The Developmental Biology Group (Prof. I. Lohmann) at the Centre for Organismal Studies (COS) in Heidelberg, Germany is looking for a
POST-DOC
to study the role of HOX transcription factors in controlling cellular plasticity and cell fate maintenance.
You will join the Lohmann lab (http://ilohmann-lab.org), which is located at the Centre for Organismal Studies (COS) at the University of Heidelberg in Germany, and studies the role of HOX transcription factors in defining cell type identities using Drosophilaas a model.
Cell fates are controlled by networks of transcription factors (TFs) that activate transcriptional programs realizing the distinct properties of cells of a given cell types.However, how TFs control different cell fates is still un unsolved question. HOX TFs represent an excellent model to address this fundamental problem, since they are broadly expressed yet perform highly specific functions within different cell types. We have previously shown that Hox TFs stabilize cell fate choices by suppressing the multipotency encoded in the genome via the interaction with the Polycomb complex. We now seek for enthusiastic new colleagues to analyse our hypothesis that elimination of the Hox code, which is maintained throughout the lifetime of an organism, results in “memory-less” naïve cells that are easy to reprogram.
The Heidelberg Molecular Life Science Community offers a vibrant molecular research community, as well as state-of-the-art core facilities.
Successful candidates should have experience in genomic approaches like RNA-seq, ChIP-seq, ATAC-seq, analysis of genomic data, confocal microscopy, advanced immunohistochemistry and possibly in Drosophila genetics and molecular biology. The ability to quickly integrate into an interdisciplinary team and work independently within an academic research environment is essential. The position is immediately available, the salary is according to TV-L regulations. Disabled persons with comparable skills will be preferentially considered.
The University of Oklahoma College of Arts and Sciences is excited to announce three open faculty positions at any rank in the Department of Biology. As part of our Biology of Behavior strategic initiative, the department invites individuals with creative, innovative, and dynamic research programs who are interested in joining a strong group of researchers to apply for these faculty positions:
A Geneticist who uses integrative molecular approaches to understand the evolution, specification, and/or regulation of how genes affect organismal behavior.
A Physiologist who studies the endocrine regulation and modulation of behavior.
An Evolutionary Developmental Biologist who studies how developmental processes give rise to organismal morphology, nervous system structures, and/or physiology that lay the foundation for the generation of behavior.
The anticipated start date is August 2020. For additional details on these positions, applicant qualifications, and how to apply, please visit http://ou.edu/bb and http://www.ou.edu/cas/biology.
Screening of candidates will begin October 15, 2019 and will continue until the positions are filled.
The University of Oklahoma is an EO/Affirmative Action institution http://www.ou.edu/eoo/. Individuals with disabilities and protected veterans are encouraged to apply.
How would you create a hole between two sticky surfaces? Simply crack it!
At a first glance, trying to pull apart the two surfaces seems to be a good idea, but in practice, you might need a lot of energy. However, it seems that the mouse embryo has found a smart and efficient way to do so during its pre-implantation development. After three rounds of cellular divisions, the 8-cell stage embryo starts to compact: cell-cell contacts are expanding, making the embryo more spherical instead of a collection of bubble-like cells [1]. After another round of cleavage, it also internalizes the cells that are more contractile [2]. They will become the Inner Mass Cell (ICM), the future fetus proper, while less contractile cells, the Trophectoderm cells (TE) form a squamous epithelium, that surrounds the ICM and will become part of the placenta. From this step, the embryo is almost spherical, with two layers of cells.
Then, at the 32-cell stage, the embryo shows a new feature: a lumen, a fluid-filled cavity, that breaks the previous radial symmetry by forming at the interface of TE and ICM cells. To grow a lumen, three conditions are needed: 1- to have a sealed compartment, here ensured by the tight junctions between TE cells at the embryo surface; 2- to draw water towards the sealed compartment: in our case, the mouse embryo builds an osmotic gradient by pumping ions in the intercellular medium and lets the water flow through pores; 3- and finally, you have to make room for the accumulated fluid. But here is the problem: the blastocoel forms systematically on the basolateral side of the TE cells, where the cells strongly adhere together! In most other examples of lumen formation, the opening happens at the apical side of the epithelium, where adhesion is repressed! Thus, arises the question: how can you create a lumen at the adhesive side of cells?
In our research [4], we combine developmental biology and physics to decipher the mechanisms of the embryogenesis. We have found that the apparition and the positioning of the lumen, the so-called blastocoel, can be explained using simple physical and biological concepts.
The formation of the blastocoel was a long-time debated topic. Studies have mainly focused on the expansion phase, when the blastocoel is already positioned, while its initiation and positioning are still poorly understood.
What Julien did first, with the help of Francesca and Ludmilla, was to look at the steps preceding the apparition of the blastocoel. In the last decades, efforts have been done on culture conditions and advances in microscopy have permitted to reduce light exposition while improving spatiotemporal resolution. Using resolutive imaging in space and time, that involved the use of both transgenics and microscopy techniques, heobserved the embryo literally boiling!Hundreds of bubbles appeared at the intercellular contacts before the final lumen, forming a network of small microlumens throughout the embryo. Some of those microlumens grew in size, while others disappeared (Fig. 1). As biologist, this observation might seem not significant, but for physicists, this coarsening process immediately rang a bell: looking at the movies, it was really analogous to a well-known process in soft-matter physics: Ostwald ripening. Basically, it describes how in a vinaigrette, the droplets of vinegar will coarsen into fewer drops, the bigger droplets growing to the detriment of the smaller.
From this observation, the collaboration between the two teams emerged, with one team of biologists (Julien, Francesca, Ludmilla and Jean-Léon), the other of physicists (Mathieu, Annette and Hervé), with two questions: i- how these water pockets form in spite of cell adhesion? ii- as they form ubiquitously through the embryo, what mechanisms ensure the formation of a single blastocoel and its final positioning?
Figure 1: Microlumens appear at the onset of cavitation and coarsen to form a final fluid-filled lumen, the blastocoel, breaking the symmetry of the embryo.
The microlumens form and expand in the extracellular space, at the interface between cells. Julien looked at those cellular contacts, showing that during the formation of microlumens, the spatial distribution of adhesion molecule (E-cadherins) evolves from a homogeneous to a localized heterogeneous distribution. From this observation came the idea of hydraulic fracturing, where water pressure cracks cell-cell contacts exactly like it would crack the rock in oil fracking [3].
After discussions, we came with two main scenarii. a- as cells are active material, they could autonomously regulate their adhesion and create weak points where the fluid could accumulate; b- adhesion is a force that opposes to fluid accumulation, and the expansion of microlumens is capable of pushing adhesion molecules away. To answer this, we had no direct way to measure inside the embryo how cells react against an increase of pressure in the intercellular space. So instead, we chose to inhibit (in three different ways) the formation of the microlumens. in the absence of microlumen, we couldn’t see any reorganization of the E-Cadherin. Our favorite interpretation from this result: this is the hydraulic pressure that breaks locally the adhesion between the cells, and from these breaking points, microlumen can expand. In a nutshell, the embryo seems to generate an increase of hydraulic pressure to break apart all cells contacts instead of specifically regulating its adhesive properties.
A coarsening process is generally made possible by the exchange of matter between different compartments. In the mouse embryo, the microlumens can exchange fluid via the intercellular contacts, which connect them throughout the embryo. A coarsening process akin to Ostwald ripening furthermore involves two other key features: the only stable state is a single droplet, and it requires a surface tension at droplet interfaces, which generate the pressure driving fluid exchange. In the embryo, we invariably observe the formation of a single lumen, and it is furthermore always located at the interface in between the TE and ICM. Thus, we quickly came to the idea that playing on the cell “surface tension” would give us great insights into the mechanical aspects of the blastocoel formation. Indeed, according to previous studies [1], we knew that ICM and TE cells have different levels of contractility, that can physically be translated into surface tensions.
We therefore built a theoretical model of the network of microlumens as a two-dimensional graph of connected hemispherical drops, to test in silico the physical predictions with an algorithm developed by Annette and Mathieu, and we designed experiments to test in situ the biological predictions. Our combined results suggest that, due to osmotic gradient and active pumping, the cells inject fluid that pressurizes the intercellular space, hence creating the hundreds of microlumens by disrupting the cell-cell adhesive contacts. The newly formed microlumens then coarsen into a single final lumen, with a characteristic biphasic dynamic of collective growth then shrinkage, observed both for the model and for the myriad microlumens measured by Julien.
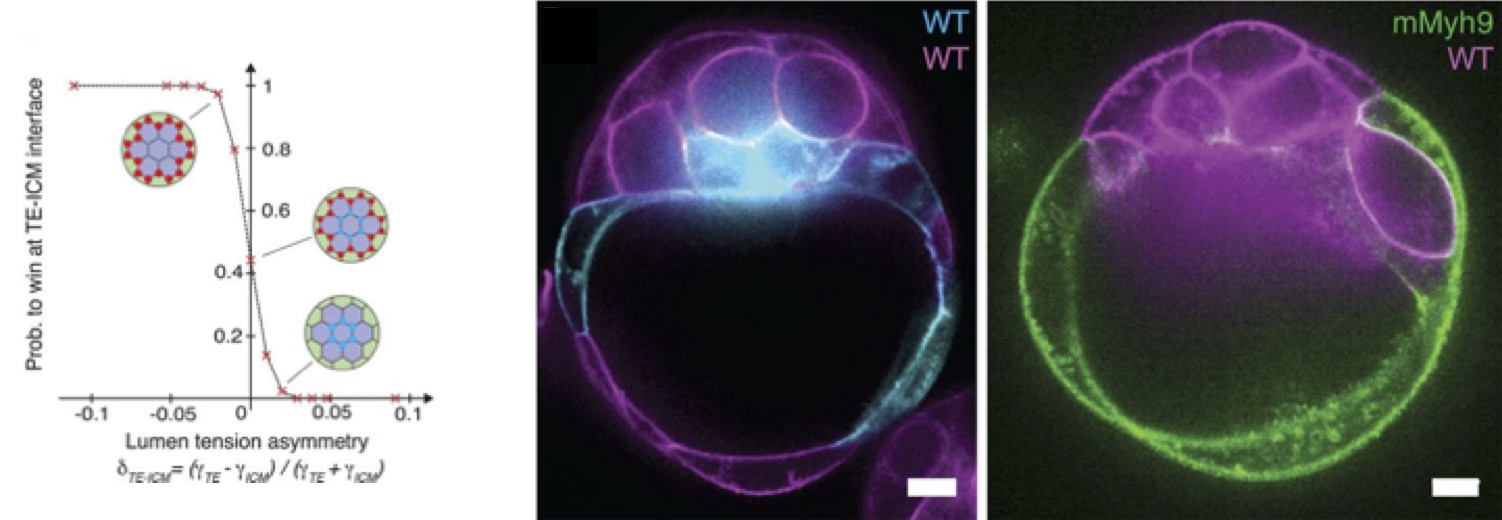
From there, Mathieu predicted with the model the formation of the blastocoel on the side of the embryo, between TE and ICM cells, hence breaking the symmetry of the embryo (Fig. 2., left panel). This prediction was tested using chimeric embryos that Julien made (Fig. 2, center and right panels): an equal mixture of wild-type cells and low-contractility cells, deficient in myosin activity, shows a clear bias for the final position of the lumen toward the low-contractility domain of the chimera, as the theory predicted. The experiments of Julien on low-adhesion mutants, lacking half of E-cadherin activity, also predicted effects of the partial loss of adhesion, that were then tested by Mathieu on the theoretical model, confirming the whole process as being a trade-off between adhesion and contractility.
Figure 2: (Left) Probability of blastocoel formation for ICM-ICM (blue) or TE-ICM (red) multi-cellular lumens vs the tension asymmetry ratio from simulations. (Right) In chimeric embryos, the final position of the blastocoel is biased towards less contractile part of the embryo (Myh9 deficient cells).
One fascinating aspect of the process is how much it is robust: even though breaking adhesive cell-cell contacts may not be thought to be the best process to form a lumen, the mouse embryo succeeds to form an internal cavity. Moreover, despite the immense molecular complexity of embryo development, the formation of the blastocoel follows rather simple physical mechanisms.
The project was going back and forth between theory and experiment all along. The geographic proximity and the excellent relationship between the two teams were key factors, speeding up the process and the constant exchanges, helping us to remove the barrier between theoretical biophysics and developmental biology.
It was a quick and extremely stimulating project for both our young teams. Part of the pleasure of the project was to gather many people with various expertise, and to see all the pieces matching together to give a comprehensive model at many levels, and of course to work with such enjoyable people.
I, Julien, come from the zebrafish community, and I was used to image embryos that develop fast, as the embryo looks like a fish in 24 hours after fertilization. Then I started to work in Jean-Léon’s team and image relatively slow embryonic development (the mouse embryo takes 3 days to build the blastocyst). Nonetheless, I was convinced that we were missing (and most probably are still missing) key steps of mammalian development and decided to push the system further. Having time resolution of minutes or seconds led to these incredible observations and were key is the direction in which we pushed our research. What I will retain from this work is the exciting collaboration with Annette, Mathieu and Hervé, that opened a new field for me: I must confess, I never heard of coarsening before! I am really happy to see that physicists can be as amazed as developmental biologists by embryogenesis and that these enthusiastic interactions can lead to exciting discoveries.
As far as I (Mathieu) am concerned, this was my first real scientific contribution, ending with a beautiful paper. The fact that such a key step in the mouse embryo development can be simply seen as a fracking and coarsening process still amazes me. Starting the study of the morphogenesis of the mouse embryo was a real challenge with my background of theoretical physicist. Hopefully, Julien and others from his team were always more than happy to speak, and to show me what they were doing, which was an invaluable help. I could not think of better conditions as a start for my PhD, and I am really thrilled to see where it will go.
Figure 3 : Representative set of collaborators (n = 6). From left to right: Ludmilla, Julien, Francesca, Jean-Léon, Hervé and Mathieu.
It opened so many foods for thoughts, promising new and exciting results about the development of the mouse embryo, that we are now trying to push forward. Are there factors that favor the final position within the embryo or is it a stochastic phenomenon? What are the consequences of this increase of pressure on cells at molecular and genetic levels? What triggers the initiation and nucleation of the microlumens?
The Hamdoun and Lyons Laboratories at U.C. San Diego’s Scripps Institution of Oceanography (https://scripps.ucsd.edu) are seeking a highly motivated postdoc candidate for a joint fellowship at the intersection of evolutionary and ecological developmental biology. The successful candidate will be nominated for the competitive Scripps Postdoctoral Scholar (SPS) Award (https://apol-recruit.ucsd.edu/JPF02248). The Hamdoun Lab (www.hamdounlab.org) focuses on the function of small molecule transporter systems in early embryogenesis. The Lyons Lab (www.lyonslab.org) focuses on the evolution of gene regulatory networks and morphogenesis. The postdoc will be part of a motivated group of students, staff, postdocs and visiting scholars within the two labs, who are broadly interested in cell and developmental biology of marine organisms.
A potential focal point of this postdoc position is the opportunity to take advantage of Lytechinus pictus, an emerging genetically-enabled echinoderm species being developed in our labs, which is useful for live imaging, cell biology, developmental biology, genomics, and toxicology. Resources available to the candidate include larval culturing facilities, a fully sequenced genome, and a new marine transgenics facility for maintenance of stable lines.
Additionally, the Lyons and Hamdoun Labs offer a broad range of other systems for comparative developmental studies among echinoderms and molluscs, and provide a highly interdisciplinary and collaborative environment between our groups and among other labs at the Scripps Institution of Oceanography, with UC San Diego’s main campus, and with other academic and industrial partners in the greater San Diego Area.
Eligibility:
To qualify, applicants must have a Ph.D. (or be close to earning one), have experience in molecular biology and/or genetics, and be eligible for nomination for the Scripps Postdoctoral Scholar (SPS) Award. Strong training in transgenesis, genome engineering technology (including CRISPR/Cas9 editing), gene delivery techniques (e.g. microinjection, electroporation), and cis-regulatory element analysis are highly desirable.
To Apply:
Interested candidates should submit the following items, as a single PDF, to Drs. Hamdoun (hamdoun@ucsd.edu) and Lyons (d1lyons@ucsd.edu):
1) Cover letter explaining your interest in the position and qualifications
2) CV
3) Statement of research/career goals
4) Names and contact information for at least three references
Review of applications will begin immediately and Drs. Hamdoun and Lyons will work directly with the successful candidate to develop their SPS Award application.
The Department of Cell, Developmental and Integrative Biology (CDIB) https://www.uab.edu/medicine/cdib/ at the University of Alabama at Birmingham (UAB) seeks highly-qualified applicants for a position of tenure-earning Assistant Professor with a research focus on mechanisms regulating development and organogenesis, developmental origins of birth defects, or developmental programs involved in tissue growth and regeneration. Applicants with a background in using animal models are strongly encouraged to apply.
Candidates should be highly motivated with excellent communication, teaching, and writing skills, and to have demonstrated ability to conduct advanced research. Candidates are expected to establish a robust and externally funded research program and contribute to teaching and mentorship activities. Applicants who can bring innovative approaches that complement existing expertise within the department will be given preference.
The successful applicant will benefit from newly renovated research facilities, strong institutional and departmental commitments to career development, and a vibrant, collaborative research environment. A competitive compensation and startup package is being offered.
Interested applicants should submit a cover letter, short essays describing past research activities and future research goals, curriculum vitae, and contact information of three references to: https://uab.peopleadmin.com/postings/5364.
UAB is an Equal Opportunity/Affirmative Action Employer.
The Center of Regenerative Medicine (CRM), together with the Departments of Cell Biology & Physiology, Developmental Biology, Genetics, Neurosurgery, Pathology and Immunology, and the Divisions of Nephrology and Gastroenterology within the Department of Medicine at Washington University in St. Louis invite applications at the level of assistant professor on the tenure track. Faculty in the CRM and these departments and divisions employ a broad range of cellular systems and model organisms to explore fundamental and translational questions in regenerative medicine.
We are seeking outstanding colleagues with an interest in any area of regenerative medicine, including the genetic and epigenetic control of pluripotent, adult, or cancer stem cells; cell fate specification and reprogramming; repair and replacement of lost or damaged tissues; gene therapy; and tissue engineering.
The CRM is a trans-institutional, interdisciplinary center with more than 80 faculty members representing diverse aspects of regenerative medicine. To serve the needs of its members, the CRM has research cores, as well as discussion groups and trainee fellowships. More information about the CRM can be found at: https://regenerativemedicine.wustl.edu/
Review of applications will begin October 15, 2019.
Interested applicants are required to submit a cover letter, curriculum vitae, and summary of their research accomplishments and plans through our online system: https://facultyopportunities.wustl.edu/apply/Posting/Detail/1010435. Please indicate in your cover letter which department(s)/division(s) you work mostly closely aligns with. We expect that selected candidates will join one or more of the participating departments and the CRM. Applicants should also be prepared to provide the names and email contact information for three referees to provide letters of recommendation during the application process.
Washington University seeks an exceptionally qualified and diverse faculty; women, minorities, protected veterans and candidates with disabilities are strongly encouraged to apply.
Written by: Aida Rodrigo Albors, Laura Pellegrini and Neil Dani.
This July, The Company of Biologists Workshops together with Maria K. Lehtinen and Fiona Doetsch organized a meeting titled “New Frontiers in the Brain: Unexpected Roles of the Choroid Plexus-Cerebrospinal Fluid System in Health and Disease”. Set in the gorgeous countryside of Steyning in the United Kingdom, Wiston House (circa. 1576), with its historic and charming past, served as a fitting venue for a workshop on a research topic that has captured scientific minds for millennia. Indeed, Hippocrates (460–370 B.C.) and Claudius Galen from Pergamon (130–200 A.D.) believed that the “pneuma psychikon” or “spiritus animalis,” which roughly translates to “the life force that powers the mind,” was located in the brain’s cerebral ventricles that contain the choroid plexus-cerebrospinal fluid (CSF) system. Over time, more fantastic but ultimately incorrect conclusions arose from the inherent challenges of studying the CSF system that lies deep within somewhat inaccessible parts of the brain. For example, it was once thought that the brain’s ventricles were not filled with CSF but instead with a vapor that ultimately condensed into a fluid upon death due to the drop in body temperature. Correcting such theories took another millennium and the work of several preeminent thinkers including Leonardo da Vinci and Andreas Vesalius (among many others), whose work helped map the ventricular system and confirm CSF flow. Undoubtedly, rewriting theories greatly depended on the continual development of new tools to better access and study the choroid plexus-CSF system.
Now, let us fast forward back to Wiston House in July 2019. Here, a small group of 30 people: 20 senior and 10 early-career researchers, continued the tradition of leveraging new technology to better understand the workings of the choroid plexus-CSF system and their roles in disease. We, the authors of this post, were lucky to count ourselves among the early-career researchers and would like to take the opportunity to share our experience with you. Laura Pellegrini is a postdoctoral researcher working at the MRC Laboratory of Molecular Biology (Cambridge, UK), in Madeline Lancaster’s group and is developing cutting-edge organoid models to study the secretion and barrier functions of the choroid plexus. Neil Dani, a postdoctoral fellow in the laboratory of Maria Lehtinen (Boston Children’s Hospital, USA) is applying single-cell transcriptomics to characterize the cellular composition of the choroid plexus and is developing tools to study tissue function in vivo using mouse models. Aida Rodrigo Albors, a Marie Skłodowska-Curie fellow in the lab of Kate Storey (Dundee, UK), after working with axolotls to study ependymal cell recruitment in spinal injury models, now uses single-cell technologies to disentangle the heterogeneity of spinal cord ependymal cells in mice. Since our field is relatively small with almost no specialized meetings, we and the other researchers were keen on attending the workshop to find peers who were equally dedicated to a research topic that is heating up and attracting researchers with diverse training. It turned out that our expertise complemented each other in just the right ways, which aroused exciting conversations and ideation throughout the workshop.
Group picture on the lawns of Wiston House.
The wide range of topics covered included the role of the choroid plexus-CSF system during embryonic development and adult life, the liver-like functions of the choroid plexus, and the emerging contributions to human disorders such as autism and childhood carcinomas. Next, we saw cutting-edge single-cell RNA-sequencing technologies applied to comprehensively characterize the cell populations that form the choroid plexus and ependymal cells that line the central canal of the spinal cord. Some investigators also shared their extraordinary live imaging techniques to observe choroid plexus function and CSF flow across the ventricles in mice and zebrafish, as well as in vitro-derived choroid plexus cells and choroid plexus organoid models that make this difficult-to-reach tissue more accessible for manipulation and functional studies. Leveraging these new technologies will surely give us a better understanding of the mechanisms of the choroid plexus-CSF system to uncover its role in health and disease.
The Company of Biologists Workshops has a winning formula when it comes to hosting scientists to debate an exciting and upcoming field. For example, junior and senior investigators were given the same amount of time for their talks, thus making everyone feel like an equal contributor. This approach helped create a comfortable environment from the start and helped spur animated discussions well beyond the scheduled question time, into the tea breaks, during meals, and even during the hike across the South Downs National Park that borders Wiston House. Each speaker was also given the freedom to present a topic of their choice, which allowed postdocs to share their most exciting and unpublished data, junior investigators to share their research vision, and senior investigators to flesh out historic landmark findings – altogether giving us a glimpse into the past, present, and future of the field.
Celebrating a successful workshop. Cheers!
Another highlight of the workshop was the amazing dining spread for each of the meals, which definitely helped fuel the discussions and excitement, for which we extend our heartfelt thanks to the amazing staff at Wiston House. Finally, the team behind The Company of Biologists Workshops deserves high praise for organizing an incredible event and ensuring that everything went off without a hitch – not a trivial task for an international conference that represented researchers from 11 countries! The relationships forged during the workshop will surely continue beyond Wiston House – already embodied by our team-writing this piece for the Node (which took place across three countries!). So, we hope that you, the reader, will feed off our enthusiasm and explore the exceptional Biologists Workshop series to find ways to help shape an exciting research area and cherish the experience as we will surely continue to do.
Stowers Research Conferences are biennial meetings that bring together researchers, faculty, and graduate students to inspire creative thought and collaboration in the basic sciences. The Developmental Cell Biology meeting will include 11 speakers with 13 talks chosen from abstracts, 2 poster sessions and a conference party to close the meeting.
Retinal ganglion cells connect the retina to the brain, and their degeneration underlies glaucoma, which is the leading cause of irreversible blindness in humans and currently untreatable. Replacement of lost cells could be achieved by in vivo reprogramming of endogenous cells in the retina, a strategy explored by authors of a new paper in Development who focus on a transcription factor renowned for its reprogramming ability in other contexts. We caught up with first author Maurício Rocha-Martins and his former supervisor Mariana Silveira, Professor at the Federal University of Rio de Janeiro in Brazil, to find out more about the story.
Maurício (L) and Mariana (R)
Mariana, can you give us your scientific biography and the questions your lab is trying to answer?
MS My research started off in microbiology, first as a technician in biotechnology and then as an undergraduate at the Federal University of Rio de Janeiro (UFRJ). Although my degree was on Microbiology and Immunology and I carried out research in drug resistance in bacteria and in Plasmodium falciparum, this shifted to neuroscience when I joined the lab of Rafael Linden for my doctoral studies at the Institute of Biophysics Carlos Chagas Filho, a renowned biomedical institute in Brazil. There, co-mentored by Marcelo T. Bozza and Fernando G. Mello, my longstanding interest in retinal neurochemistry and molecular neurobiology started with studies on neuropeptide regulation of cell death and differentiation in the developing and mature retina. As a postdoc visiting Victor May at the University of Vermont, I was drawn to the Klf family of transcription factors even before Klf4 became famous as one of the Yamanaka factors. In my own research group, my interest in developmental biology led me to start working on retinal cell reprogramming with talented students. We are interested in understanding the molecular mechanisms of cell fate acquisition as building blocks for the design of new regenerative strategies. Particularly, in this first study we wanted to elucidate Klf4-dependent mechanisms of retinal cell fate acquisition. The findings suggest that these mechanisms could be applied to design novel regenerative strategies to replace and reconnect retinal ganglion cells (RGCs) lost in the adult retina as a result of neurodegeneration in glaucoma.
And Maurício, how did you come to work in the Silveira lab?
MR-M My interest in science began at a young age. I used to do experiments around the house at the age of ten, and I even kept a little science notebook. My family was always supportive, even when, for example, I needed to use the freezer to test how long insects can sustain cold temperatures. It was no surprise then that I started a bachelor’s degree in biology at the UFRJ, and soon enough I got a real internship in a real lab.
I was sure I wanted to be an entomologist, but slowly I became more and more interested in developmental biology, in particular how the different steps of embryo and organ formation are coordinated in time. The idea of an internal clock that determines when different types of cells are generated really fascinated me. Towards the end of my graduation I was fortunate enough to meet Mariana, who introduced me to a powerful system to study temporal control of development: the vertebrate retina. The neurons of the retina come in six ‘flavours’ and are generated in a stereotypic birth order. The predefined order and windows of cell type specification make the retina an excellent system for studying the mechanisms underlying temporal progression. I then dedicated my PhD in the Silveira lab to studying how neurogenesis is controlled in time and how to manipulate the birth order of RGCs, a disease-relevant cell type.
Before your work, what was known about the role of Klf4 in RGC development and regeneration?
MS Other research groups and our own had shown that RGCs express Klf4, and the influence of Klf4 and other members of the family in axon growth have been studied in depth. Moore et al. (2009) showed that Klf4 inhibits axonal growth, as its specific deletion in RGCs increased axon growth and regeneration, and the same group recently suggested that Klf4 regulates axon density during retinal development, but not RGC specification and survival. However, a direct role of Klf4 in RGC generation remained unexplored, which we addressed by combining a conditional knockout mouse in which the Klf4 gene is deleted very early in retinal development and CRISPR/Cas9-based approaches in zebrafish. Interestingly, Todd and Fischer (2015) had also showed that Klf4 was upregulated during endogenous reprogramming of Müller glial cells in chickens. Müller glia in fish are an endogenous source of regeneration, and upon damage can re-enter the cell cycle and generate all retinal neurons. However, this capacity is reduced in chicken and almost lost in mammals. These lines of evidence encouraged us to pursue the study of Klf4 in retinal development and cell reprogramming.
Induced ganglion cells (iRGC) generated in vivo upon Klf4 overexpression for 30 days. iRGC (GFP-positive, depth colour code) project TUBB3-positive axons which are aligned with endogenous fascicles (green).
Can you give us the key results of the paper in a paragraph?
MS We wanted to know whether Klf4 played a role in the development of specific cell types in the retina, in particular RGCs. To answer this, we used loss-of-function approaches in both mouse and zebrafish. The data showed that Klf4 might not be essential for RGC generation, although other family members could compensate for its absence, as was shown in embryonic stem cells (Jiang et al., 2008). What was really striking was that when we overexpressed Klf4 in late retinal progenitors, which have a restricted neurogenic potential, these progenitors re-acquired the potency to generate RGCs. Next, we asked what changed in the expression pattern of these late progenitors shortly after Klf4 overexpression. We detected the upregulation of some characteristic elements of the molecular programme of early retinal development, such as Atoh7, which is essential for RGC generation. Although these induced RGCs (iRGCs) lacked some markers of mature RGCs (such as Brn transcription factors), they projected axons toward the head of the optic nerve and survived up to 30 days after the beginning of Klf4 overexpression, the longest time analysed. In conclusion, this single transcription factor can reprogramme the fate of late retinal progenitors that normally generate other types of retinal neurons. This opens the door to investigating Klf4 as a promising candidate for new strategies to regenerate RGCs at late disease stages of glaucoma.
Even though your iRGCs matured and survived for a long time, they didn’t express known regulators of RGC maturation and survival: why do you think that is?
MS Our working hypothesis is that the role of Klf4 as a pioneer factor might affect epigenetic modifications crucial to the progression of retinogenesis, which normally prevents the neurogenic programming of RGCs at late stages of retinal development. Then, to reprogramme late retinal progenitors, Klf4 might interfere in mechanisms that, in combination with Atoh7, succeed in the generation of RGCs. However, although iRGCs do survive for at least 30 days, the programme activated by Klf4 may not follow the exact time course of gene expression of the key transcription factors responsible for terminal differentiation and survival, such as Brn3b. Alternatively, the sustained expression of Klf4 could interfere with later steps in iRGC differentiation. In fact, Klf4 has been described as an inhibitor of axonal outgrowth. So, we believe that although it activates a programme that is efficient in giving rise to RGCs, in the long run Klf4 may impair the final stages of the process. Either there is a missing step and/or Klf4 may actively inhibit maturation.
More longer term, what do you think are the advantages (and disadvantages) of cell reprogramming compared with cell therapy for the treatment of glaucoma?
MS There are different strategies involving cell therapy and cell reprogramming. One which has shown promising results uses stem cells as a source for the secretion of neurotrophic factors. This and other neuroprotective approaches, including gene therapy, are efficient to prevent the progression of RGC degeneration. However, in late stages of disease progression, RGC regeneration would be essential. The generation of RGCs in vitro either from embryonic stem cells or induced pluripotent stem cells to transplant into affected eyes is an alternative, but cost, safety and low efficiency are the major limitations of this approach. Finally, an appealing option is to coax endogenous cell sources to replace the lost RGCs. In this context, re-activating the intrinsic regenerative potential of Müller glial cells is an interesting option, and has potential advantages related to cell integration; however, it is very early to state that it may lead to a treatment for glaucoma. The first step would be to demonstrate that functional RGCs can be generated in vivo from Müller glia, and to attain visual function recovery in preclinical models of glaucoma.
When doing the research, did you have any particular result or eureka moment that has stuck with you?
MR-M As a junior group leader, Mariana was very hands-on, helping with bench work whenever possible, so it was no surprise that we shared the eureka moment of this project. We went together to the microscope to check what long-term overexpression of Klf4 does to progenitor cells: could Klf4’s reprogramming abilities could be used to promote de novo genesis of RGCs? I had electroporated the retina of rat pups in vivo, a method that consists of delicate surgery and injection of DNA into the subretinal space. To be honest, because this injection is tricky, we would have been happy to see any sign of electroporation at this point. But, to our surprise, the retinas were nicely electroporated – ‘beginner’s luck’ – and we observed a striking change in the position of the neurons within the retina. In the control, most neurons were located at the apical side, whereas in the Klf4 group they were positioned at the basal side. This change in position indicated a possible change in fate. Looking through more sections we saw beautiful bundles of axons coming from the basal neurons all the way to the optic nerve, a unique feature of RGCs. We were literally jumping with excitement in the microscope room. This finding motivated us to characterize in detail the fate of these basal neurons, and we confirmed that Klf4 overexpression is sufficient to induce de novo genesis of RGCs.
We were literally jumping with excitement in the microscope room
And what about the flipside: any moments of frustration or despair?
MR-M The moments of frustration were usually associated with the lack of infrastructure that is common even in the best universities and institutes in Brazil. Although I was lucky to work in a well-equipped lab, many times I lost experiments due to power blackouts or could not start because there was no water, so no ice. Another recurrent issue is the time it takes to import lab reagents. It is not uncommon to wait 3 months or more to receive an antibody or a restriction enzyme. For these reasons, simple experiments that should take weeks were stretched for months. So, I really would like to thank the labs at the Institute of Biophysics (UFRJ) and neighbouring institutes for sharing reagents and equipment. This work would not have been possible without this collaborative spirit.
So what next for you after this paper – I understand you’re now in Germany?
MR-M Part of my PhD work was carried out in Caren Norden’s lab at the Max Planck Institute of Molecular Cell Biology and Genetics. The goal of this collaboration was to develop zebrafish mutants to complement our mouse data on Klf4 function. But during this time I took on a side project investigating neuronal migration in the retina. This was when I did my first live-imaging experiments of zebrafish embryos and realized how dynamic retinal morphogenesis is. I saw that the different cell types initially share the same space and later sort themselves into layers. It became clear to me that the formation of a functional retina crucially requires that each cell finds its way through the crowded tissue to its correct destination. But, although this neuronal layering is clearly essential for proper circuit assembly and vision, we are just beginning to understand how retinal neurons migrate and de-mix to form layers. So, fascinated by this question, I returned to the Norden lab as a postdoc. I am currently investigating the formation of the layer responsible for collecting light from the environment: the photoreceptor layer.
What is the situation like for developmental biology – and research in general – in Brazil at the moment?
MS These are hard times for research and education in Brazil. We are facing a drastic reduction in research investment from the federal government and the budget now is the lowest in the last 14 years. This affects the number of fellowships awarded to PhD students and postdocs and the budget for research projects. The impact is evident in the reduction of students seeking PhD programmes and in the increase of dropout rates. For this reason, many talented students go to other countries and are not attracted to return to Brazil. This state of affairs, which has been the subject of many articles in scientific journals and reports from scientific societies (see for example https://www.nature.com/articles/d41586-019-01079-9 and https://www.sciencemag.org/news/2019/05/brazil-useful-idiots-protest-cuts-research-and-education), is something that will threaten the future of Brazilian research. I believe one way to overcome difficulties and to survive through this hard period is to intensify international and local research collaborations. But the government must soon realize and embrace the slogan from the Brazilian Academy of Sciences: ‘Science is not an expense. It’s an investment’.
Where will this work take the Silveira lab?
MS Understanding the basic mechanisms associated with the acquisition of specific cell identities in neural development underpins my interests. This work opens crucial questions in the fields of neuronal development and in vivo reprogramming. For example, we are interested in understanding the mechanisms behind the Klf4-induced change in cell fate; in the design of complementary strategies to guarantee the generation of mature RGCs; and in the development of reprogramming strategies directed to Müller glia in disease models of glaucoma. Müller glial cells are good targets if we think about the replacement of RGCs from an endogenous source. To re-activate the regenerative potential that has been lost over evolutionary time is a promising alternative.
I hope that the visibility of this study helps us to build international collaborations to pursue these lines of investigation. Collaborative work with other colleagues and a group of very motivated and talented students were essential to achieve the results we have obtained up to now. However, the situation in Brazil is now very difficult for those who want to pursue a scientific career and it is hard to keep the students in the lab motivated in such scenario. This was also one of the reasons that made me decide to spend a sabbatical period at João Relvas’ lab at the Institute for Investigation and Innovation in Health (i3S, University of Porto, Portugal) to diversify our research lines and build new collaborations. It is a survival strategy.
Finally, let’s move outside the lab – what do you like to do in your spare time?
MR-M: Coming from Rio de Janeiro, I had low expectations about life in a ‘small’ city in Germany. But soon I was surprised by the cultural diversity the city of Dresden has to offer. Now, I occasionally enjoy jazz concerts and even jam sessions of Brazilian chorinho. Dresden also taught me to appreciate nature in a different way. I have replaced the beaches of Rio with hikes through the beautiful Sächsische Schweiz national park. These hikes are usually slow because the kid in me still wants to turn over every rock. Also, my love for insects stays strong. I currently keep a family of stick insects at home … but I don’t do experiments on them anymore.
MS Besides spending time playing with my 4-year-old son, I really love the diversity of Brazilian music, the rhythm and beat of samba, maracatu, coco, congo, ciranda. I try to enjoy it by participating in groups that use the instruments from samba schools to play not only samba but this rhythmic diversity. I learned how to play with musicians in groups like Monobloco and Bangalafumenga, which in the early 2000s contributed to the rebirth of street carnival in Rio de Janeiro. It is really a fantastic experience. I do recommend it.
Image: Skin cancer cells – Squamous cell carcinoma, NIH Image Gallery via Flickr Attribution-NonCommercial 2.0 Generic (CC BY-NC 2.0)
In this episode we’re digging into some of the mysteries around what’s often seen as the ultimate genetic disease: cancer.
We talk to Phil Jones at the Sanger Institute in Cambridge to unpack his latest research showing that low doses of X-rays – similar to what someone might receive from a few CT scans – can trigger the growth of potentially cancerous cells. And we sit down with Steve Elledge from Harvard Medical School to hear how he and his team are using their Cancer Research UK Grand Challenge Award to understand why tumours tend to arise in some tissues and not others. Plus, the evolutionary genetics of rats leaving a sinking ship.
If you enjoy the show, please do rate and review and spread the word. And you can always send feedback and suggestions for future episodes and guests to podcast@geneticsunzipped.com
 (No Ratings Yet)
(No Ratings Yet)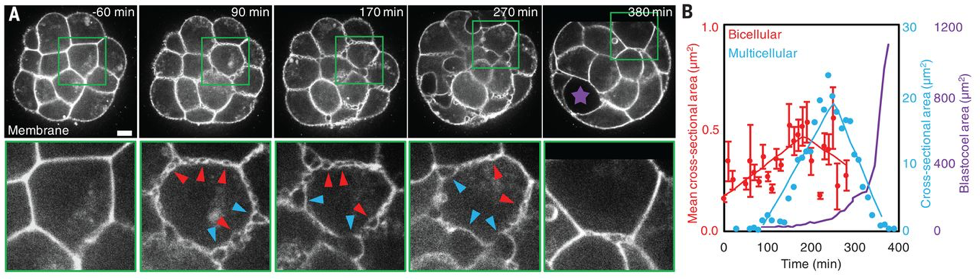

 (3 votes)
(3 votes)