DEPARTMENT OF MOLECULAR, CELL, & DEVELOPMENTAL BIOLOGY DIVISION OF PHYSICAL AND BIOLOGICAL SCIENCES
1156 HIGH STREET
SANTA CRUZ, CALIFORNIA 95064
Postdoctoral Fellow – Neural Circuit Development – Anatomy and Genomics
The Kim Lab at Molecular, Cell, and Developmental Biology Department, the University of California, Santa Cruz is seeking highly motivated and talented postdoctoral research fellows with a Ph.D. degree to join our new and innovative research group.
The University of California, Santa Cruz is one of 10 universities within the prestigious University of California system. MCD department has top-tier neuroscience and molecular biology labs and UCSC with its genomics institute is an undisputed leader in genomics and bioinformatics. Santa Cruz is a wonderful small progressive town on the central coast of sunny California, nestled into mountains that teem with giant redwood trees, approximately 35 minutes to San Jose/Silicon Valley or 1 hour and half to San Francisco Bay.
The Kim Laboratory aims to investigate connectivity, development, genetic identity, and function of neural circuits using mouse cerebral cortex as a model system. Our ultimate goal is to understand the fundamental principle of neural connectivity and its functions in animal’s perception and behavior. We address our questions using novel neural circuit tracing systems with next-generation trans-synaptic viral tracers, mouse genetics, single-cell genome-wide sequencing, and in vivo imaging. My lab is determined to offer excellent research opportunities to advance your scientific career, strong academic interactions and collaborations across the neuroscience and other biology laboratories at UCSC and more. For additional information, please refer to the following webpage: http://www.ejkimlab.com/
We prefer, but not limited to, candidates with expertise in the following areas: (neuro)developmental biology with genomics experiences, mouse surgery and handling related to neural circuit tracing and manipulations, and molecular and cellular neuroscience. Above all, outstanding applicants with strong quantitative skills are strongly encouraged to apply. Interested individuals should submit an application with a curriculum vitae, a brief cover letter including research interests, and the contact information of three individuals who will provide letters of reference to:
Euiseok Kim, Ph.D.
Assistant Professor
Department of Molecular, Cell, and Developmental Biology University of California, Santa Cruz
ekim62@ucsc.edu
The University of California is an Equal Opportunity/Affirmative Action Employer. All qualified applicants will receive consideration for employment without regard to race, color, religion, sex, sexual orientation, gender identity, national origin, age, disability, protected veteran status, or any other characteristic protected by law.
Join our team! The University of Washington Department of Orthopaedics and Sports Medicine is seeking a Postdoctoral scholar in the Musculoskeletal Systems Biology Lab (MSBL) of Ronald Kwon (https://sites.uw.edu/msblgroup/). The lab is housed in the Institute of Stem Cell and Regenerative Medicine (https://iscrm.uw.edu) in Seattle, Washington. This is an NIH-funded position with potential renewal for up to three years.
The goal of this project is to identify causal genes underlying genetic risk for osteoporosis. Our team has identified genetic variants that protect some individuals from osteoporosis, and make others more susceptible to this disease. To identify the causal genes underlying these risk variants, our lab has developed a rapid zebrafish-based pipeline for the prediction of human skeletal gene function. The candidate will use this pipeline to identify novel genes regulating bone biology and which mediate genetic risk for osteoporosis. Projects feature in vivo modeling in zebrafish, CRISPR-based gene editing, next generation phenotyping, and bioinformatics.
About the MSBL
The MSBL has three major research thrusts: 1) genetic risk for osteoporosis, 2) axial skeletal development, and 3) appendage regeneration. Our focus is on taking bold, innovative approaches.
The MSBL believes that diverse experiences and perspectives are mandatory for scientific excellence (https://sites.uw.edu/msblgroup/equity/). We welcome all willing participants – regardless of gender, race, ethnicity, age, disability, sexual orientation, beliefs, or socioeconomic or cultural background – and work to support each other through trust, encouragement, and honest feedback.
What we are looking for
We are not looking for specific experience or skill but rather candidates with a strong interest in our work. We are also looking for candidates with any combination of the following: curious, creative, passionate learners, willing to take risks, and motivated to take on new challenges. Finally, we are looking for candidates who possess excellent communication skills and a strong commitment to diversity.
Qualifications
Applicants must have a Ph.D. degree (or equivalent) in engineering, life sciences, or a related field at the time of start date.
Application instructions
Applicants should send their CV and cover letter to ronkwon@uw.edu.
Currently, bioinformatics is playing an increasingly important role in life science research. Biologists, clinicians and biomedical researchers have become more dependent on bioinformatics outcomes. Despite the crucial role of bioinformatics in accomplishing multidisciplinary projects, collaborations between biologists and bioinformaticians encounter several difficulties. Here, I outline different types of collaborations and provide an overview of how the relationship between bioinformatics and life science experts can be facilitated.
Potential means of collaboration
Several options for collaboration are available to research groups. Depending on resource availability, they can hire their own bioinformaticians, collaborate with a bioinformatics group within their organization, use the services of an internal bioinformatics core facility, or employ a bioinformatics consulting company outside their organization.
If a bioinformatician is hired by a given research group, all members of the group can have easy access to bioinformatics assistance, communication is easy, and both life scientists and the bioinformatician will feel they have ownership and input into the project, so conflict over assigning credit for output is less likely. However, the work environment might be less competitive for bioinformaticians hired for a specific task because there is insufficient opportunity to expand their expertise, given that they focus on certain types of data related to a specific topic, and there may be minimal constructive criticism from colleagues.
If a research group decides to collaborate with an internal bioinformatics group, creating a good working relationship is more challenging. Bioinformatics groups within academia typically have their own research projects, so their ability to assign time and services to other groups may be limited. Consequently, it is important that resource allocation and expectations should be clearly established and both sides should agree on anticipated credit gains. The risk for bioinformaticians in this scenario is that despite devoting time and expertise to external projects, their contributions might not be valued sufficiently. For example, despite a bioinformatician providing data/results of publishable quality, they may not be assigned a correspondingly prominent position in the authorship list of the resulting scientific papers.
Nowadays, many research institutes have established their own bioinformatics core facilities, with the objective of supporting all research groups in the institute. This could represent an ideal model of collaboration provided that the core facility has a sufficient number of experts, expertise, and resources to tackle the research questions it is presented with. In reality, given the considerable diversity of life science data, newly established core facilities are unlikely to have a sufficient depth of experience to handle all types of data. Accordingly, research groups should be aware of such limitations and must be willing to help core facility bioinformaticians to develop their skills. Where such core facilities are overburdened with requests, there might also be a significant delay in data analysis and revisions.
Finally, outsourcing data analysis by recruiting the services of a professional bioinformatics consultancy is another option, but it appears to be adopted less in academic contexts. Compared to bioinformatics core facilities in academia, professional consultancies tend to be better at project management and generally do not expect authorship rights in publications. However, they are likely to be more expensive, accessibility might be more limited compared to the previous options, and the limitations of core facilities can also be relevant to external consultancies.
Who should sit where?
Bioinformatics is many things. As an interdisciplinary field of science, it has multiple applications including database creation and management, development of software and analytical tools, creation and implementation of computational pipelines to analyze next generation sequencing data, gene expression studies, prediction of macromolecular 3D structures, drug design, precision medicine, phylogenetic studies, amongst many others.
This multitude of applications means that bioinformaticians also tend to have different specialties. It is relatively rare to find a bioinformatician that possesses experience in all or even many of these applications. Life science data is diverse, expansive and complex. Mining such “big data” to extract useful knowledge is complicated and requires careful analysis using appropriate techniques. Mistakenly, bioinformaticians might be seen as “a jack of all trades” by some life scientists, who may think that a bioinformatician should be able to do all types of analysis quickly just by running a few lines of code.
To achieve a successful collaboration, it is crucial that all contributory parties clearly establish the goals, requirements and scope of the project, allowing the right person(s) to be recruited for the right task. For example, if a specific algorithm or computational tool must be developed for a project, it would be more relevant to recruit a bioinformatician with a computer engineering background who can rapidly develop the desired tool. Alternatively, if assistance in data analysis is needed to answer a specific biological question, then it would be better to recruit a bioinformatician with a biological background, who could better comprehend the research context and apply or modify appropriate tools and pipelines to fulfill the needs of the research group. Since biological applied research often involves several rounds of data analyses, data optimization based on feedback, and repetition of pipelines on different datasets, strong lines of communication are essential.
Similar principles should be considered when selecting the leaders/coordinators to manage multidisciplinary projects. A bioinformatics leader should be familiar with the challenges of a broad diversity of bioinformatics applications. He or she should be acutely aware that applied works are as challenging as development tasks and that sufficient time and resources should be allocated to teaching bioinformatics to biologists. It is crucial to understand the needs of life science researchers and to plan resources accordingly so that those needs can be met. The leader of a bioinformatics group should also ensure that the right person(s) is assigned to each project and that whoever requested bioinformatics help is comfortable with the person and process allocated to them.
Assignment of credit
Appropriate assignment of credit is another important factor to maintaining a high level of motivation in collaborations between life science and bioinformatics experts. Credit should be distributed fairly between those who own the scientific idea, those who produce the primary data, and those who add value to it through data analysis or the development of analytical tools. Assigning credit in multidisciplinary projects is a relative concept, and it can be a significant source of conflict, being very much dependent on the characteristics, scope and contributors of a project.
If development of algorithms and computational pipelines is the main focus of the project, most of the credit is attributed to the bioinformaticians whereas, in applied works, partitioning of credit can be more challenging because measuring added value and comparing it among contributors is difficult. Since the life sciences largely remain the domain of biologists, there might be a risk for bioinformaticians to be viewed more of as service providers rather than scientific partners.
Conclusion
Conducting multidisciplinary projects is challenging and success requires a coordinated effort by all contributing disciplines. To facilitate the cooperation necessary between bioinformaticians and life scientists, firstly, it is important to bear in mind that the life sciences and bioinformatics are dependent on each other. Without bioinformatics it would be impossible to manage and analyze the ever-growing amounts of data from life science research and, without that “big data”, bioinformatics could not gain its prestige.
Secondly, human resources have a central role in creating the good working relationships necessary to enable successful collaborations. It is crucial to find a suitable bioinformatician for each role, to be clear about expectations, to provide opportunities for skill development, and to listen to feedback, all of which will help ensure that good bioinformaticians are retained. Managers have a very important role in facilitating collaborations, and it is their responsibility to create an environment that bolsters employee satisfaction because “people leave managers, not companies”.
Thirdly, the needs, interests and benefits for both sides of a collaboration should be well aligned. Only when everything is based on mutual advantage can optimal performance be attained and everyone involved can prosper. To achieve that, it is better if life scientists invite participation from bioinformaticians during the planning phase of their projects.
Finally, measuring the quality of the collaborative relationship is very important. Efforts should be made to find and apply suitable methods to regularly assess such relationships.
Interneurons are crucial to cortical function and their dysregulation has been implicated in various neurological pathologies, yet how they are generated during development is still poorly understood. A new paper in Developmentinvestigates interneuron neurogenesis in the mouse embryo and its control by Magoh, a component of the exon junction complex. We heard more about the work from the paper’s two first authors, Charles Sheehan and John McMahon, and their supervisor Debby Silver, Associate Professor at Duke University Medical Center in Durham, North Carolina.
Debby, Charles and John (L to R)
Debby, can you give us your scientific biography and the questions your lab is trying to answer?
DS I am broadly trained as a developmental neurobiologist. I have had the opportunity to pursue undergraduate research on the biochemistry of the cytoskeleton, PhD studies on cell migration in Drosophila, and postdoctoral research on neural development of the cerebral cortex and neural crest in mice. I started my independent lab at Duke Medical Center in 2010, where I am now an Associate Professor. The overall goal of my lab is to elucidate fundamental principles governing brain development and contributing to neurodevelopmental pathologies. We specifically investigate development of the embryonic cerebral cortex. A main focus is to understand post-transcriptional control, a fascinating but poorly understood regulatory layer of brain development. We have several questions of interest, including how RNA-binding proteins control cortical development at a cellular and molecular level, and the role of subcellular RNA localization and local translation in neural progenitors. We are also fascinated to understand the genetic underpinnings of human cortical evolution. We are guided by the premise that the same mechanisms at play during normal development were co-opted during evolution and, when dysregulated, can cause neurodevelopmental disease.
John and Charles, how did you come to work with Debby and what drives your research today?
JM I have always been driven to apply science to gain a better understanding of disorders, with the goal of ultimately developing novel therapies to treat a clinical need. After finishing my graduate studies, I came across Debby’s lab, which was doing fascinating work and working on very translatable science.
CS I first learned about Dr Silver’s work when I was applying for Research Technician jobs in 2017. When I interviewed with her, there was a genuine shared excitement for science that convinced me to join her lab.
What first got you interested in the role of post-transcriptional regulation in interneuron development?
JM Interneurons have been of interest to me since my graduate work on epilepsy. In my opinion they have been a largely understudied and underappreciated cell type, especially given their prominent role in numerous disorders. Likewise, there are significant gaps in our understanding of post-transcriptional regulation in both ‘normal’ and disease states. Therefore, I found exploring the role of the exon junction complex in interneuron development a particularly exciting project to work on.
DS Development of cortical inhibitory neurons is essential for establishing brain circuitry and is relevant for the etiology of many neurological disorders. Yet in contrast to the field of excitatory neurogenesis, and despite a lot of excellent work being done, I have been struck by the fundamental gaps in our understanding of the genesis of interneurons. For example, we lack a basic understanding of the role of the cell cycle in directing interneuron fates, as well as how RNA regulatory modules shape development. Given our lab’s prior work on Magoh’s role in the cortex, I felt we were in a unique position to make contributions to this field and address these knowledge gaps. I was also motivated to pursue this line of research given the increasing human genetic data linking the exon junction complex to neurodevelopmental pathologies.
Can you give us the key results of the paper in a paragraph?
JM The findings of the paper center around the integral role of Magoh in interneuron progenitors. We find absence of Magoh is detrimental to the balanced production of progeny in neural stem cells. We identify that a contributing factor to the pathogenic brain development is the activation of the p53 pathway. These findings raise important questions about the role of post-transcriptional regulation in neurogenesis.
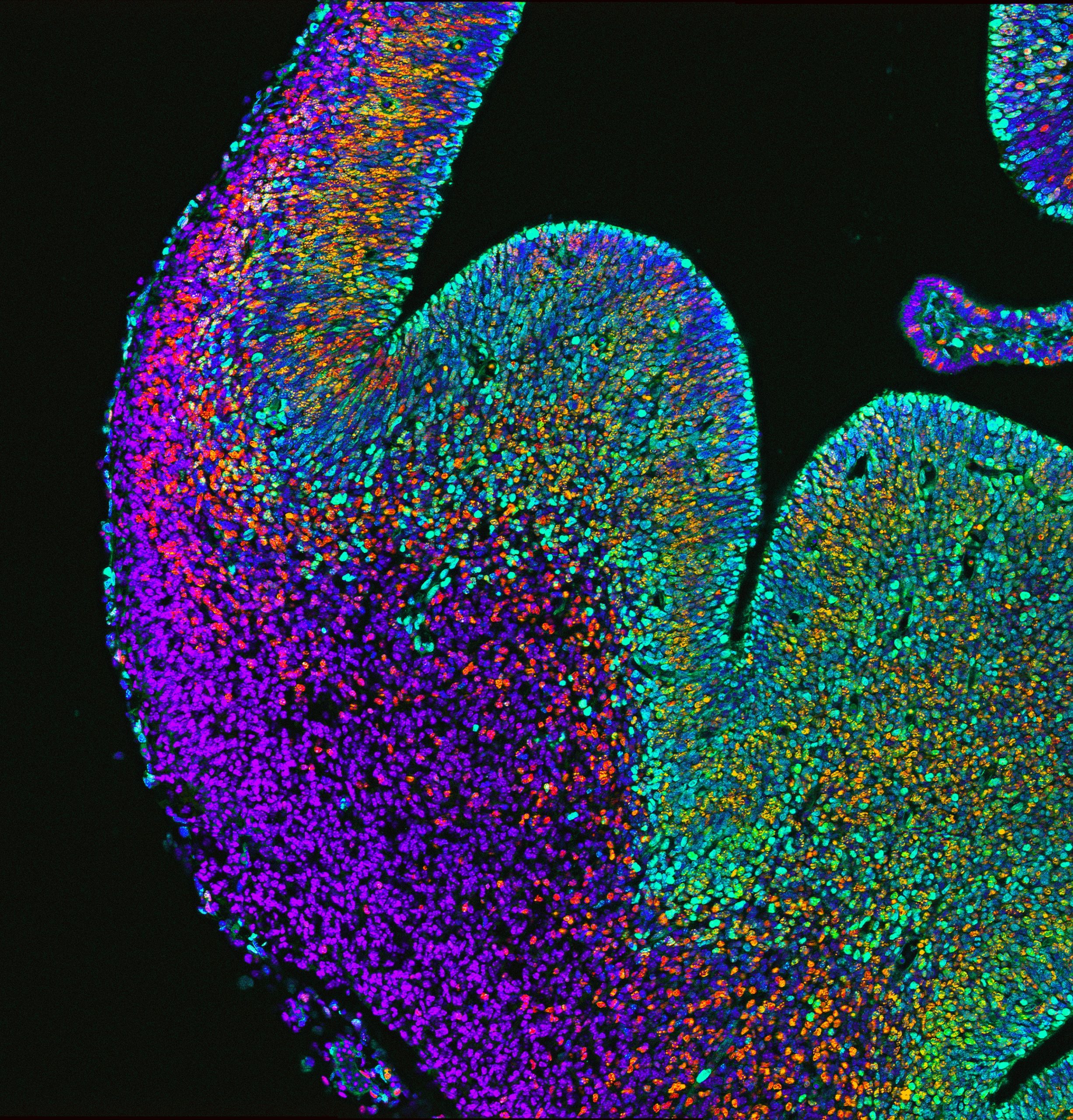
Immunofluorescence depicting the embryonic brain and progenitors (green) of the ganglionic eminence giving rise to newborn interneurons (red) that migrate into the developing cerebral cortex
What do you think explains the difference in the effect of p53 loss in Magoh heterozygotes compared with homozygotes?
JM I think it really comes down to a dose-dependent function of Magoh, and likely the exon junction complex. We have seen dose dependence in a number of our conditional mutants, which points toward a criticality in the balance of EJC components. If a cell loses some Magoh expression, it seems to drive a less severe phenotype. A more extensive depletion results in a striking amount of cell death. My suspicion is that there is a fundamental role of the EJC components in mitosis, which we have yet to fully uncover.
CS We believe that p53 loss differentially affects Magoh heterozygotes and homozygotes because of the severity of their mitotic defects. Simply put, p53 deletion cannot rescue daughter cell death if the progenitors can’t divide. How Magoh dosage causes these differential phenotypes via transcriptome alteration and the effect that p53 loss on has on these changes would be of interest to understand.
DS What we discovered in this study, as well as in prior research, is that cells undergoing mitosis are exquisitely sensitive to Magoh levels. Losing a single copy of this gene causes progenitors to be delayed in mitosis, resulting in inappropriate activation of p53 signaling in newborn progeny, likely due to accumulating DNA damage. We postulate that in Magoh homozygotes, interneuron loss is caused by two defects: p53-dependent apoptosis of newborn cells and p53-independent depletion/death of precursors.
What does your study suggest about the role of EJC components in neurodevelopmental pathologies?
JM This study, along with others from the Silver lab, highlight that EJC components are absolutely necessary for proper function of neural stem cells, likely serving a critical role in mitosis.
CS This work suggests that neurodevelopmental pathologies caused by EJC mutations may not be caused solely by dysfunction of excitatory progenitors, but also interneuron progenitors. This finding gives a more complete understanding on how mutations in Magoh, and other EJC components, alter neural stem cell function and cause neurodevelopmental disorders.
DS Copy number variations and point mutations in EJC components are linked to intellectual disability, autism and microcephaly. Our study, together with prior work from our lab, suggests that mutations in EJC components disrupt proper generation and survival of both excitatory and inhibitory neurons. It may be that in patients carrying EJC mutations, there is imbalanced production and/or survival of these two populations or there may be qualitative aberrant defects in viable interneurons. Historically, microcephaly is linked to abnormal mitosis, with a heavy emphasis on cortical neurogenesis. Our study indicates that mutations affecting mitosis of interneuron progenitors may also be relevant for the etiology of microcephaly.
When doing the research, did you have any particular result or eureka moment that has stuck with you?
JM After a session of live imaging, reviewing the videos always provided me with tremendous insight into the dynamics of neural progenitors and cell divisions. Not necessarily a eureka moment, but very insightful and absolutely beautiful to watch.
CS I think the most memorable eureka moment was when I was imaging the p53 rescue brains. Immediately when looking at the cKO, you could tell that there was still a drastic loss of cortical interneurons, which was unexpected based on what we knew at the time.
Many who have been at the bench for a while will tell you science is not for the weak hearted
And what about the flipside: any moments of frustration or despair?
JM I think many who have been at the bench for a while will tell you science is not for the weak hearted. From what I have seen, even the best scientists suffer more failures than successes. There were plenty of late-night experiments that were revealed to be pure failures once the results were in, and plenty of time spent trouble-shooting technical issues before seeing the light at the end of the tunnel.
CS Similar to the eureka moment, the p53 KO rescue experiments were quite challenging. The mouse genetics required to obtain the samples was a difficult and long process. Even once we had the data, interpreting how p53 was functioning took several discussions and looking back to other data until we reached on our current model.
So what next for you two after this paper?
JM Since my time in Debby’s lab I have been working in regulatory affairs, and furthering my understanding of how a concept grows from an idea, to the bench, to a clinical trial, to a marketed therapy.
CS I have recently started my PhD at Duke in Cell and Molecular Biology and am currently trying to decide on a lab and mentor for this next step.
Where will this work take the Silver lab?
DS We would like to better understand the molecular mechanisms by which Magoh dosage impairs interneuron development, including its role in splicing and translation and its direct targets, as well as discriminating why Magoh loss causes such a striking mitotic defect. It is also exciting to consider whether all EJC components act similarly in development, and how the EJC fits into a larger network as a master regulator of corticogenesis. Beyond the EJC, these studies have sparked our interest in understanding fundamental aspects of interneuron development and how genetic modifications over the course of evolution have shaped this process.
Finally, let’s move outside the lab – what do you like to do in your spare time in Durham?
JM Hiking in Eno park or exploring a new brewery.
CS Durham has a lot of great local breweries, restaurants and concerts, so I spend most of my free time exploring those with friends.
DS I spend my free time mainly with my husband, two kids and our new doggie. We enjoy the outdoors, including mountain biking in our backyard, as well as the great food and music scenes in Durham and Carrboro.
The group of Chen Luxenburg at the Faculty of Medicine, Tel Aviv University, invites applications for a PhD student position.
Our laboratory is looking for excellent and highly motivated PhD students to study the role of the actin cytoskeleton in skin development. Our goal is to understand how cytoskeleton derived signals regulate stem cell ability to create the skin epidermis during development, maintain it in the adult, and repair it upon wounding. On top of molecular biology, tissue culture, advanced microscopy, and mouse work, we utilize state of the art technology that allows us to rapidly manipulate the function of any gene of interest in epidermal stem cells in utero. Several exciting projects are available for successful candidates.
We offer state of the art laboratory with a dynamic and international atmosphere and full financial support (tuition and stipend)
Candidates should hold a Master’s degree in Biology/Life-sciences or related fields.
Interested candidates should email their CV and a brief paragraph describing their research experience and career plans to Chen Luxenburg (lux@tauex.tau.ac.il)
Today, we move to mouse adult haematopoiesis, with an elegant work on inflammation and stem cell biology. As always, feedback is more than welcome!
Chronic inflammation is one of the classic events underlying aging and infection response. When associated with loss of quiescence in haematopoietic stem cells (HSCs), it is commonly the initiating event in leukemia progression. Chavez and colleagues present the impact of chronic interleukin-1 (IL-1) exposure on the most immature HSCs. First of all, they exposed adult mice to IL-1β for 20 days, observing major changes in the HSCs transcriptome. In particular, IL-1β treatment reduced the expression of genetic programs associated with cell cycle progression and protein synthesis. In vitro short-term culture of the most immature HSCs (called LT-HSC), in the presence of IL-1β, confirmed a slower cell cycle entry. Using a genetic reporter for the myeloid master regulator Pu.1, the authors proved that IL-1 is directly linked to the transcription factor increased activity. Specifically, PU.1 is able to bind more than two-thirds of the genes downregulated by IL-1β exposure. Finally, the authors used a genetic mouse model for reduced Pu.1 activity, proving elegantly that an inefficient PU.1 opens the way to the aberrant increase in LT-HSCs cell cycle activity and protein synthesis, leading to an increased cellular pool of stem cells.
Taken altogether, those results describe an IL-1 – PU.1 system that physiologically contributes to stabilizing the number of LT-HSCs. Those results are relevant, because IL-1β prematurely initiate myeloid differentiation, and in general promote the expansion of white blood cells. How to conciliate therefore those apparently contradicting results? Here, the authors focused on a smaller subset of HSCs, and affirm that PU.1 in these cells acts as an activation barrier against unwanted proliferation, while still able to prime HSCs for myeloid differentiation. It will be interesting to assess if other signals could activate similar pool size regulation mechanisms for HSCs.
Chavez J et al. “PU.1 enforces quiescence and limits hematopoietic stem cell expansion during inflammatory stress”
Among many other (many more important) things, pandemic times have meant an end to travel, which for the Development team means no conferences and no lab visits. For me personally, it’s meant cancelled trips to Shanghai, Kyoto, Washington and Chicago, and while I’m delighted to be keeping my carbon footprint down, I’m definitely missing the opportunity to get out and meet the developmental biology community – and hear about your latest work.
I do think, and hope, that this travel hiatus will lead to long-term changes in the way we do conferences – and a more sustainable and eco-friendly conference culture (as discussed in this recent preprint, preLight and Node post). But I also believe that nothing beats face-to-face meetings, and I’m looking forward to getting back on the road once it’s safe and practical to do so. In the meantime, we at Development have been thinking about ways to connect with and support the community while we’re all stuck at home. As James Briscoe wrote in his recent update, we’re always happy to hear from you. So if you’ve got a paper you might be interested in submitting to Development, would like to discuss an idea for a possible review, or just want to talk about the journal or publishing more generally, please get in touch. You can drop us an email and, if it’d be helpful, we can try and set up a phone or video call.
We’ve also been spending some time giving (hopefully!) useful tips to the community here on the Node: if you’ve not read them already, check out Seema’s recent post on writing reviews, and Alex’s piece on how to get involved in peer review. If there’s something you’d like us to write about, do let us know and we’ll see what we can do. But we’d like to be more interactive – we’d normally be out and about giving talks at institutes or panel discussions at conferences, so are thinking about ways of replicating some of this online.
I’m sure many of you already have far too many Zoom calls and webinars to attend, but would you be interested in joining us for webinars on various aspects of publishing? We’ve set up a brief survey for you to indicate your interest and to let us know what you’d be most keen to hear about. Please do fill it in if you’d like to get involved and we’ll try and set something up. We’re also considering the possibility of hosting a series of scientific webinars, perhaps featuring authors of Development papers – again, please let us know in the survey if you would be interested in a devbio webinar series. Finally, we’re beginning to think about whether we could replace in-person lab visits and institute seminars with virtual equivalents – we’ve got limited capacity to do this, but if you’re interested in having one of the Development team ‘visit’ your institute, let me know!
One of the joys of working in this field is the wonderful community of developmental biologists. So while we can’t meet in person, I hope we can continue to connect in other ways – and if you’ve got thoughts about other things we should be doing to help with this, I’m all ears…!
Stay well and stay safe, and see you on the other side…
Anne McLaren – photo courtesy of The Gurdon Institute
In the latest episode of Genetics Unzipped, Kat Arney reflects on the life and work of Dame Anne McLaren, one of the leading embryologists of the 20th century, whose work underpinned the development of the in vitro fertilisation (IVF) techniques responsible for bringing millions of ‘test tube babies’ into the world, and more besides.
Not bad for someone who ended up studying biology at Oxford University almost by accident. Apparently she picked the course because cramming for the zoology entrance exam seemed like an easier option than doing the required reading for English literature.
Anne’s curiosity about early mammalian embryos was the fire that fuelled a lifetime of research. Her groundbreaking work in the 1950s laid the foundations for in-vitro fertilisation, cloning and genetically engineered mice — technologies that have revolutionised human reproduction and biomedical research.
Later in life, she turned her attention to what she used to call “the most fascinating and deeply mysterious cells of all” — the germ cells that will become eggs and sperm in an adult, which are specified just a few days after fertilisation. Anne devoted the rest of her research career to pinning down the characteristics of these unique cells, trying to understand where they came from, where they were going, and what made them so special.
As well as being a highly respected scientist, Anne was also deeply involved in the scientific community. She was the first woman to serve as an officer of the prestigious (not to mention male dominated) Royal Society, and also sat on the Warnock Committee – the fore-runner of the Human Fertilisation and embryology Authority.
She was also a role model and mentor to countless scientists, many of them women working hard to establish a career in a male-dominated world. Anne occasionally complained about the “old boy’s network”, which she felt sometimes led to men only putting forward male friends for jobs. But she did note wryly that there seemed to be an “old women’s network” developing, at least in her field of developmental biology, which was helping to even up the balance.
As she wrote in an eloquent review paper on genetic inheritance, “history may be circular, but the history of science is helical: it repeats itself, but each time at a deeper level”. Rather than the Newtonian idea of standing on the shoulders of giants, Anne saw scientists as forming a twisted helix through time, intertwining as we pass on skills, knowledge and friendship to those who come up behind us.
Anne McLaren’s influence on the world of reproductive science and medicine corkscrews deeply back in time, and legacy will stretch for years to come in the lives of those she knew, those who knew her work, and the many, many more who benefit from it.
If you enjoy the show, please do rate and review on Apple podcasts and help to spread the word on social media. And you can always send feedback and suggestions for future episodes and guests to podcast@geneticsunzipped.com Follow us on Twitter – @geneticsunzip
In pre-COVID19 times, back when conferences happened largely in person, early-career researchers (ECRs) often asked me how they could get more direct invitations to be a reviewer. Peer review is a crucial part of the publishing ecosystem and therefore it’s not uncommon for group leaders to invite members of their lab to review articles with them as part of their academic training. However, direct invitations to review mark a point at which trainees begin feel established in the field (along with moves to start their own group) and can make a useful contribution to a CV. During the current disruption, some reviewers might be feeling overburdened, and editors might be looking to expand their pool of reviewers. Here, I’ve provided a short guide with tips on how ECRs can make themselves known to editors with the hope of receiving direct invitations to be a reviewer. This post does not prescribe what makes a “good” reviewer, nor does it provide training on how to write a peer review report, but you can read advice from Elsevier’s Reviewer Hub or participate in training programmes from Publons Academy, Society for Neuroscience and Genetics Society of America.
So, to stand a chance at being invited to review a manuscript, editors need to know just two things: (1) who you are and (2) your scientific expertise. Editors can’t invite reviewers they don’t know exist or can’t find! Each editor will have a different strategy for finding potential reviewers, and these approaches might vary between professional vs. academic editors. Ideally, an editor aims for three reviewers with complementary expertise to cover the major topics in the manuscript. At Development (and presumably other journals), we also consider aspects of diversity, including career stage, geographical location and gender – see this Editorial by James Briscoe and Katherine Brown. But where do editors look for inspiration?
Author suggested reviewers Most manuscript submission systems allow authors to suggest (or exclude) particular researchers for reviewing their manuscript. Editors will check that these suggestions are sensible, and that the suggested reviewers have the right expertise. It’s also important that there is no conflict of interest between parties, which could be negative (e.g. a direct competitor) or positive (e.g. a previous member of the lab or a recent collaborator). Note that editors are unlikely to only use reviewers suggested by the author, and it is up to the editor who they decide to invite to review a manuscript, although we do respect exclusions.
Reference list Editors might also look through the references cited in the manuscript to see whether it heavily features the work of another group or – if they know the field well – whether certain relevant publications are missing. Authors of these publications might be invited to review the manuscript.
Journal databases Most journals have databases containing researchers, expertise terms and records of previous reviews. These records might detail the number of reviewers a researcher has accepted (or declined), the length of time they took to complete their report, previous reports and, in some cases, a ranking system where the editor can comment on the quality of the report.
Talks, seminars, conferences and meetings For editors, talks and conferences are a great way to meet new researchers. They also provide a broader view of the field as a whole, highlighting areas of debate, uncertainty or controversy, which is useful to know when selecting balanced reviewers.
Publications Editors might be inspired by other articles they’ve handled recently and look to the authors of those articles to review new work. In addition, they might get ideas from publications in other journals within the field or through keyword searches of publication databases such as PubMed.
Google search A Google search using key terms can be a useful way to widen a reviewer pool. Such key terms might be the subject area, a particular technology, specific gene name etc. Editors will explore group and departmental websites to have a clear understanding of the group’s research focus, background and publications.
Social media Social media might be another way for editors to meet reviewers, although this may lead to bias towards particularly vocal scientists. I (and others) have also used the Node Network to find reviewers – read more about the Node Network in this announcement.
Expert advice Usually, if an invited reviewer is unable to review an article, there is the option for them to suggest alternative researchers and it’s really helpful for them to do so! Editors might also seek the advice of their Advisory Editorial Board, either directly or for reviewer suggestions.
Editor expertise Particularly experienced editors will have a thorough understanding of the community in their field and will be able to choose reviewers based on their own knowledge of everything above.
So, with these things in mind, here are three tips for how you can expand your own profile:
1. Get involved: Participate, publish, present and be pro-active
Participate in co-reviewing manuscripts with mentor or group leader and ask them to provide your name to the journal to acknowledge your contribution (see “get credit”’ below). If you’ve left the lab and your former mentor or group leader is unable to review a manuscript, you can ask that they suggest that the editor contact you in their place. Some publishers also have the option to “volunteer” to review, such as Elsevier’s “VolunPeer” initiative and some individual journals – such as eLife, STAR protocols, Stem Cell Research and PeerJ journals – have opportunities to sign up.
Your publication record is the primary way you can demonstrate your research interests and expertise. Although this obviously isn’t the main incentive for publishing, each publication has the added benefit of exposing your name to editors and it might get you inside a journal database.
Try to present your research at conferences, either through a talk or a poster, so that editors become familiar with you and your work. You can also use social media to disseminate your research to a wider audience or talk about subjects that interest you.
You can also be pro-active by approaching editors at meetings and conferences to talk about your work and your interests – when doing so make sure that the editors are left with a clear idea of the topics you cover in your research. Along similar lines, you can email journals and editors to ask to be added to their reviewer database – make sure to also include a list of key terms for your expertise and keep this profile up-to-date (see “get up-to-date” below). The Node Network has been set up specifically for developmental and stem cell biologists with the aim of finding speakers and reviewers that would not normally come to mind – add yourself! Other initiates also exist, such as the GoogleDoc mentioned below.
Postdoctoral Fellows, if you are willing to review manuscripts, please put your information in this form. Please share if you think this is a valuable resource.https://t.co/ljyQm3mXC3
— Dr. Michael D. L. Johnson (@blacksciblog) May 2, 2020
2. Get up-to-date
Ask that your institution or departmental page is kept up-to-date, easy to find and easy to navigate, or include a link to your professional/group website that you can keep on top of it yourself. Make sure that within your page you include a list of your recent publications and keep this updated. Also have a page or paragraph that outlines your research interests as specifically as possible; details of any interdisciplinary research, as well as the model organisms or technologies that your group utilises, are also useful for editors to know.
Keep your profiles on various databases and directories (such as the Node Network, Google Scholar, ORCID and ResearchGate) up-to-date with your current institution, email address and expertise. If old information is in a database, then you might miss out on an invitation; although an editor should check for that your most recent contact information is correct before they get in touch! Again, an accurate departmental website can be crucial for editors to know old email addresses from new ones.
3. Get credit
As mentioned above, if you have participated in peer review with your mentor or group leader, consider asking them to provide you with some credit. At Development, we encourage lab members to contribute as co-reviewers and have a specific part of the report where these lab members are named. The names provided here are still withheld from the authors, but the journal will contact to the co-reviewer and invite them to join the journal’s database.
You can also receive credit through initiatives that track peer review activity, such as Publons and ORCID. Some publishers, such as Elsevier, also have reviewer recognition platforms, which allow editors to acknowledge reviewers that provide particularly useful reports with a “certificate of excellence”.
Finally – and perhaps more controversially – you could consider signing the report if the journal allows you to do so. There are various arguments for and against waiving anonymity, but if the authors felt that your report was particularly constructive they might suggest you to review future manuscripts. In journals where peer review reports are published alongside the article, the recognition might go even further. Ultimately, you should do what make you comfortable and therefore allows you to produce the most constructive report possible.
I’d like to finish by saying thank you to all the researchers that participate in peer review, especially those who have taken the time to review manuscripts for Development. Although the publishing landscape is changing, and there are important discussions to be had around this, peer reviewers continue to be an essential part of moving science forward.
 (No Ratings Yet)
(No Ratings Yet)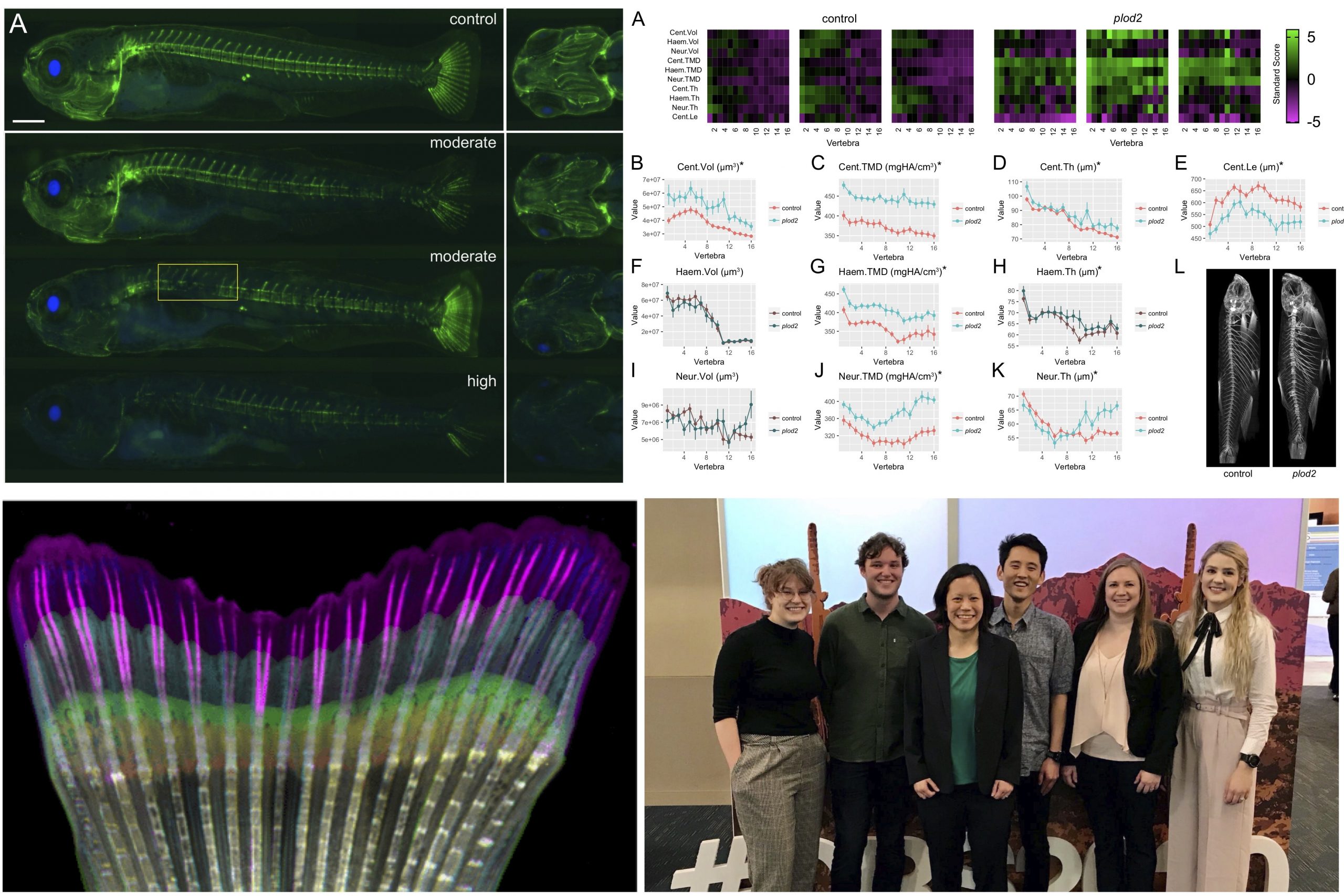
 (1 votes)
(1 votes)

 (11 votes)
(11 votes)