CRTD Center for Regenerative Therapies Dresden (Germany) & MPI-CBG is searching for a PhD candidate.
The major focus of the group is to understand the molecular and cellular mechanisms underlying regeneration in adult vertebrates, using salamanders (newts and axolotls) as model systems. The group uses a broad range of molecular, genetic, biochemical and advanced imaging techniques to investigate regenerative processes (https://www.crt-dresden.de/de/forschung/research-groups/core-groups/crtd-core-groups/yun/). The group is co-affiliated to the Max Planck Institute of Molecular Cell Biology and Genetics (https://www.mpi-cbg.de/home/).
The project will involve the investigation of regenerative processes in hematopoietic organs so a background in immunology would be an advantage.
Requirements:
-University degree in the areas of Biology/Biochemistry
-Excellent degree marks (top 10% of year)
-Strong motivation to pursue an academic career.
-Strong interest in the group’s field of work
-Experience in experimental laboratory work (Experience in transgenesis and/or amphibian research would be an asset)
-Excellent knowledge of English (written & oral)
-Strong background in experimental design
-Capacity for working independently
-Will to work in an international team within a highly collaborative environment
Interested candidates should send a CV, list of publications, qualifications/research work description and two references/contacts for reference to Dr Maximina Yun (maximina.yun@tu-dresden.de).
In this episode, sponsored by Thermo Fisher Scientific, we’re taking a look at how genomic technologies are transforming cancer care – now and in the future, and the importance of making sure that these advances are available to all.
We look at what the future looks like for both adult and childhood cancers, as well as exploring how important fast genetic testing is to patients and how new technology could help.
With guests:
Greg Simon, past president of the Biden Cancer Initiative and former executive director of the White House Cancer Moonshot Task Force.
Jim Downing – president and CEO of St Jude Children’s Research Hospital
Dr Marianne Grantham, Head of Cytogenetics and Molecular Haematology department at the Royal London Hospital
Kim Wood, Thermo Fisher Scientific’s Clinical Sequencing Division
Research position investigating the mechanisms underlying cardiac morphogenesis
The Bloomekatz laboratory in the Department of Biology at the University of Mississippi in Oxford, MS is seeking a researcher to assist in our investigations of cardiac morphogenesis and disease using zebrafish. Please see our website https://thebloomekatzlaboratory.org for details on our research. The successful candidate will have the opportunity to be involved in all aspects of the research and publication process, from conducting developmental biology experiments and zebrafish husbandry, to analyzing imaging data to writing and publication.
-Interested in joining our dynamic group apply online: https://careers.olemiss.edu, search zebrafish. Salary dependent on experience. This position is eligible for benefits. Anticipated start date Jan 2020.
-Further questions can be directed to Dr. Bloomekatz at josh@olemiss.edu. The University of Mississippi is an EOE/AA/Minorities/Females/ Vet/Disability/Sexual Orientation/Gender Identity/Title VI/Title VII/Title IX/ADA/ADEA employer.
Department of Biological Sciences, College of Sciences and Mathematics
Auburn University
The Department of Biological Sciences at Auburn University invites applications for a tenure-track faculty position beginning Fall 2021 at the rank of Assistant or Associate Professor in Plant Developmental Biology.
We seek a highly collaborative candidate who will examine fundamental mechanisms governing developmental processes in plants. A successful candidate is expected to establish an extramurally funded, internationally recognized research program focused on plant developmental biology. Instructional responsibilities include development of graduate and/or undergraduate courses in plant developmental biology related to their area of emphasis. Faculty will join existing Plant Biologists as well as recent Developmental hires in Marine and Terrestrial systems as an emerging group of Developmental Biologists in the department.
Review of applications will begin December 1, 2020 and will continue until a suitable individual is hired. Applicants should submit curriculum vitae, a description of research interests, a statement of teaching philosophy and experience, and contact information of three professional references. Applicants should endeavor to describe how your past and/or potential contributions in teaching, research, and/or service will serve to advance the College of Sciences and Mathematics’s mission of creating an inclusive environment. This should include the following: Statements of values as they relate to your understanding and commitment to diversity, inclusion, and equity in STEM fields; examples of experiences that demonstrate your commitment to fostering the success of traditionally underrepresented groups in STEM (students, staff, and/or peers) and supporting a diversity of perspectives in the classroom, lab, campus, and/or community; and Future plans for continuing to advance inclusive excellence, diversity, or equity in your research, teaching, and service.
Apply your developmental biology / neurobiology skills to the problem of brain cancer.
The lab of Dr. Jennifer Chan is seeking to recruit a motivated postdoctoral fellow to investigate cell fate decisions during the process of brain tumour development and progression. Research activities in the lab focus on growth factor signalling and transcriptional regulation as determinants of neural precursor identity and fate. We use model systems that include patient-derived glioma cultures, xenografts, and engineered mouse models generated from in utero and post-natal electroporation to address our research questions.
The successful candidate will collaborate with investigators in bioinformatics to apply to apply tools of genomics, epigenomics, and transcriptomics to further define important alterations during neoplastic transformation, including interactions of tumour cells with other constituents in the microenvironment. Early career-stage candidates (within 3 years of receiving PhD or equivalent) with experience in advanced immunohistochemistry / fluorescence microscopy; gene editing, and molecular biology; mouse modeling of disease; and/or experience in genomic approaches like RNA-seq, ChIP-seq, ATAC-seq will be preferentially considered.
The position is available immediately.
Other information
Located in Calgary, Alberta, Canada, the Chan Lab is part of the Charbonneau Cancer Institute at the University of Calgary’s Cumming School of Medicine. Within the Charbonneau Institute, we are part vibrant multidisciplinary research groups focused on childhood cancers and brain cancers. Calgary is a very ‘livable’ and family-friendly city located less than an hour’s drive from the Canadian Rocky Mountains – a haven for outdoor enthusiasts.
Application details:
Submit a brief letter of interest, your academic CV, and the names and contact information of at least three references.
Applications should be submitted as a single PDF file and sent as an email with the subject line “Post-Doctoral Fellowship, Glioma Biology” to: jawchan@ucalgary.ca
 (No Ratings Yet)
(No Ratings Yet)
 (6 votes)
(6 votes) In this episode, sponsored by Thermo Fisher Scientific, we’re taking a look at how genomic technologies are transforming cancer care – now and in the future, and the importance of making sure that these advances are available to all.
In this episode, sponsored by Thermo Fisher Scientific, we’re taking a look at how genomic technologies are transforming cancer care – now and in the future, and the importance of making sure that these advances are available to all.